
Abstract
Ligand recognition by cell-surface receptors underlies development and immunity in both animals and plants. Modulating receptor signalling is critical for appropriate cellular responses but the mechanisms ensuring this are poorly understood. Here, we show that signalling by plant receptors for pathogen-associated molecular patterns (PAMPs) in immunity and CLAVATA3/EMBRYO SURROUNDING REGION-RELATED peptides (CLEp) in development uses a similar regulatory module. In the absence of ligand, signalling is dampened through association with specific type-2C protein phosphatases. Upon activation, PAMP and CLEp receptors phosphorylate divergent cytosolic kinases, which, in turn, phosphorylate the phosphatases, thereby promoting receptor signalling. Our work reveals a regulatory circuit shared between immune and developmental receptor signalling, which may have broader important implications for plant receptor kinase-mediated signalling in general.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
Identifiers for published or publicly available lines are provided in the Methods. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE51 partner repository with the dataset identifier PXD031441 and https://doi.org/10.6019/PXD031441. Source data are provided with this paper. All other relevant data are available from the corresponding authors upon request.
References
Hohmann, U., Lau, K. & Hothorn, M. The structural basis of ligand perception and signal activation by receptor kinases. Annu. Rev. Plant Biol. 68, 109–137 (2017).
Dievart, A., Gottin, C., Périn, C., Ranwez, V. & Chantret, N. Origin and diversity of plant receptor-like kinases. Annu. Rev. Plant Biol. 71, 131–156 (2020).
Shiu, S. & Bleecker, A. B. Expansion of the receptor-like kinase/pelle gene family and receptor-like proteins in Arabidopsis. Plant Physiol. 132, 530–543 (2003).
Gómez-Gómez, L. & Boller, T. FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 5, 1003–1011 (2000).
Zipfel, C. et al. Bacterial disease resistance in Arabidopsis through flagellin perception. Nature 428, 764–767 (2004).
Zipfel, C. et al. Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell 125, 749–760 (2006).
Liang, X. & Zhou, J. Receptor-like cytoplasmic kinases: central players in plant receptor kinase-mediated signaling. Annu. Rev. Plant Biol. 69, 267–299 (2018).
Veronese, P. et al. The membrane-anchored BOTRYTIS-INDUCED KINASE1 plays distinct roles in Arabidopsis resistance to necrotrophic and biotrophic pathogens. Plant Cell 18, 257–273 (2006).
Lu, D. et al. A receptor-like cytoplasmic kinase, BIK1, associates with a flagellin receptor complex to initiate plant innate immunity. Proc. Natl Acad. Sci. USA 107, 496–501 (2010).
Kadota, Y. et al. Direct regulation of the NADPH oxidase RBOHD by the PRR-associated kinase BIK1 during plant immunity. Mol. Cell 54, 43–55 (2014).
Li, L. et al. The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity. Cell Host Microbe 15, 329–338 (2014).
DeFalco, T. A. & Zipfel, C. Molecular mechanisms of early plant pattern-triggered immune signaling. Mol. Cell 81, 3449–3467 (2021).
Couto, D. et al. The Arabidopsis protein phosphatase PP2C38 negatively regulates the central immune kinase BIK1. PLoS Pathog. 12, e1005811 (2016).
Holton, N., Nekrasov, V., Ronald, P. C. & Zipfel, C. The phylogenetically-related pattern recognition receptors EFR and XA21 recruit similar immune signaling components in monocots and dicots. PLoS Pathog. 11, e1004602 (2015).
Park, C.-J. et al. Rice XB15, a protein phosphatase 2C, negatively regulates cell death and XA21-mediated innate immunity. PLoS Biol. 6, e231 (2008).
Segonzac, C. et al. Negative control of BAK1 by protein phosphatase 2A during plant innate immunity. EMBO J. 33, 2069–2079 (2014).
Couto, D. & Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 16, 537–552 (2016).
Yu, L. P., Miller, A. K. & Clark, S. E. POLTERGEIST encodes a protein phosphatase 2C that regulates CLAVATA pathways controlling stem cell identity at Arabidopsis shoot and flower meristems. Curr. Biol. 13, 179–188 (2003).
