
Abstract
The chromatin modifier GRAIN WEIGHT 6a (GW6a) enhances rice grain size and yield. However, little is known about its gene network determining grain size. Here, we report that MITOGEN-ACTIVED PROTEIN KINASE 6 (OsMAPK6) and E3 ligase CHANG LI GENG 1 (CLG1) interact with and target GW6a for phosphorylation and ubiquitylation, respectively. Unexpectedly, however, in vitro and in vivo assays reveal that both of the two post-translational modifications stabilize GW6a. Furthermore, we uncover two major GW6a phosphorylation sites (serine142 and threonine186) targeted by OsMAPK6 serving an important role in modulating grain size. In addition, our genetic and molecular results suggest that the OsMAPK6-GW6a and CLG1-GW6a axes are crucial and operate in a non-additive manner to control grain size. Overall, our findings identify a previously unknown mechanism by which phosphorylation and ubiquitylation non-additively stabilize GW6a to enhance grain size, and reveal correlations and interactions of these posttranslational modifications during rice grain development.
Similar content being viewed by others


Introduction
For rice (Oryza sativa), a major cereal crop worldwide, seed size is a crucial component trait of grain quality and yield. In the past two decades, many genetic factors determining grain size in rice have been identified, which were categorized into several pathways such as those mediated by plant hormones, ubiquitin proteasome, mitogen-activated protein kinase (MAPK), G proteins and transcriptional regulators1,2. In particular, MAPK cascades are highly conserved signaling modules in many species across eukaryotes that coordinate environmental stimuli and developmental inputs to alter substrate activities via the sequential phosphorylation: MAKKKs (MAPK kinase kinases) phosphorylate and activate downstream MAPKKs (MAPK kinases), which in turn phosphorylate and activate MAPKs3,4,5. It is worth noting that a genetically defined OsMKKK10-OsMKK4-OsMAPK6 pathway has recently been reported to play an important role in the regulation of rice grain size, in which each of the three components positively regulates grain size by promoting cell proliferation in spikelet hulls6,7,8,9. Furthermore, one regulator of the MAPK cascade is MAPK PHOSPHATASE 1 (GRAIN SIZE AND NUMBER 1, GSN1), which interacts with and deactivates OsMAPK6 via dephosphorylation to negatively regulate grain size10. The Rho-family GTPase OsRac1 could positively regulate grain size through enhancement of phosphorylation and stabilization of OsMAPK611. In addition, OsWRKY53 is a direct substrate of the OsMKKK10-OsMKK4-OsMAPK6 pathway, in which OsMAPK6 phosphorylates OsWRKY53 to increase its transcriptional activity to function in the control of grain size12,13,14,15. These findings greatly advanced our understanding of the MAPK pathway in the regulation of grain size; however, to date only a few of direct MAPK substrates that are involved in controlling the trait have been reported.
Meanwhile, ubiquitylation is well-known for targeting cellular proteins for the 26S proteasome-mediated degradation and homeostasis, in addition to its other functions such as modulation of protein interactions, subcellular distribution, transcription, DNA repair and propagation of transmembrane signaling, which has been linked to almost every cellular process16,17,18. The modification occurs through the sequential action of three types of protein: ubiquitin-activating enzymes (E1s), ubiquitin-conjugating enzymes (E2s), and ubiquitin ligase (E3s)18,19. Interestingly, several E3-participated pathways have been identified in controlling rice grain size; for example, the E3 ubiquitin ligase GRAIN WIDTH ON CHROMOSOME 2 (GW2) ubiquitinates and destabilizes the glutaredoxin protein WIDE GRAIN 1 (WG1) that interacts with and represses the transcriptional activity of OsbZIP47 by recruiting the transcriptional co-repressor ABERRANT SPIKELET AND PANICLE 1 (ASP1)20,21. Another significant E3 ligase CHANG LI GENG 1 (CLG1, also called HAEMERYTHRIN MOTIF-CONTAINING RING-AND ZINC-FINGER PROTEIN 2, OsHRZ2) targets the Gγ protein GRAIN SIZE 3 (GS3) for ubiquitylation and subsequent degradation through the endosome pathway to control rice grain size22,23. Similarly, recent studies suggested that the E3 ubiquitin ligase DECREASED GRAIN SIZE 1 (DGS1) interacts with and ubiquinates the brassinosteroid receptor BRASSINOSTEROID INSENSITIVE 1 (BRI1) facilitating its endoplasmic reticulum-associated degradation to increase grain size24,25. In addition, our recent finding has revealed that the ubiquitin receptor HOMOLOG OF DA1 ON RICE CHROMOSOME 3 (HDR3) increases the K63-linked ubiquitylation of and stabilizes GRAIN WEIGHT 6a (GW6a) to positively regulate rice grain size26, although the corresponding E3 ubiquitin ligase(s) that collaborate(s) with HDR3 has still remained elusive.
GW6a is a positive modulator of grain size and yield, which encodes a histone H4 acetyltransferase (chromatin modifier), functioning presumably through the regulation of transcription27. However, its gene network controlling grain size has remained largely unknown. Here, we coincidentally identified a mitogen-activated protein kinase (OsMAPK6) and a RING-type E3 ligase (CLG1) as another two GW6a interacting proteins in a yeast two-hybrid screening assay. Furthermore, our in vitro and in vivo biochemical evidence suggested that OsMAPK6 and CLG1 targeted GW6a, respectively, for protein phosphorylation and ubiquitylation. It was unexpected that, however, both of the posttranscriptional modifications facilitated the stabilization of GW6a, which was different from the known mechanisms for the OsMAPK6-mediated control of grain size in rice. Meanwhile, our genetic and corresponding molecular evidence revealed that both of the viable OsMAPK6-GW6a and CLG1-GW6a axes play crucial roles in the regulation of rice grain size. In addition, we showed that the OsMAPK6-mediated phosphorylation and CLG1-mediated ubiquitylation of GW6a stabilize the substrate protein in a non-additive manner, and the triggered effects have non-additively altered grain growth in rice. Collectively, our findings revealed a previously unknown mechanism where the integration of several posttranscriptional modifications was utilized to fine-tune grain size in rice.
Results
Identification of mitogen-activated protein kinase OsMAPK6 and RING-type E3 ligase CLG1 as interacting partners of GW6a
We initially sought to identify additional interacting proteins of GW6a using a yeast two-hybrid (Y2H) screening strategy. Ultimately, one of the putative GW6a interacting partners we identified was OsMAPK6, which was known as a positive modulator of rice grain size8,9,10,15. To verify the interaction, we resorted to pull-down assays. The Escherichia coli-produced fusion proteins GST-OsMAPK6 (or GST instead) and His-GW6a were incubated in a pull-down buffer along with anti-GST resin beads. Obviously, His-GW6a was pulled down by GST-OsMAPK6 (detected with an antibody to GW6a), but not by GST (Fig. 1a). We also confirmed the interaction using co-immunoprecipitation (Co-IP) assays in Nicotiana benthamiana (tobacco) leaves. Proteins were extracted from the leaves transiently co-expressing GW6a-Myc and OsMAPK6-GFP (or GFP instead); upon IP with an anti-GFP antibody, immunoblotting with anti-GFP and anti-Myc antibodies suggested that GW6a-Myc was immunoprecipitated by OsMAPK6-GFP, but not by GFP (Fig. 1b). Furthermore, we performed bimolecular flurescence complementation (BiFC) assays to verify the interaction, and observed strong fluorescence signals in the tobacco leaves co-expressing C-terminal part of yellow fluorescent protein fused with OsMAPK6 (OsMAPK6-cYFP) and N-terminal part of yellow fluorescent protein fused with GW6a (nYFP-GW6a), but did not in those leaves co-expressing OsMAPK6-cYFP and nYFP, or nYFP-GW6a and cYFP (Fig. 1c). Thus, these results suggest that OsMAPK6 interacts with GW6a.

a Pull-down assays showing that OsMAPK6 interacts with GW6a in vitro. PD pulling down, IB immunoblotting. b OsMAPK6-GFP interacts with GW6a-Myc in vivo. Total protein from Nicotiana benthamiana leaves co-expressing OsMAPK6-GFP (or GFP) and GW6a-Myc was immunoprecipitated by anti-GFP agarose beads and the immunoblots were probed using anti-Myc and anti-GFP antibodies. IP immunoprecipitation. c BiFC assays in N. benthamiana (tobacco) leaves of the interaction between nYFP-GW6a (nYFP) and OsMAPK6-cYFP (cYFP). The split YFP system was used. H2B-mCherry is a nucleus-localized marker. d Pull-down assays showing that CLG1 interacts with GW6a in vitro. e CLG1-GFP interacts with GW6a-Myc in tobacco leaves. f BiFC assays in rice protoplast cells of the interaction between nYFP-GW6a (nYFP) and CLG1-cYFP (cYFP). YFP fluorescence and autofluorescence signals from chloroplasts were pseudo-colored as yellow and red, respectively. The experiments in (a–f) were repeated at least two times will similar results.