Heazlewood, J. I. et al. PhosPhAt: a database of phosphorylation sites in Arabidopsis thaliana and a plant-specific phosphorylation site predictor. Nucleic Acids Res. 36, 1015–1021 (2008).
Mergner, J. et al. Mass-spectrometry-based draft of the Arabidopsis proteome. Nature 579, 409–414 (2020).
Thor, K. et al. The calcium-permeable channel OSCA1.3 regulates plant stomatal immunity. Nature 585, 569–573 (2020).
Hazak, O. & Hardtke, C. S. CLAVATA 1-type receptors in plant development. J. Exp. Bot. 67, 4827–4833 (2016).
Nimchuk, Z. L. CLAVATA1 controls distinct signaling outputs that buffer shoot stem cell proliferation through a two-step transcriptional compensation loop. PLoS Genet. 13, e1006681 (2017).
Goad, D. M., Zhu, C. & Kellogg, E. A. Comprehensive identification and clustering of CLV3/ESR-related (CLE) genes in plants finds groups with potentially shared function. New Phytol. 216, 605–616 (2017).
Fletcher, J. C. Recent advances in Arabidopsis CLE peptide signaling. Trends Plant Sci. 25, 1005–1016 (2020).
Yamaguchi, Y. L., Ishida, T. & Sawa, S. CLE peptides and their signaling pathways in plant development. J. Exp. Bot. 67, 4813–4826 (2016).
Anne, P. et al. CLERK is a novel receptor kinase required for sensing of root-active CLE peptides in Arabidopsis. Development 145, dev162354 (2018).
Hazak, O. et al. Perception of root‐active CLE peptides requires CORYNE function in the phloem vasculature. EMBO Rep. 18, 1367–1381 (2017).
Rao, S. et al. Roles of receptor-like cytoplasmic kinase VII members in pattern-triggered immune signaling. Plant Physiol. 177, 1679–1690 (2018).
Wang, W. et al. Receptor-like cytoplasmic kinases PBL34/35/36 are required for CLE peptide-mediated signaling to maintain shoot apical meristem and root apical meristem homeostasis in Arabidopsis. Plant Cell https://doi.org/10.1093/plcell/koab315 (2021).
Depuydt, S. et al. Suppression of Arabidopsis protophloem differentiation and root meristem growth by CLE45 requires the receptor-like kinase BAM3. Proc. Natl Acad. Sci. USA 110, 7074–7079 (2013).
Stahl, Y., Wink, R. H., Ingram, G. C. & Simon, R. A signaling module controlling the stem cell niche in Arabidopsis root meristems. Curr. Biol. 19, 909–914 (2009).
Crook, A. D. et al. BAM1/2 receptor kinase signaling drives CLE peptide-mediated formative cell divisions in Arabidopsis roots. Proc. Natl Acad. Sci. USA 117, 32750–32756 (2020).
Rodriguez-Leal, D. et al. Evolution of buffering in a genetic circuit controlling plant stem cell proliferation. Nat. Genet. 51, 786–792 (2019).
Yu, L. P., Simon, E. J., Trotochaud, A. E. & Clark, S. E. POLTERGEIST functions to regulate meristem development downstream of the CLAVATA loci. Development 127, 1661–1670 (2000).
Song, S. K. & Clark, S. E. POL and related phosphatases are dosage-sensitive regulators of meristem and organ development in Arabidopsis. Dev. Biol. 285, 272–284 (2005).
Gagne, J. M. & Clark, S. E. The Arabidopsis stem cell factor POLTERGEIST Is membrane localized and phospholipid stimulated. Plant Cell 22, 729–743 (2010).
Song, S. K., Lee, M. M. & Clark, S. E. POL and PLL1 phosphatases are CLAVATA1 signaling intermediates required for Arabidopsis shoot and floral stem cells. Development 133, 4691–4698 (2006).
Blümke, P. et al. Receptor-like cytoplasmic kinase MAZZA mediates developmental processes with CLAVATA1 family receptors in Arabidopsis. J. Exp. Bot. 72, 4853–4870 (2021).
Fujita, S. et al. SCHENGEN receptor module drives localized ROS production and lignification in plant roots. EMBO J. 39, e103894 (2020).
Luo, X. et al. Tyrosine phosphorylation of the lectin receptor‐like kinase LORE regulates plant immunity. EMBO J. 39, e102856 (2020).
Kutschera, A. et al. Bacterial medium-chain 3-hydroxy fatty acid metabolites trigger immunity in Arabidopsis plants. Science 364, 178–181 (2019).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Longair, M. H., Baker, D. A. & Armstrong, J. D. Simple neurite tracer: open source software for reconstruction, visualization and analysis of neuronal processes. Bioinformatics 27, 2453–2454 (2011).
Kurihara, D., Mizuta, Y., Sato, Y. & Higashiyama, T. ClearSee: a rapid optical clearing reagent for whole-plant fluorescence imaging. Development 142, 4168–4179 (2015).
Kang, Y. H. & Hardtke, C. S. Arabidopsis MAKR 5 is a positive effector of BAM 3‐dependent CLE 45 signaling. EMBO Rep. 17, 1145–1154 (2016).
Monaghan, J., Matschi, S., Romeis, T. & Zipfel, C. The calcium-dependent protein kinase CPK28 negatively regulates the BIK1-mediated PAMP-induced calcium burst. Plant Signal. Behav. 10, e1018497 (2015).
Hansen, S. et al. Design and applications of a clamp for Green Fluorescent Protein with picomolar affinity. Sci. Rep. 7, 6292 (2017).
Nimchuk, Z. L. & Perdue, T. D. Live imaging of shoot meristems on an inverted confocal microscope using an objective lens inverter attachment. Front. Plant Sci. https://doi.org/10.17615/c3nc-5w03 (2017).
Marhava, P. et al. A molecular rheostat adjusts auxin flux to promote root protophloem differentiation. Nature 558, 297–300 (2018).
Perez-Riverol, Y. et al. The PRIDE database and related tools and resources in 2019: improving support for quantification data. Nucleic Acids Res. 47, D442–D450 (2019).
Acknowledgements
We thank J.-M. Zhou (CAS, Beijing) for kindly providing published rlck-vii mutants and P. Tarr (Caltech, USA) for the Myr-mTurquoise2 plasmid. The Nimchuk laboratory thanks T. D. Perdue, director of the University of North Carolina—Chapel Hill Genome Sciences Microscopy Core, for assistance with confocal imaging. The Zipfel group thanks all members for discussions and critical reading of the manuscript. This research was supported by the Gatsby Charitable Foundation (C.Z.), the University of Zürich (C.Z.), the European Research Council under the grant agreement nos. 309858 and 773153 (grants ‘PHOSPHOinnATE’ and ‘IMMUNO-PEPTALK’ to C.Z.), the Swiss National Science Foundation (grant agreement nos. 31003A_182625 to C.Z. and 310030B_185379 to C.S.H.), a National Institute of General Medical Sciences—Maximizing Investigators’ Research Award from the NIH (R35GM119614 to Z.L.N.), the National Science Foundation (IOS-1455607 to Z.L.N.), startup funds from Virginia Tech to Z.L.N. and a joint European Research Area Network for Coordinating Action in Plant Sciences (ERA-CAPS) grant (‘SICOPID’) from UK Research and Innovation (BB/S004734/1 to C.Z.) and National Science Foundation (IOS-1841917 to Z.L.N.), respectively. T.A.D. and P.A. were supported by the European Molecular Biology Organization (fellowships EMBO-LTF-100-2017 to T.A.D. and EMBO-LTF-480-2016 to P.A.). T.A.D. was further supported by the Natural Sciences and Engineering Council of Canada (fellowship PDF-532561-2019). P.A. and Y.G. were also supported by a Tremplin grant from the University of Lausanne.