In the meantime, we also paid special attention to another candidate GW6a interacting partner that is equivalent to the RING-type E3 ligase CLG1 (also called OsHRZ2)22,23. Similarly, we tested the interaction in a pull-down assay; as anticipated, CLG1-Myc was pulled down by GST-GW6a, but not by GST (Fig. 1d). The interaction was also verified using Co-IP assays, the results of which suggesting that GW6a-Myc was immunoprecipitated by CLG1-GFP, but not by GFP (Fig. 1e). Further BiFC assays revealed strong fluorescence signals in rice protoplasts co-expressing CLG1-cYFP and nYFP-GW6a, but did not in the corresponding controls (Fig. 1f). These results thus suggest that CLG1 is another interacting partner of GW6a. Collectively, we identified the mitogen-activated protein kinase OsMAPK6 and RING-type E3 ligase CLG1 that interact with GW6a.
OsMAPK6 phosphorylates GW6a on Ser142 and Thr186 residues to control grain size
We inferred that GW6a might be a substrate of OsMAPK6. To test this idea, we performed kinase assays in tobacco leaves, and the following results indicated that co-expressing GFP fused with constitutively active version of OsMAPK6 (CA-OsMAPK6-GFP)9 and GW6a-Myc could evidently increase the phosphorylation level of GW6a-Myc relative to that of the control (Fig. 2a). Consistent with the results, although in vitro kinase assays using a phos-tag SDS-PAGE gel of a low concentration (8%) with short exposure showed that MBP-GW6a cannot be visibly phosphorylated by GST tagged CA-OsMAPK6 (upper lane), the corresponding long exposure revealed that the phosphorylation seemingly occurred (lower lane), and MBP tagged N-terminal part of GW6a protein (amino acids 1-191; MBP-nGW6a) could be unambiguously phosphorylated independent of the addition of His-CA-OsMKK4, whereas MBP-cGW6a (amino acids 192-419) could not (Fig. 2b). In particular, in vitro kinase assays using a phos-tag SDS-PAGE gel of a high concentration (12%) showed that GW6a can be visibly phosphorylated by OsMAPK6, and calf intestinal alkaline phosphatase (CIAP) treatment could greatly compensate for the enhancement of GW6a phosphorylation (Fig. 2c). As we have anticipated, CA-OsMAPK6-GFP also enhanced the phosphorylation level of nGW6a-Myc in tobacco leaves (Fig. 2d). Accordingly, CIAP treatment could greatly compensate for the enhancement of His-nGW6a phosphorylation caused by MBP-CA-OsMAPK6 (Fig. 2e). Hence, OsMAPK6 has the capability to phosphorylate GW6a.

a Constitutively active version of OsMAPK6 (CA-OsMAPK6) phosphorylates GW6a. The CA-OsMAPK6-GFP (or GFP) and GW6a-Myc fusion proteins were expressed in tobacco leaves as indicated. IB immunoblotting. b GST-CA-OsMAPK6 phosphorylates MBP-nGW6a and possibly MBP-GW6a in vitro. The red arrowheads indicate phosphorylated MBP-nGW6a or phosphorylated MBP-GW6a. c GST-CA-OsMAPK6 phosphorylates MBP-GW6a in vitro, the biochemical effect of which was inhibited by calf intestinal alkaline phosphatase (CIAP) treatment for 1 h. The red arrowheads indicate MBP-GW6a (lower panel) or phosphorylated MBP-GW6a. d CA-OsMAPK6-GFP phosphorylates nGW6a-Myc in tobacco leaves. e MBP-CA-OsMAPK6 phosphorylates His-nGW6a. The red arrowhead indicates phosphorylated His-nGW6a. f Schematic diagram showing the viable GW6a phosphorylation sites by Mass spectrometry. g MBP-CA-OsMAPK6 substantially loses the ability to phosphorylate His-nGW6aS142/T186D in vitro. The red bracket indicates shifted bands of phosphorylated His-nGW6a, His-nGW6aS142D, His-nGW6aT186D, and His-nGW6aS142/T186D in phos-tag SDS-PAGE gel. h CA-OsMAPK6-GFP markedly loses the ability to phosphorylate nGW6aS142/T186D-Myc in tobacco leaves. i CA-OsMAPK6-GFP greatly loses the ability to phosphorylate nGW6aS142/T186A-Myc in tobacco leaves. The experiments in (a–e and g–i) were repeated at least two times will similar results.
Furthermore, we examined the GW6a phosphorylation sites targeted by OsMAPK6, and performed an in vitro kinase assay incubating His-nGW6a, GST-CA-OsMKK4, and MBP-CA-OsMAPK6; the reaction mixture was then subjected to liquid chromatography-tandem mass spectrometry. We thus identified two viable phosphorylation sites of GW6a (Ser142 and Thr186) located within the GNAT domain (Fig. 2f, Supplementary Fig. 1). To test the results, we mutated these amino acid residues to generate His tagged nGW6aS142/T186D (the two amino acid residues mutated into aspartic acid to mimic the phosphorylated nGW6a), nGW6aS142D and nGW6aT186D, and examined their effects on GW6a phosphorylation utilizing in vitro kinase assays; the following results hinted that these mutations have distinctly altered the phosphorylation (Fig. 2g). Consistently, we found that the nGW6a phosphorylation levels of protein extracts of the tobacco leaves transiently co-expressing CA-OsMAPK6-GFP and nGW6aS142/T186D-Myc were markedly reduced compared with those co-expressing CA-OsMAPK6-GFP and nGW6a-Myc (Fig. 2h). In addition, we also mutated both of the amino acid residues into an alanine and co-expressed Myc tagged nGW6aS142/186A (or nGW6a instead) and CA-OsMAPK6-GFP in tobacco leaves, and found that OsMAPK6 also greatly losses the ability to phosphorylate the mutant version of nGW6a (Fig. 2i). These observations suggest that Ser142 and Thr186 of GW6a constitute the major sites phosphorylated by OsMAPK6.
These above results suggest that nGW6a, but not cGW6a, was markedly phosphorylated by OsMAPK6 (Fig. 2b, d, e), and coincidentally, we observed that relative to those of non-transgenic plants, the mature grains of transgenic rice plants over-expressing HA tagged nGW6a (OE-HA-nGW6a) and HA-cGW6a have increased grain length by over 14% and less than 5%, respectively (Supplementary Fig. 2). On the basis of these observations, we next investigated whether the phosphorylation of GW6a is involved in grain size control, and produced transgenic rice lines over-expressing Myc tagged nGW6aS142/T186D (OE-nGW6aS142/T186D-Myc) and nGW6aS142/T186A (the two amino acid residues mutated into alanine to mimic unphosphorylation; OE-nGW6aS142/T186A-Myc). The OE-nGW6aS142/T186D-Myc plants produced grains obviously larger than, whereas those of OE-nGW6aS142/T186A-Myc set seeds almost comparable in size to did the control, although the two types of transgenic lines harbored similar transcriptional expression (Supplementary Fig. 3). These observations suggest that phosphorylation of GW6a by OsMAPK6 is involved in the regulation of grain size.
GW6a is an ubiquitylation substrate of CLG1
We next investigated whether CLG1 was able to ubiquitinate GW6a. For this purpose, we obtained the fusion proteins His-GW6a and MBP-CLG1, and performed in vitro E3 ubiquitin ligase activity assays. In the presence of His-ubiquitin (His-Ub), His-E1 and E2, MBP-CLG1 can ubiquitinate His-GW6a (detected by anti-His and anti-Ub), whereas in the absence of any of His-E1, E2 or E3 enzymes, or His-Ub, we did not observe any His-GW6a ubiquitination signals (Fig. 3a). By contrast, MBP-CLG1△R (deletion of the RING domain of CLG1) was deprived of the ability to ubiquitinate GW6a (Fig. 3b). Consistent with these observations, the ubiquitylated GW6a-GFP levels of protein extract of rice protoplast cells co-expressing CLG1-Myc and GW6a-GFP were markedly enhanced, whereas those co-expressing CLG1△R-Myc and GW6a-GFP were not (Fig. 3c), echoing a conclusion that CLG1 ubiquitinates GW6a in a RING domain-dependent manner. The ubiquitylation inhibitor PYR-41 exerts an effect to block onset of ubiquitylation28. As expected, treatment of PYR-41 in rice protoplasts could largely inhibit the effect of CLG1 ubiquitylation of GW6a (Fig. 3d).