Author information
Authors and Affiliations
Contributions
T.A.D., P.A., S.R.J., C.Z., C.S.H. and Z.L.N. were responsible for conceptualization and methodology. T.A.D., P.A., S.R.J., A.W. and F.S. undertook validation. T.A.D., P.A., S.R.J., A.W., F.S., O.J., P.D., Y.G., Q.W. and S.R. were involved in formal analysis. T.A.D., P.A., S.R.J., A.W., F.S., O.J., P.D., Y.G., Q.W., S.R., A-M.P. and Z.L.N. undertook investigation. T.A.D., P.A., S.R.J., A.W., F.S. and P.D. were involved in data curation. T.A.D., P.A. and S.R.J. wrote the original draft. T.A.D., P.A., S.R.J., C.Z., C.S.H. and Z.L.N. undertook reviewing and editing of the manuscript. T.A.D., P.A. and S.R.J. were involved in visualization. F.L.H.M., C.Z., C.S.H. and Z.L.N. undertook supervision. C.Z., C.S.H. and Z.L.N. were responsible for project administration. T.A.D., P.A., C.Z., C.S.H. and Z.L.N. acquired funding.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 PLL4 and PLL5 dynamically associate with EFR and FLS2 in ligand-dependent manner.
(a) elf18 triggers EFR/PLL4,5 dissociation in planta. CoIP assay of transiently expressed EFR-GFP and HA-tagged PLL4 or PLL5 in N. benthamiana leaves with or without treatment with 1 μM of elf18 for 10 minutes. (b) flg22 triggers FLS2/PLL4,5 dissociation in planta. CoIP assay of transiently expressed FLS2-GFP and HA-tagged PLL4 or PLL5 in N. benthamiana leaves with or without treatment with 1 μM of flg22 for 10 minutes. CBB: Coomassie brilliant blue. Experiments were performed twice with similar results.
Extended Data Fig. 2 The S-X-X-L domain is conserved among the PLL family.
(a) Schematic representation of PLLs protein domains. light blue: S-X-X-L domain, green: catalytic domains. Numbers indicates amino acid residues. (b) Protein alignment of the S-X-X-L domain of POL family members. Pink square: S-X-X-L residues; bold: residues targeted for mutagenesis in current study in POL (black arrow head) and PLL4 (white arrow head). (c) Sequence logo of the S-X-X-L domain of the POL/PLL family created from the alignment in panel B using the WebLogo3 online application (http://weblogo.threeplusone.com). (d) Table resuming the in silico predicted phosphorylation in the S-X-X-L domain identified in PhosPhAt419 and/or Athena databases20. (e) Phylogenetic tree of the POL family based on protein alignment. PP2C38 is used as an outgroup.
Extended Data Fig. 3 PLL4 regulates PTI in a phosphorylation-dependent manner.
(a) BIK1-dependent phosphosites identified on MBP-PLL4* by LC–MS/MS following in vitro kinase assays. Numbers indicate sum of total spectra identified across 4 replicates corresponding to phosphorylation at the specified residue(s) in PLL4. Some spectra were ambiguous for two phosphorylation positions. Sites within the S-X-X-L domain of PLL4 are outlined and spectra corresponding to sites targeted for mutagenesis are highlighted. (b) BIK1 phosphorylates S-X-X-L motif sites within the PLL4 N terminus. Autoradiogram of in vitro kinase assay PLL4-N (WT) or PLL46A-N. Kinase assays were performed three times with similar results. (c) Treatment with 100 nM flg22 induces a phosphosite-dependent mobility shift in PLL4-HA. N. benthamiana leaves transiently expressing FLS2-GFP and PLL4-HA or PLL46A-HA were treated with or without 100 nM flg22 for 10 min prior to protein extraction, λPPase treatment, and blotting. Assays were performed three times with similar results. (d) PLL4 phosphomimetic (PLL46D) mutation disrupts direct interaction between PLL4 and EFR-CD in vitro. Amylose pulldown assay of 6xHis-tagged cytosolic domain (CD) of EFR with MBP-tagged WT version (PLL4) or phosphovariant (PLL6D) of PLL4. Assays were performed three times with similar results. (e) Expression of PLL46A dampens PTI responses in N. benthamiana. ROS burst induction by elf18 (100 nM) on leaf discs of N. benthamiana transiently expressing FLS2-GFP and PLL4-HA variants (curves show mean values ±s.e.). Bottom panel displays the time to maximum response; letters indicate statistically different values (one-way ANOVA with Dunnett’s test); n = 24 leaf discs from 3 biologically-independent leaves. Box plots show 25th to 75th percentile range with a line at the median and whiskers from minimum to maximum values. Assays were performed three times with similar results. (f) S-X-X-L phosphorylation does not alter accumulation of PLL4-YFP at the plasma membrane. Confocal images showing PLL4-YFP (WT or mutant variants; yellow channel) and Myr-mTQ2 (myristoylatable-mTurquoise2, PM marker; blue channel) following co-expression in N. benthamiana leaves.