a In vitro E3 ubiquitin ligase activity assays suggesting that CLG1 has the ability to ubiquitinate GW6a. MBP was used as a negative control. b CLG1ΔR (deletion of the RING domain of CLG1) loses the ability to ubiquitinate GW6a in vitro. c CLG1-Myc has, but CLG1ΔR-Myc loses the ability to ubiquitylate GW6a-GFP in rice protoplasts. d The ubiquitylation inhibitor PYR-41 inhibits the ubiquitination of GW6a-GFP by CLG1-Myc in rice protoplast cells. e The ubiquitylation levels of GW6a are enhanced in the transgenic young panicles of over-expression of CLG1. Total proteins from young panicles of OE-CLG1 and the non-transgenic control (‘Nip’) were extracted, and immunoprecipitated with an antibody to GW6a. The ubiquitylation of GW6a levels are detected by following immunoblotting with anti-Ub and anti-GW6a antibodies. The arrowhead points to the GW6a band. f Enhanced ubiquitination of GW6a in OE-CLG1 seedling leaves relative to that of the ‘Nip’ control. g The ubiquitylation levels of GW6a are enhanced in the young panicles of CLG1p-CLG1-GFP, but not in those of OE-CLG1ΔR-GFP. The arrowhead points to the GW6a band. The experiments in (a–g) were repeated at least two times will similar results.
Supporting the above results, we monitored the ubiquitylation of GW6a in the protein extracts of OE-CLG1 young panicles and seedling leaves, and the following results suggest that over-expression of CLG1 clearly promoted the ubiquitylation levels of GW6a (Fig. 3e, f). Similarly, we found that CLG1p-CLG1-GFP also markedly enhanced the ubiquitylation of GW6a, whereas OE-CLG1△R-GFP did not (Fig. 3g). Thus, we concluded that GW6a is a viable ubiquitylation substrate of CLG1.
Both modifications mediated by OsMAPK6 and CLG1 facilitate GW6a stabilization
We were curious to know the biochemical consequences of CLG1 and OsMAPK6-mediated modifications of GW6a. Interestingly, we noted that concomitant with the increased GW6a phosphorylation caused by OsMAPK6, GW6a stabilization was markedly enhanced (Fig. 2a, c). Coincidentally, along with the promoted GW6a ubiquitylation, its stabilization was also boosted by the CLG1 action as evidenced by some in vitro and in vivo assays (Fig. 3c–e, g), suggesting that both the OsMAPK6 and CLG1-mediated modifications of GW6a stabilize the substrate. To reinforce the conclusion, we evaluated the GW6a levels under the circumstance of co-expressing tag fused GW6a and CA-OsMAPK6 (or DN-OsMAPK6 instead, which changes the TEY activation loop motif of OsMAPK6 to AEF, causing loss of the catalytic activity)9,29,30. As expected, the GW6a levels were obviously enhanced by addition of the constitutive active version of CA-OsMAPK6, but not by that of DN-OsMAPK6 in both rice protoplast and tobacco systems (Fig. 4a, b). Supporting these observations, relative to that of ZH11, GW6a accumulation in the protein extracts of transgenic young panicles over-expressing CA-OsMAPK6 (OE-CA-OsMAPK6) was clearly boosted, but that of OE-DN-OsMAPK6 was marginally increased (Fig. 4c). Considering that the mature transgenic rice grains of OE-CA-OsMAPK6 became larger, whereas those of OE-DN-OsMAPK6 were smaller9,15, we inferred that OsMAPK6-mediated phosphorylation of GW6a is essential to stabilize the substrate to control grain size.

a CA-OsMAPK6-Myc, but not DN-OsMAPK6-Myc, substantially promotes the protein accumulation of GW6a-GFP in rice protoplasts. b CA-OsMAPK6-GFP, but not DN-OsMAPK6-GFP, substantially promotes the protein accumulation of GW6a-Myc in tobacco leaves. c The GW6a protein level is enhanced in the young panicles of OE-CA-OsMAPK6, but not in those of OE-DN-OsMAPK6. d The proteins levels of GW6a-Myc (or GW6a-GFP) are substantially enhanced with the addition of CLG1-GFP (CLG1-Myc) compared with those of addition of GFP (Myc) in rice protoplasts. e CLG1-GFP, but not CLG1ΔR-GFP, clearly promotes the protein accumulation of GW6a in transformed tobacco leaves. f In vivo degradation assays of GW6a-Myc by CA-OsMAPK6-GFP or DN-OsMAPK6-GFP in tobacco leaves. Protein extractions are incubated for the indicated time and used for immunoblotting assays. g A normalized plot for degradation of GW6a-Myc in (f). Three independent biological repeats are performed for the analysis. The details of quantification and normalization are described under Methods. h In vivo degradation assays of GW6a-Myc by CLG1-GFP or CLG1ΔR-GFP, or GFP in tobacco leaves. Protein extractions are incubated for the indicated time and used for immunoblotting assays. i A normalized plot for degradation of GW6a-Myc in (h). Three independent biological repeats are performed for the analysis. j Immunobloting analysis of GW6a-Myc amount in the cytoplasm and nuclear components extracted from tobacco leaves co-expressing the indicated GW6a and CA-OsMAPK6 (or DN-OsMAPK6) fusion proteins. k Immunobloting analysis of GW6a-Myc amount in the cytoplasm and nuclear components extracted from tobacco leaves co-expressing the indicated GW6a and CLG1 (or CLG1ΔR) fusion proteins. The experiments in (a–f, h, j, k) were repeated at least three times will similar results.
Similarly, we tested GW6a protein levels under the condition of co-expressing tag fused GW6a and CLG1 (or CLG1△R instead), and observed that GW6a abundance was significantly enhanced by addition of CLG1-GFP (or CLG1-Myc), but not by that of CLG1△R-GFP in both rice protoplast and tobacco systems (Fig. 4d, e). To support these results, we conducted in vivo degradation assays, and found that half-life (T1/2) of GW6a-Myc in tobacco leaves co-expressing GW6a-Myc and CA-OsMAPK6-GFP was over two times that co-expressing GW6a-Myc and DN-OsMAPK6-GFP (Fig. 4f, g). Meanwhile, we found that T1/2 of GW6a-Myc in tobacco leaves with addition of CLG1△R-GFP was almost half that with CLG1-GFP addition, which was comparable to that co-expressing GW6a-Myc and GFP (Fig. 4h, i). In addition, we investigated the changes of GW6a amount in cell components, and immunoblotting revealed that the altered GW6a-Myc levels arose predominantly in the nuclear fraction (Fig. 4j, k), which is consistent with a previous observation26.
OsMAPK6-GW6a and CLG1-GW6a define previously unknown genetic axes to control grain size through alteration of cell number
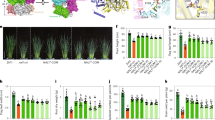
We next examined the genetic relationships between OsMAPK6 and GW6a in grain size control. For this purpose, we crossed OE-GW6a-Myc-1 with OE-CA-OsMAPK6 to achieve OE-CA-OsMAPK6/OE-GW6a-Myc-1 (Fig. 5a, b). The following phenotypic analyses showed that grain length and weight of OE-CA-OsMAPK6/OE-GW6a-Myc-1 were significantly longer and heavier than those of OE-GW6a-Myc-1, but shorter and lighter than those of OE-CA-OsMAPK6 (Fig. 5c, d). In the meantime, we also obtained OE-DN-OsMAPK6/OE-GW6a-Myc-2 by a genetic cross (Fig. 5e, f). As expected, we observed that grain length and weight of OE-DN-OsMAPK6/OE-GW6a-Myc-2 was between those of OE-GW6a-Myc-2 and OE-DN-OsMAPK6 (Fig. 5g, h), hinting that OE-GW6a-Myc could suppress the inhibitory effect on grain size and weight by Os-DN-OsMAPK6. Thus, we conclude that OsMAPK6 most likely functions in the same genetic pathway as and acts upstream of GW6a to control grain size and weight.

a Mature grains of ‘ZH11’, OE-CA-OsMAPK6 (pACTIN::CA-OsMAPK6), OE-CA-OsMAPK6/OE-GW6a-Myc-1, and OE-GW6a-Myc-1 (pSUPER::GW6a-Myc). Scale bar = 5 mm. b Relative expression of OsMAPK6 and GW6a in plants shown in (a) (n = 3). The UBQ5 was used as the internal reference gene. c, d Phenotypic comparisons of grain length (n = 4), grain width (n = 10), grain thickness (n = 10), and 1,000-grain weight (n = 3) in (a). e Mature grains of ‘ZH11’, OE-DN-OsMAPK6 (pACTIN::DN-OsMAPK6), OE-DN-OsMAPK6/OE-GW6a-Myc-2, and OE-GW6a-Myc-2. Scale bar = 5 mm. f Relative expression of OsMAPK6 and GW6a in plants shown in (e) (n = 3). The UBQ5 was used as the internal reference gene. g, h Phenotypic comparisons of grain length (n = 4), grain width (n = 10), grain thickness (n = 10), and 1000-grain weight (n = 3) in (e). Data in (b–d and f–h) represent mean ± SD, and P values were obtained by one-way Student’s t-test compared with the corresponding controls.