Extended Data Fig. 4 L135F is conserved among the PBL clade.
Protein alignment of the PBL family. Magenta square: RLCK-VII-5 clade; cyan square: RLCK-VII-8 clade; yellow square: the highly conserved L residue.
Extended Data Fig. 5 RLCK-VII-5 clade kinases are specifically required for CLEp perception.
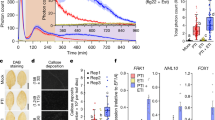
(a-c) RLCK-VII-5 members are semi-redundant in the CLEp signalling. (a) Root length of 8-day-old seedlings grown on media with or without 100 nM CLV3p. NT: not treated. Kruskal Wallis non-parametric ANOVA test, **** indicates p value <0.0001, *** indicates p value <0.001, * indicates p value <0.001, ns: not significant, n = 30 independent biological replicates. (b) Corresponding growth ratio inhibition observed in (a) calculated as following: \(\left( {\frac{{root\,length_{NT} - root\,length_{CLV3}}}{{root\,length_{NT}}}} \right)x100\). All of the rlck-vii-5 mutant combinations were less sensitive to CLV3p than the Col-0 background. (c) 7-day-old seedlings grown on media with 100 nM of indicated CLE peptides. NT: not treated. Letters indicate significant differences within the treatments (ANOVA followed by Tukey test, two-sided). n = 26–46 independent biological replicates. (d) rlck-vii-5 is less sensitive to CLV3p and CLE45p treatments than the dominant negative pbl34-2 mutant. 7-day-old seedlings grown on media complemented with 100 nM of indicated peptides. NT: not treated. n = 19–45 independent biological replicates. (e,f) Complementation of pbl34-3 mutants expressing PBL34::gPBL34-CIT construct. 7-day-old seedlings grown on media complemented with 50 nM CLEp. NT: not treated. Letters indicate significant differences within the treatments (ANOVA followed by Tukey test, two-sided). (e) Complementation assay on CLE45p media, n = 37-58 independent biological replicates. (f) Complementation assay on CLE26p media. n = 17–28 independent biological replicates. (g,h) Complementation of rlck-vii-5 mutants expressing PBL34::gPBL34-CIT construct. (g) Eight-day-old seedlings grown on media supplemented with 100 nM CLV3p (g; n = 25-31 independent biological replicates) or (h) seven-day-old seedlings grown on media supplemented with 50 nM CLE45p (h; n = 22-47 independent biological replicates). NT: not treated. Letters indicate significant differences within the treatments (ANOVA followed by Tukey test, two-sided). (i) Expression of PBL34::gPBL34-CIT in the shoot apical meristem. Yellow, PBL34-CIT; blue, chlorophyll. Image represents cross section through L4 of SAM capturing the organizing centre. Low levels of expression were detected in all cell layers and zones of the SAM and developing floral primordia and meristems. (k) ROS production in response to 100 nM flg22 or elf18. Curves show mean values ±s.e., n = 12 independent leaf discs. All box plots show 25th to 75th percentile range with a line at the median and whiskers from minimum to maximum values. ROS assays were performed three times with similar results.
Extended Data Fig. 6 Pathway-specific roles of PLL family members.
(a) Root length of 7-day-old seedlings in response to 15 nM CLE45p. NT = not treated (n = 15–32 independent biological replicates). Different letters indicate statistically significant differences (p < 0.01, ANOVA followed by Tukey test, two-sided). Box plots show 25th to 75th percentile range with a line at the median and whiskers from minimum to maximum values. (b) QC divisions in response to 100 nM CLE40p. (c) Quantification of response shown in (b); n = 10-18 independent biological replicates. (d) PAMP-induced ROS in pll mutants. Curves show mean values ±s.e., n = 12 independent leaf discs. ROS assays were performed three times with similar results.
Extended Data Fig. 7 Conservation of RK-PBL-PLL circuitry in CLEp signalling.