CLG1 is a positive modulator of rice grain size (length), and transgenic rice with over-expression of a mutated form of CLG1 substituting three key amino acids in its RING domain set significant smaller grains in size (a dominant negative mutant)23. Consistent with these observations, our transgenic rice plants containing over-expression of CLG1 or CLG1 native promoter-driven CLG1-GFP fusion (CLG1p-CLG1-GFP-1) produced markedly increased grain length and weight (Supplementary Fig. 4a–d). In contrast, transgenic rice plants containing OE-CLG1△R-GFP set significantly shorter and lighter grains (Supplementary Fig. 4e–h). We then investigated the genetic relationship between CLG1 and GW6a in the regulation of rice grain size, and generated the hybrid OE-CLG1△R-GFP-1/OE-GW6a-Flag-1 through a genetic cross (Fig. 6a, b). As expected, we observed that the mature rice grains of OE-CLG1△R-GFP-1/OE-GW6a-Flag-1 were significantly larger and heavier than those of OE-CLG1△R-GFP-1 but smaller and lighter than those of OE-GW6a-Flag-1 (Fig. 6c, d). We also found that the mature grains of the hybrid CLG1p-CLG1-GFP-1/OE-GW6a-Flag-2 did not exhibit additive effects relative to those of CLG1p-CLG1-GFP-1 and OE-GW6a-Flag-2 (Fig. 6e–h). We also found that knockout of GW6a (cg-gw6a) significantly reduced grain length, although it profoundly harmed grain filling; obviously, the grain size (length) of cr-gw6a/CLG1p-CLG1-GFP-1 was significantly longer than that of cr-gw6a, but slightly shorter than that of CLG1p-CLG1-GFP-1 (Supplementary Fig. 5). Taken these results together, we concluded that CLG1 and GW6a most likely function in the same genetic pathway to control grain size.

a Mature grains of Nip, OE-GW6a-Flag-1 (p35S::GW6a-Flag), OE-CLG1∆R-GFP-1, and OE-CLG1∆R-GFP-1/OE-GW6a-Flag-1. Scale bar = 5 mm. b Relative transcriptional expression of GW6a and CLG1 in Nip and the transgenic young panicles shown in (a). The UBQ5 was used as the internal reference gene (n = 3). Comparisons of grain length (n = 4), grain width (n = 12), grain thickness (n = 12) (c), and 1,000-grain weight (n = 4) (d) between Nip and OE-GW6a-Flag-1, OE-CLG1∆R-GFP-1, and OE-CLG1∆R-GFP-1/OE-GW6a-Flag-1. Data in (b–d) represent mean ± SD, and P values were obtained by one-way Student’s t-test compared with the corresponding controls. e Mature grains of Nip, OE-GW6a-Flag-2, CLG1p-CLG1-GFP-1, and CLG1p-CLG1-GFP-1/OE-GW6a-Flag-2. Scale bar = 5 mm. f Relative transcriptional expression of GW6a and CLG1 between Nip and the transgenic young panicles shown in (e). The UBQ5 was used as the internal reference gene (n = 3). Comparisons of grain length (n = 4), grain width (n = 10), grain thickness (n = 10) (g), and 1000-grain weight (n = 4) (h) between Nip and OE-GW6a-Flag-2, CLG1p-CLG1-GFP-1, and CLG1p-CLG1-GFP-1/OE-GW6a-Flag-2. Data in (f–h) represent mean ± SD, and P values were obtained by one-way Student’s t-test compared with the corresponding controls.
Previous studies revealed that both OsMAPK6 and GW6a regulate grain length through change of cell number in spikelet hulls8,27. These observations prompted us to examine whether CLG1 regulates grain length through alteration of cell number and/or cell size. To this end, we performed comparative analyses of the central parts of outer epidermal cells of mature grains between OE-CLG1-1, CLG1p-CLG1-GFP-2, and OE-CLG1△R-GFP-2 and the corresponding control by scanning electron microscopy (Supplementary Fig. 6a, c). As expected, in the grain-length orientation, compared with those of non-transgenic control (Nip), the cell numbers of OE-CLG1-1 and CLG1p-CLG1-GFP-2 grains were significantly increased, with cell sizes unchanged (Supplementary Fig. 6b, d). By contrast, the cell numbers of OE-CLG1△R-GFP-2 grains exhibited a significant decrease relative to those of the control (Supplementary Fig. 6e, f). These results suggest that like GW6a and OsMAPK6, CLG1 regulates grain size (length) through alteration of cell number in spikelet hulls.
OsMAPK6 phosphorylation and CLG1 ubiquitylation of GW6a regulate grain size in a non-additive manner
To further test the relationship between CLG1-GW6a and OsMAPK6-GW6a axes in the regulation of grain size, we created hybrids between OE-GW6a-Myc, OE-CA-OsMAPK6 and OE-CLG1-GFP and the triple transgenic lines (Fig. 7a). Phenotypic analyses revealed that compared with those of single and double over-expressions, the grain size and weight of the triple crossed lines did not exhibit additive effects (Fig. 7b, c), suggesting that the CLG1-GW6a and OsMAPK6-GW6a axes might correlate with each other rather than operate independently in the regulation of grain size. To support this assumption, we examined GW6a protein amounts in tobacco leaves transiently co-expressing a combination of GW6a-Myc and GFP alone or GFP fused with CLG1 or CA-OsMAPK6; as expected, the increase of GW6a amount did not exhibited additive effect with simultaneous CLG1-GFP and CA-OsMAPK6-GFP addition relative to CLG1-GFP or CA-OsMAPK6-GFP addition instead (Fig. 7d). We also obtained a very similar result in rice protoplast cells co-expressing the indicated fusion proteins (Fig. 7e). In addition, we evaluated GW6a protein levels in transgenic rice plants of the single, double, and the triple over-expression of GW6a, CLG1, and OsMAPK6, and found that, as expected, the protein amount in the triple over-expression did not exhibit additive effect (Fig. 7f).

a Mature grains of ZH11, OE-GW6a-Myc, OE-CA-OsMAPK6, OE-CLG1-GFP (pSUPER::CLG1-GFP) and their hybrid progeny. Scale bar = 5 mm. Comparisons of grain length (n = 4), grain width (n = 12), grain thickness (n = 12) (b), and 1000-grain weight (n = 3) (c) between ZH11 and the indicated transgenic lines shown in (a). Data in (b, c) represent mean ± SD, and P values were obtained by one-way Student’s t-test compared with the corresponding controls. d The protein levels of GW6a-Myc in tobacco leaves transiently co-expressing CLG1-GFP and/or CA-OsMAPK6-GFP (or GFP instead). Image J software was used to quantify the protein levels of GW6a-Myc. e The protein levels of GW6a-Myc in rice protoplasts transiently co-expressing CLG1-GFP and/or CA-OsMAPK6-GFP (or GFP instead). Image J software was used to quantify the protein levels of GW6a-Myc. f The protein levels of GW6a in transgenic OE-GW6a-Myc, OE-GW6a-Myc/OE-CA-OsMAPK6, OE-CLG1-GFP/OE-GW6a-Myc, and OE-GW6a-Myc/OE-CA-OsMAPK6/OE-CLG1-GFP rice plants. Image J software was used to quantify the protein levels of GW6a-Myc. The experiments in (d-f) were repeated at least three times will similar results.
GW6a regulates grain size presumably through regulation of transcription26,27. We inferred that the OsMAPK6-GW6a and CLG1-GW6a effects could be seen in the transcriptional expression of their co-targeted genes. For this purpose, we compared the transcriptome of the transgenic OE-GW6a-Myc, OE-CA-OsMAPK6, and CLG1p-CLG1-GFP young panicles with that of the wild type. Ultimately, we identified 27 up-regulated genes shared by the three lines (Fig. 8a, b). It was worth noting that among these genes, OsWRKY53 has been characterized as an important modulator of rice grain size through integration of BR signaling and MAPK pathway and OsERF64 encodes an APETALA2/ethylene-response element binding protein14,15. Considering that APETALA2-type protein could play a key role in regulating rice grain size and yield31, we selected OsWRKY53 and OsERF64 as target genes and validated by quantitative real-time PCR analysis that the transcriptional levels of the two genes were evidently up-regulated in these over-expression lines (Fig. 8c, d). We next examined the transcriptional levels of OsWRKY53 and OsERF64 in the young panicles of the single, double, and triple over-expression of GW6a, CLG1, and OsMAPK6, and unexpectedly, however, we found that the triple over-expression line did not exhibit additive effects on the expression of OsERF64 but exhibited a different effect on that of OsWRKY53 (Fig. 8e, f). In support of these observations, we transiently co-expressed a combination of tag fused GW6a, CA-OsMAPK6 and CLG1 proteins in rice protoplasts and examined the transcriptional levels of OsERF64 and OsWRKY53; consistent with above observations, simultaneously expressing the three proteins did not exert additive effects on the expression of OsERF64 but had a different influence on that of OsWRKY53 (Fig. 8g, h).

a Venn diagrams showing the overlap of genes up-regulated in OE-GW6a-Myc, OE-CA-OsMAPK6, and CLG1p-CLG1-GFP compared to the corresponding WT as indicated. b Heatmap analysis of 27 up-regulated genes shared by OE-GW6a-Myc, OE-CA-OsMAPK6, and CLG1p-CLG1-GFP versus the corresponding WT. Relative transcriptional expression of OsERF64 (c) and OsWRKY53 (d) in WT, OE-GW6a-Myc, OE-CA-OsMAPK6, and CLG1p-CLG1-GFP young panicles as indicated in (c). Data are mean ± SD. Three biological replicates are performed for each assay. Relative transcriptional expression of OsERF64 (e) and OsWRKY53 (f) in young panicles of ZH11 (WT), and the single, double, and triple over-expression of GW6a, OsMAPK6 and CLG1 as indicated in (e). Data are mean ± SD. Three biological replicates are performed for each assay. Relative transcriptional expression of OsERF64 (g) and OsWRKY53 (h) in rice protoplast cells expressing the Myc tag or a combination of fusion proteins as indicated in (g). Data are mean ± SD. Three biological replicates are performed for each assay.
Discussion
In this current study, we have identified and defined the genetic regulatory axes OsMAPK6-GW6a and CLG1-GW6a that modulate grain size in a non-additive manner (Fig. 7). Mechanistically, the mitogen-activated protein kinase OsMAPK6 and E3 ligase CLG1 directly interacted with and targeted GW6a for phosphorylation and ubiquitylation, respectively and coincidentally, both of the modifications facilitated the stabilization of GW6a (Fig. 4). It was noteworthy that the OsMAPK6 phosphorylation of GW6a was involved in the regulation of grain size evidenced by the transgenic analyses in rice (Supplementary Fig. 3). In addition, consistent with the non-additive roles of the genetic axes in controlling grain size, the two posttranslational modifications exerted non-additive impact on the GW6a protein stability, and as well, on the transcriptional expression of their co-regulated downstream genes (Figs. 7, 8). Indeed, numerous proteins can be modified by multiple different types of posttranslational modifications whose effects on the biological functions of proteins could be synergistic or antagonistic, thus regulating distinct biological outcomes32,33. We have also performed pull-down assays to test whether phosphorylation of GW6a by OsMAPK6 affects its interaction with CLG1, by incubating the E. coli-produced GST fused GW6a, GW6aS142/T186A and GW6aS142/T186D with CLG1-Myc. Upon IP with GST, immunoblot assays using the anti-GST and anti-Myc antibodies suggested that compared with GST-GW6a, GST-GW6aS142/T186D exhibits much stronger binding to CLG1-Myc, whereas GST-GW6aS142/T186A has a much weaker binding (Supplementary Fig. 7), hinting that phosphorylation of GW6a by OsMAPK6 facilitates its interaction with CLG1, which presumably has a positive effect on the GW6a ubiquitylation by CLG1. Nevertheless, our findings have revealed correlations and interactions of multiple posttranslational modifications, i.e., protein phosphorylation, ubiquitylation and acetylation during rice grain development. However, to reveal the detailed mechanisms of how the viable interplay between OsMAPK6-mediated phosphorylation and CLG1-mediated ubiquitylation of GW6a operates to fine-tune grain size would be a considerable challenge, which awaits future experimentations.
Posttranslational modifications, including protein phosphorylation and ubiquitination, play pivotal roles in regulating the stability, activity, subcellular localization, and interaction of the proteins being modified33,34. Especially, so far several substrates of OsMAPK6 have been identified in rice, which are intimately associated with the almighty functions of OsMAPK6 in plant growth and development. For instance, OsWRKY53 positively regulates brassinosteroid signaling and grain size (length) in rice, the encoded protein of which was targeted by OsMAPK6 for phosphorylation that results in the enhancement of its transcriptional activity14. Similarly, OsMAPK6 also phosphorylates the finger transcription factor DST to enhance its transcriptional activation of OsCKX2 to regulate spikelet number through modulation of cytokinin metabolism35. Furthermore, OsMAPK6 phosphorylates the raf-like kinase OsEDR1 at S861 that destabilizes OsEDR1 ultimately leading to enhanced resistance to rice disease36. In contrast, we showed that OsMAPK6 phosphorylates GW6a at S142 and T186 that facilitates the stability of GW6a to control grain size (Figs. 2, 4). Nevertheless, we have revealed a OsMAPK6-mediated mechanism for grain size control in rice.
In addition to the OsMAPK6-mediated phosphorylation, several GLYCOGEN SYNTHASE KINASE (GSK)-centered phosphorylation axes were described to play an important role in the regulation of rice grain size. In rice, the GSK3-like family has a total of 9 members that were classified into four major groups37. Of these members, OsGSK2, the central negative modulator of brassinosteroid signaling, interacts with and phosphorylates OVATE FAMILY PROTEIN 3 (OFP3) and MEI2-LIKE PROTEIN4 (OML4) to facilitate the protein stability of the modified substrates, whereas it phosphorylates and destabilizes DLT, OFP8, and SMOS1/RLA138,39,40,41,42. OsGSK2 also interacts with the plant-specific transcription factor GROWTH REGULATING FACTOR 4 (OsGRF4) (also called GL2, GRAIN LENGTH ON CHROMOSOME 2) and inhibits its transcription activation activity to regulate grain size43. By contrast, another rice GSK3-like member OsGSK5 (also called TGW3) interacts with OsARF4 and facilitates OsARF4 accumulation, whereas it interacts with and phosphorylates OsIAA10 leading to OsIAA10 degradation by the 26 S proteasome pathway44,45.
Similarly, an early study showed that CLG1 ubiquitylates GS3 triggering its degradation via the endosome pathway23. In contrast, in this study, we observed that CLG1 targets GW6a for ubiquitination to stabilize the substrate (Fig. 4). Based on these above observations, we are surprisingly aware of an explicit fact that the action of these kinases and E3 ligases can steer the diverse substrates towards completely different fates through posttranslational modifications. We have revealed a non-proteolysis-mediated mechanism by which HDR3 increases the K63-linked poly-ubiquitination of and stabilizes GW6a to regulate grain size; by contrast, GW2 ubiquitylates and boosts the degradation of GW6a26. Similar to that of HDR3, the overexpression of CLG1 (OE-CLG1-GFP) has obviously increase of both K63-linked and K48-linked polyubiquitinations of GW6a (Supplementary Fig. 8a). By contrast, the overexpression of GW2 (OE-GW2) has markedly and merely increased the modification of K48-linked polyubiquitinations of GW6a (Supplementary Fig. 8b). We reasoned that the different types of polyubiquitinations of GW6a by CLG1 and GW2 might at least partially contribute to altered GW6a fates of stability. Furthermore, to test whether monoubiquitylation is involved in the regulation of GW6a stabilization, we observed the GW6a ubiquitylation by utilizing the UbK0 variant (relative to the wild-type Ub), in which all seven lysine residues in Ub were changed into arginine to prevent the forming of polyubiquitylation chains28. The following results suggest that both CLG1 and GW2 could monoubiquitylate GW6a in vitro. Moreover, it was worth noting that the three genetic factors CLG1, HDR3, and GW6a are positive regulators of grain length, but GW2 negatively controls the trait, we reasoned that CLG1, HDR3, and GW2 could have a competitive interaction with GW6a. To test the assumption, we performed split firefly luciferase complementation assays of CLG1, HDR3, and GW2. We observed that the GW2 addition exerted a significantly negative effect on both the GW6a-CLG1 and GW6a-HDR3 interactions, while the individual addition of HDR3 and CLG1 did not significantly affect the GW2-GW6a interaction (Supplementary Fig. 9a–d). By contrast, the addition of HDR3 and CLG1, respectively, significantly enhanced the effect of the GW6a-CLG1 and GW6a-HDR3 interactions (Supplementary Fig. 9e, f). Thus, these primary results suggest that HDR3 and CLG1 synergistically enhance their interaction with GW6a, but GW2 behaves as an aggressive competitor of CLG1 and HDR3. However, to figure out the detailed mechanism underlying the discrepancy of biochemical consequences is a great challenge but deserves future investigations.