(a) POL is a substrate of active PBL34. Autoradiogram of in vitro kinase assay incubating equal amounts of MBP-tagged POL with MBP-tagged WT PBL34 or mutant forms of PBL34 (PBL34D275A or PBL34L135F). (b) PBL34 phosphorylates PLL1 in vitro. In vitro kinase assay incubating equal amounts of MBP-tagged WT version (PBL34) or inactive (PBL34*) of PBL34 recombinant protein with MBP-tagged N terminus (PLL1-N), catalytically dead full-length (PLL1*-FL), or catalytically dead C-terminus (PLL1*-C). CBB: Coomassie brilliant blue. Kinase assays in (a) and (b) were performed twice with similar results. (c) PBL34-dependent MBP-POL phosphosites identified by LC–MS/MS following in vitro kinase assays. Numbers indicate sum of total spectra identified across 3 replicates corresponding to phosphorylation at the specified residue(s) in POL. Some spectra were ambiguous for two phosphorylation positions. Sites within the S-X-X-L domain of POL are outlined and spectra corresponding to sites targeted for mutagenesis are highlighted. (d) POL phosphorylation status determines its interaction with CLV1 in planta. CoIP assay of GFP-tagged CLV1 with HA-tagged WT POL or phosphovariants (POL7A or POL7D). (e) POL phosphosites control direct interaction with BAM3 in vitro. Amylose pulldown assay using equal amounts of GST-tagged cytosolic domain (CD) of BAM3 with MBP-tagged WT (POL) or phosphomimetic (POL7D) variants of POL. Experiments in (d) and (e) were performed three times with similar results. (f) S-X-X-L phosphorylation does not regulate POL-YFP accumulation at the plasma membrane. Confocal images showing POL-YFP (WT or mutant variants; yellow channel) and Myr-mTQ2 (myristoylatable-mTurquoise2, PM marker; blue channel) following co-expression in N. benthamiana leaves.
Extended Data Fig. 8 PBL-phosphorylation sites negatively regulate POL function.
(a-c) POL phosphovariants complement pol pll1 to varying degrees. (a) Representative pictures of 4-week-old plants expressing different phosphovariants of POL-HA protein fusion (WT, POL7D or POL7A) under control of native POL promoter. (b) Representative pictures of 6-week-old stems displaying post-seedling stem cell defect-terminated silique phenotype. white asterisk: terminated silique; cyan arrow head: one successful silique formation for entire line of POL7D-HA #1. (c) Corresponding quantification of the shoot complementation based on terminated silique phenotype. n ≥ 942 siliques from ≥30 independent biological replicates (plants) per genotype. (d) Detailed pictures of terminated flower compared to WT non-terminated one. cyan arrow head: presence of the pistil; white arrow head: absence of pistil. Scale bar: 3 mm. (e) Western blot showing expression of POL-HA in POL::POL-HA complementation lines. (f) Relative fold change (∆∆CT) expression analysis of POL in complementation lines. n = 3 independent biological replicates. Error bars indicate ±SD.
Supplementary information
Supplementary Information
Supplementary Tables 1 and 2.
Source data
Source Data Fig. 1
Unprocessed western blots and gels.
Source Data Fig. 2
Unprocessed western blots and gels.
Source Data Fig. 4
Unprocessed western blots and gels.
Source Data Extended Data Fig. 1
Unprocessed western blots and gels.
Source Data Extended Data Fig. 3
Unprocessed western blots and gels.
Source Data Extended Data Fig. 7
Unprocessed western blots and gels.
Source Data Extended Data Fig. 8
Unprocessed western blots and gels.
Rights and permissions
About this article

Cite this article
DeFalco, T.A., Anne, P., James, S.R. et al. A conserved module regulates receptor kinase signalling in immunity and development. Nat. Plants 8, 356–365 (2022). https://doi.org/10.1038/s41477-022-01134-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41477-022-01134-w
This article is cited by
-
A phosphoinositide hub connects CLE peptide signaling and polar auxin efflux regulation
Nature Communications (2023)
-
A network of CLAVATA receptors buffers auxin-dependent meristem maintenance
Nature Plants (2023)