Methods
Plant materials and growth conditions
The rice (Oryza sativa L.) japonica cultivars ‘Nipponbare’ (‘Nip’) and ‘Zhonghua 11’ (‘ZH11’) were used for transgenic recipient as indicated. The transgenic over-expression lines of OE-GW6a-Flag were described previously26. The ACTIN promoter-driven transgenic over-expression lines of OE-CA-OsMAPK6 and OE-DN-OsMAPK6 were reported earlier9. To construct the 35 S promoter-driven transgenic OE-CLG1 rice plants, the CLG1 coding sequence (CDS) was inserted into pCAMBIA1300-GFP vector. To construct the CLG1 promoter-driven transgenic CLG1p-CLG1-GFP rice plants, the CLG1 CDS and 2-kb promoter were inserted into the BamHI and SpeI sites of pCAMBIA2300-eGFP vector. To construct the SUPER promoter-driven transgenic OE-CLG1△R-GFP rice plants, CLG1△R (deletion of a segment spanning 2077-2208 bp of CLG1 CDS) was inserted into the pSuper1300-GFP vector. To construct the 35 S promoter-driven transgenic OE-HA-nGW6a and OE-HA-cGW6a rice plants, these truncated GW6a sequences were inserted into pCAMBIA1300-GFP vector. To produce the OE-nGW6a-Myc, OE-nGW6aS142/T186D-Myc, and OE-nGW6aS142/T186A-Myc rice plants, the wild-type and mutated GW6a CDS sequences were inserted into the pSuper1300-Myc vector for transgenic assays. To generate the Super promoter-driven transgenic OE-GW6a-Myc and OE-CLG1-GFP rice plants, the CDSs of GW6a or CLG1 were inserted into the pSuper1300-Myc or pSuper1300-GFP vectors. To generate the cr-gw6a mutant, the single-guide RNAs (sgRNAs) was designed and the expression cassette was inserted into pBWA(V)H-Cas9 vector. The primers used in this work are listed in Supplementary Data 1. Transgenic rice plants were grown in experimental fields in Hainan (winter seasons) or Beijing (summer seasons). Nicotiana benthamiana (tobacco) plants were cultivated in growth chambers (16 h light/8 h dark at 25°C).
Pull-down assays
The CDS of GW6a was individually cloned into the BamHI and SalI sites of pGEX-4T-1 and the BamHI and HindIII sites of pET-32a to generate fusion proteins GST-GW6a and His-GW6a. The mutated GW6a CDS was cloned into the BamHI and SalI sites of the pGEX-4T-1 vector to express the fusion proteins GST-GW6aS142/T186D and GST-GW6aS142/T186A. The 4 × Myc and CDS of CLG1 sequences were inserted into the multiple cloning site of pMAL-c2X to express a fusion CLG1-Myc protein. The CDS of OsMAPK6 was cloned into the BamHI and SalI sites of pGEX-4T-1 to express GST-OsMAPK6. The constructs were transformed into Escherichia coli BL21 (DE3) and the recombinant proteins were induced by 0.5-mM IPTG (isopropyl-β-D-thio-galactopyranoside). For pull-down assays, GST-tag purification resin (Beyotime) were incubated with GST or GST-GW6a/GST-OsMAPK6 at 4°C for 1 h, and then incubated with CLG1-Myc/His-GW6a at 4 °C for another 1 h. After incubation, the GST beads were washed thoroughly with PBS buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, 0.05% SDS, 1% Triton X-100), resolved by SDS-PAGE, and detected and probed with anti-GST (EASYBIO), or anti-Myc (TransGen), or anti-GW6a as indicated. Uncropped scans of immunoblotting results are shown in Source data.
Co-IP assays
The CDS of GW6a was amplified and cloned into the HindIII and SpeI sites of the pSuper1300-Myc vector to express a fusion protein GW6a-Myc. The CDS of CLG1 or OsMAPK6 were cloned into pSuper1300-GFP to express CLG1-GFP or OsMAPK6-GFP. Each of the constructs was transformed into Agrobacterium EHA105 cells and co-infiltrated into tobacco leaves as indicated. After 72 h of cultivation, the leaves were harvested and ground in liquid nitrogen. Total proteins were extracted with a buffer (50 mM Tris-MES [PH 8.0], 10 mM EDTA [PH 8.0], 0.5 M sucrose, 1 mM MgCl2, 1 mM DTT, 1 mM PMSF, 1 × complete protease inhibitor cocktail) and incubated with anti-GFP affinity beads (SMART) at 4°C for 3 h. The beads were washed three times with washing Buffer (10 mM Tris-HCl [PH 7.5], 0.5 mM EDTA [PH 8.0], 150 mM NaCl, 0.05% SDS, 1% Triton X-100, 1 mM DTT, 1 mM PMSF, 1 × complete protease inhibitor cocktail). Samples were resolved by SDS-PAGE, detected with anti-GFP (TransGen) and anti-Myc (TransGen). Uncropped scans of immunoblotting results are shown in Source data.
BiFC assays
The CDS of OsMAPK6 was cloned into pBI-2YC to generate a fusion protein OsMAPK6-cYFP. The CDS of GW6a was cloned into pBI-2YN to express nYFP-GW6a. Each construct was transformed into Agrobacterium EHA105 cells and co-infiltrated into tobacco leaves as indicated. Infiltration of EHA105 cells of H2B-mCherry into tobacco leaves serves as a localization signal for the nucleus. The fluorescence signals were examined using a confocal laser scanning microscope (Leica TCS SP5). The CDS of CLG1 was cloned into the SalI and SacI sites of pSY735 to express CLG1-cYFP. The CDS of GW6a was cloned into the SalI and BamHI sites of pSY736 to express nYFP-GW6a. After co-transformation of the constructs, rice protoplasts were incubated overnight and YFP fluorescence signals were observed using a confocal laser scanning microscope (Leica TCS SP5). Proteins from rice protoplasts were extracted as described previously26.
Kinase assays in tobacco leaves
The wild-type (or variant) N-terminal (1–573 bp) and whole CDS of GW6a were cloned into pSuper1300-Myc to express nGW6a-Myc, nGW6aS142/T186D-Myc, nGW6aS142/T186A-Myc, and GW6a-Myc. CA-OsMAPK6 was cloned into the HindIII and SpeI sites of pSuper1300-GFP to express CA-OsMAPK6-GFP. Each of these constructs was transformed into Agrobacterium EHA105 cells and co-infiltrated into tobacco leaves as indicated. Total leave protein was extracted with buffer (50 mM Tris-MES [PH 8.0], 10 mM EDTA [PH 8.0], 0.5 M sucrose, 1 mM MgCl2, 1 mM DTT, 1 mM PMSF, 1 × complete protease inhibitor cocktail) and incubated with anti-Myc Nanobody Agarose Beads (KT HEALTH) at 4 °C for 3 h. The beads were washed for three times with washing buffer (10 mM Tris-HCl [PH 7.5], 0.5 mM EDTA [PH 8.0], 150 mM NaCl, 1 mM DTT, 1 mM PMSF, 1 × complete protease inhibitor cocktail). The samples were analyzed by SDS-PAGE with 50 μM Phos-tag and detected with anti-phospho-(Ser/Thr) (Abcam) or anti-Myc. Uncropped scans of immunoblotting results are shown in Source data.
In vitro kinase assays and phosphopeptide analysis
CA-OsMKK4 was cloned into pGEX-4T-1 or pET-32a to express GST-CA-OsMKK4 or His-CA-OsMKK4, respectively. CA-OsMAPK6 was cloned into pGEX-4T-1 or pMAL-c2X to express GST-CA-OsMAPK6 or MBP-CA-OsMAPK6, respectively. The nGW6aS142/T186D, nGW6aS142D, and nGW6aT186D sequences were amplified with the indicated primers and cloned into pEASY-blunt3 (TransGen, CB301) for sequencing. The cDNA segments of nGW6a, nGW6aS142/T186D, nGW6aS142D, and nGW6aT186D and cGW6a were then cloned into pET-32a or pMAL-c2X to express His-nGW6a, MBP-nGW6a, MBP-cGW6a, respectively. For an in vitro kinase assay, the indicated purified proteins of 1 μg His-CA-OsMKK4 (or GST-CA-OsMKK4), 1.5 μg GST-CA-OsMAPK6 (or MBP-CA-OsMAPK6), and 1.5 μg MBP-nGW6a (or His-nGW6a, His-nGW6aS142D, His-nGW6aT186D, His-nGW6aS142/T186D) and 1.5 μg MBP-cGW6a were incubated in reaction buffer (25 mM Tris-HCl [PH 7.5], 10 mM MgCl2, 1 mM DTT, 200 μM ATP) at 30°C for 1 h. The samples were then analyzed by 8% or 12% SDS-PAGE added with 50 μM Phos-tag (Wako, Phos-tag Acrylamide AAL-107) and detected by immunoblotting with anti-His or anti-MBP antibodies. After in vitro kinase assays, proteins were digested with trypsin (Promega). The digested peptide mixtures were injected into a mass spectrometer (ORBITRAP FUSION LUMOS) for Mass spectrometry analysis. The phosphorylated residues in MBP-nGW6a were found by using Proteome Discoverer software. Uncropped scans of immunoblotting results are shown in Source data.
E3 ubiquitin ligase activity assays
The in vitro E3 ubiquitin ligase activity assay was performed referring to a previous description with some modifications46. The coding sequences of CLG1 and CLG1△R were cloned into pMAL-c2X to express MBP-CLG1 and MBP-CLG1△R, respectively. GW6a was cloned into pET-32a to express His-GW6a. The CDS of GW2 was cloned into pET-32a to express His-GW2. Then, 1 μg E3s (His-GW2, MBP-CLG1 or MBP-CLG1△R), 2 μg His-GW6a, 100 ng His-E1 (R&D Systems), 250 ng E2 (R&D Systems), and 5 μg His-Ub (R&D Systems) or His-UbK0 (all seven lysine residues in ubiquitin were changed to arginine, R&D Systems) were incubated in a 30-μL reaction buffer (50 mM Tris-HCl [pH 7.5], 10 mM MgCl2, 2 mM DTT, and 5 mM ATP) at 30 °C for 2 h. Polyubiquitinated proteins were detected by immunoblotting with anti-Ub, anti-MBP, anti-His, and anti-GW6a antibodies.
To examine CLG1 ubiquitylation of GW6a in rice protoplasts, pSuper1300-GW6a-GFP (expressing GW6a-GFP) was co-transformed with pSuper1300-CLG1△R-Myc (expressing CLG1△R-Myc) or pSuper1300-CLG1-Myc (expressing CLG1-Myc) into rice protoplasts. Total proteins were extracted and immunoprecipitated with anti-GFP affinity beads (a final concentration of 50 μM PYR-41 (Sigma) was applied as shown). The beads were then washed 3 times with washing buffer (10 mM Tris-HCl [PH 7.5], 0.5 mM EDTA [PH 8.0], 150 mM NaCl, 1 mM DTT, 1 mM PMSF, 1 x complete protease inhibitor cocktail). Ubiquitylation was detected by immunoblotting with anti-Ub.
To analyze the CLG1 or GW2 ubiquitination of GW6a in transgenic samples, total proteins from 1 g of seedling leaves or young panicles of OE-CLG1, CLG1p-CLG1-GFP, OE-CLG1△R-GFP, and OE-GW2 were extracted for immunoprecipitation assays as indicated. The rProtein A/G magnetic beads (SMART) of 50 μL conjugated with 10 μg of anti-Ub or anti-GW6a antibodies were incubated with the total protein extracts at 4°C for 3 h. The ubiquitination levels of GW6a were detected by immunoblotting with anti-Ub, anti-UbK48 (CST), anti-UbK63 (CST), or anti-GW6a antibodies. Uncropped scans of immunoblotting results are shown in Source data.
Protein degradation assay
The constructs pSuper1300-CA-OsMAPK6, pSuper1300-DN-OsMAPK6-GFP, pSuper1300-GW6a-Myc, pSuper1300-GFP, pSuper1300-CLG1-GFP, and pSuper1300-CLG1△R-GFP were individually transfected into Agrobacterium EHA105 cells, and co-infiltrated into tobacco leaves as indicated. Total leaves proteins were extracted with degradation buffer (25 mM Tris-HCl [PH 7.5], 10 mM NaCl, 10 mM MgCl2, 5 mM DTT, 4 mM PMSF, and 2 mM ATP) at room temperature. Total GW6a-Myc proteins were then analyzed by SDS-PAGE and detected by immunoblotting with anti-Myc. Coomassie Brilliant Blue staining was used as protein loading control. Three independent biological repeats were performed for the analysis of quantification and normalization of the GW6a protein levels. Uncropped scans of immunoblotting results are shown in Source data.
Nuclear/cytoplasmic fractionation and quantification of proteins
The constructs pSuper1300-GW6a-Myc, pSuper1300-CA-OsMAPK6-GFP, pSuper1300-DN-OsMAPK6-GFP, pSuper1300-CLG1-GFP, pSuper1300-GFP, and pSuper1300-CLG1△R-GFP were co-transformed into tobacco leaves as indicated. Total proteins from transformed leaves were extracted by grinding on rice with Honda buffer (2.5% Ficoll 400, 5% Dextron T40, 0.4 M Sucrose, 25 mM Tris-HCl [PH 7.4], 100 mM MgCl2, 5 mM PMSF, 5 mM DTT, 1 × complete protease inhibitor cocktail) and centrifuged at 1,500 g for 20 min to obtain crude nuclear and cytoplasmic fractions. The supernatant containing cytoplasmic fraction was centrifuged at 16,000 g for 15 min to remove the possible cellular debris. The above pellet containing nuclear fraction was dissolved in ChIP extraction buffer (0.25 M Sucrose, 10 mM Tris-HCl [PH 8.0], 10 mM MgCl2, 1% Triton X-100, 0.1 mM PMSF, 1 mM DTT, 1 x complete protease inhibitor cocktail) and centrifuged at 16,000 g for 10 min at 4°C. Ultimately, the pellet was re-suspended in nuclear lysis buffer (50 mM Tris-MES [PH 8.0], 10 mM EDTA [PH 8.0], 0.5 M sucrose, 1 mM MgCl2, 1 mM DTT, 1 mM PMSF, 1 x complete protease inhibitor cocktail). The samples were analyzed by SDS-PAGE and detected by immunoblotting with anti-Myc and anti-Actin antibodies. Protein levels were quantified using Image J software. Image J software was used for the quantitative analysis of the relative protein abundance of the WB results47. Briefly, for example, in Fig. 4A of our manuscript, changing the format and removing possible background interference of the protein bands of interest was performed in the software. Furthermore, the relative density data for the control (the sample extracted from the rice protoplast cells co-expressing Myc-tag and GW6a-GFP) to the HSP82 (or Actin instead in other Figures) protein integrated density was defined as being equal to 1.0, and the relative density data of another sample (for example, the sample extracted from the rice protoplasts co-expressing CA-OsMAPK6-Myc and GW6a-GFP) divided by the relative control value led to the indicated value (1.5). Similarly, we have obtained the other relative sample values in this study. Uncropped scans of immunoblotting results are shown in Source data.
Split firefly luciferase complementation assay
The CDS of GW6a, CLG1, GW2, and HDR3 were individually cloned into pCAMBIA1300-nLUC or pCAMBIA1300-cLUC to express GW6a-nLUC, CLG1-cLUC, GW2-cLUC, and HDR3-cLUC. The CDS of CLG1, GW2, and HDR3 were cloned into pSuper1300-GFP, respectively, to express CLG1-GFP, GW2-GFP, and HDR3-GFP in tobacco leaves. Each construct was transformed into the Agrobacterium EHA105 cells and co-infiltrated into tobacco leaves as indicated. Tobacco leaves were submerged in a solution with luciferin (YEASEN) for 48 h to capture LUC signals using a CCD imaging system (Tanon).
RNA extraction and quantitative real-time PCR analysis
Total RNA was extracted using the RNA Kit (ZOMANBIO) and reverse transcription was performed using a kit (TIANGEN FastQuart). ACTIN1 was used as an internal reference gene to normalize the expression of GW6a in RT-PCR. Real-time quantitative PCR was performed on a Bio-Rad CFX96 system. Rice UBQ5 was used to normalize the data and relative expression levels were calculated using 2−▵▵Ct method. The primers used in this work are listed in Supplementary Data 1.
Histological analysis
For cytological analysis of cell size and number of spikelet hulls, the central parts of lemmas of mature seeds of OE-CLG1, CLG1p-CLG1-GFP, OE-CLG1△R-GFP and the corresponding non-transgenic control were treated and observed with a scanning electron microscope (Hitachi). The longitudinal cell length was quantified using Image J software. Cell number was decided by grain length and the average cell length.
RNA sequencing and analysis
Young panicles (5 cm in length) of OE-GW6a-Myc, OE-CA-OsMAPK6, CLG1p-CLG1-GFP, and the corresponding wild type were collected and immediately frozen in liquid nitrogen. The RNA sequencing was performed using Illumina NovaSeq 6000. Raw data were trimmed and then mapped to the Nipponbare genome database. Differential expression of genes was analyzed using the DESeq2 program and the genes were considered to be differentially expressed when the FDR was less than 0.05 and the relative fold was higher than 1.5.
Statistics & reproducibility
Data are shown as mean ± SD (standard deviation), calculated using GraphPad Prism 8.0 version. Statistical analysis and number of biologically independent sample (n) were indicated in the figure legends. Significant differences were determined with one-way Student’s t-test by using Microsoft Excel 2021 software.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All materials in this study are available from the corresponding authors upon request. The authors declare that all data supporting the findings of this study are available within the article, supplementary information files, and source data. The primers used in this study are provided as Supplementary Data 1. The RNA-seq data generated in this study has been deposited in NCBI SRA database under accession number PRJNA979276. Sequence data from this study can be found in the GenBank database under the following accession numbers: GW6a (Os06g0650300), CLG1 (Os05g0551000), OsMAPK6 (Os06g0154500), OsMKK4 (Os02g0787300), OsWRKY53 (Os05g0343400), OsERF64 (Os03g0183300), UBQ5 (Os05g0160200), GW2 (Os02g0244100), OsHDR3 (Os03g0267800), ACTIN1 (Os03g0718100). Source data are provided with this paper.
References
Li, N., Xu, R. & Li, Y. Molecular networks of seed size control in plant. Annu. Rev. Plant Biol. 70, 435–463 (2019).
Li, P. et al. Genes and their molecular functions determining seed structure, components, and quality of rice. Rice 15, 18 (2022).
Widmann, C., Gibson, S., Jarpe, M. & Johnson, G. Mitogen-activated protein kinase: conservation of a three-kinase module from yeast to human. Physiol. Rev. 79, 143–180 (1999).
Xu, J. & Zhang, S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 20, 56–64 (2015).
Zhang, M., Su, J., Zhang, Y., Xu, J. & Zhang, S. Conveying endogenous and exogenous signals: MAPK cascades in plant growth and defense. Curr. Opin. Plant Biol. 45, 1–10 (2018).
Duan, P. et al. SMALL GRAIN 1, which encodes a mitogen-activated protein kinase kinase 4, influences grain size in rice. Plant J. 77, 547–557 (2014).
Kishi-Kaboshi, M. et al. A rice fungal MAMP-responsive MAPK cascade regulates metabolic flow to antimicrobial metabolite synthesis. Plant J 63, 599–612 (2010).
Liu, S. et al. OsMAPK6, a mitogen-activated protein kinase, influences rice grain size and biomass production. Plant J. 84, 672–681 (2015).
Xu, R. et al. Control of grain size and weight by the OsMKKK10-OsMKK4-OsMAPK6 signaling pathway in rice. Mol. Plant 11, 860–873 (2018).
Guo, T. et al. GRAIN SIZE AND NUMBER 1 negatively regulates the OsMKKK10-OsMKK4-OsMPK6 cascade to coordinate the trade-off between grain number per panicle and grain size in rice. Plant Cell 30, 871–888 (2018).
Zhang, Y., Xiong, Y., Liu, R., Xue, H. & Yang, Z. The Rho-family GTPase OsRac1 controls rice grain size and yield by regulating cell division. Proc. Natl. Acad. Sci. USA 116, 16121–16126 (2019).
Chujo, T. et al. Overexpression of phosphomimic mutated OsWRKY53 leads to enhanced blast resistance in rice. PLoS One 9, e98737 (2014).
Hu, L. et al. The rice transcription factor WRKY53 suppresses herbivore-induced defenses by acting as a negative feedback modulator of mitogen-activated protein kinase activity. Plant Physiol. 169, 2907–2921 (2015).
Tian, X. et al. Transcription factor OsWRKY53 positively regulates brassinosteroid signaling and plant architecture. Plant Physiol. 175, 1337–1349 (2017).
Tian, X. et al. WRKY53 integrates classic brassinosteroid signaling and the mitogen-activated protein kinase pathway to regulate rice architecture and seed size. Plant Cell 33, 2753–2775 (2021).
Goldstein, G. et al. Isolation of a polypeptide that has lymphocyte-differentiating properties and is probably represented universally in living cells. Proc. Natl. Acad. Sci. USA 72, 11–15 (1975).
Goldberg, A. Nobel committee tags ubiquitin for distinction. Neuron 45, 339–344 (2005).
Metzger, M., Hristova, V. & Weissman, A. HECT and RING finger families of E3 ubiquitin ligases at a glance. J. Cell Sci. 125, 531–537 (2012).
Komander, D. The emerging complexity of protein ubiquitination. Biochem. Soc. Trans. 37, 937–953 (2009).
Song, X., Huang, W., Shi, M., Zhu, M. & Lin, H. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet. 39, 623–630 (2007).
Hao, J. et al. The GW2-WG1-OsbZIP47 pathway controls grain size and weight in rice. Mol. Plant 14, 1266–1280 (2021).
Kobayashi, T. et al. Iron-binding haemerythrin RING ubiquitin ligases regulate plant iron responses and accumulation. Nat. Commun. 4, 2792 (2013).
Yang, W. et al. The RING E3 ligase CLG1 targets GS3 for degradation via the endosome pathway to determine grain size in rice. Mol. Plant 14, 1366–1713 (2021).
Li, J. et al. An endoplasmic reticulum-associated degradation-related E2-E3 enzyme pair controls grain size and weight through the brassinosteroid signaling pathway in rice. Plant Cell 35, 1076–1091 (2023).
Zhu, X. et al. Decreased grain size 1, a C3H4-type RING protein, influences grain size in rice (Oryza sativa L.). Plant Mol. Biol. 105, 405–417 (2021).
Gao, Q. et al. The ubiquitin-interacting motif-type ubiquitin receptor HDR3 interacts with and stabilizes the histone acetyltransferase GW6a to control the grain size in rice. Plant Cell 33, 3331–3347 (2021).
Song, X. et al. Rare allele of a previously unidentified histone H4 acetyltransferase enhances grain weight, yield, and plant biomass in rice. Proc. Natl. Acad. Sci. USA 112, 76–81 (2015).
Ma, X. et al. Ligand-induced monoubiquitination of BIK1 regulates plant immunity. Nature 581, 199–203 (2020).
Bardwell, L. et al. Repression of yeast Ste12 transcription factor by direct binding of unphosphorylated Kss1 MAPK and its regulation by the Ste7 MEK. Genes Dev. 12, 2887–2898 (1998).
Ma, D., Cook, J. & Thorner, J. Phosphorylation and localization of Kss1, a MAP kinase of the Saccharomyces cerevisiae pheromone response pathway. Mol. Biol. Cell 6, 889–909 (1995).
Yu, J. et al. OsLG3 contributing to rice grain length and yield was mined by Ho-LAMap. BMC Biol. 15, 28 (2017).
Csizmok, V. & Forman-Kay, J. Complex regulatory mechanisms mediated by the interplay of multiple post-translational modifications. Curr. Opin. Struct. Biol. 48, 58–67 (2018).
Zhang, Y. & Zeng, L. Crosstalk between ubiquitination and other posttranslational protein modifications in plant immunity. Plant Commun. 1, 100041 (2020).
Willems, P. et al. The plant PTM viewer, a central resource for exploring plant protein modifications. Plant J. 99, 752–762 (2019).
Guo, T. et al. ERECTA1 acts upstream of the OsMKKK10-OsMKK4-OsMPK6 cascade to control spikelet number by regulating cytokinin metabolism in rice. Plant Cell 32, 2763–2779 (2020).
Ma, H. et al. Pathogen-inducible OsMPKK10.2-OsMPK6 cascade phosphorylates the Raf-like kinase OsEDR1 and inhibits its scaffold function to promote rice disease resistance. Mol. Plant 14, 620–632 (2021).
Yoo, M., Albert, V., Soltis, P. & Soltis, D. Phylogenetic diversification of glycogen synthase kinase 3/SHAGGY-like kinase genes in plants. BMC Plant Biol 6, 3 (2006).
Lyu, J. et al. Control of grain size and weight by the GSK2-LARGE1/OML4 pathway in rice. Plant Cell 32, 1905–1918 (2020).
Qiao, S. et al. The RLA1/SMOS1 transcription factor functions with OsBZR1 to regulate brassinosteroid signaling and rice architecture. Plant Cell 29, 292–309 (2017).
Tong, H. et al. DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice. Plant Cell 24, 2562–2577 (2012).
Xiao, Y. et al. GSK2 stabilizes OFP3 to suppress brassinosteroid responses in rice. Plant J. 102, 1187–1201 (2020).
Yang, C., Shen, W., He, Y., Tian, Z. & Li, J. OVATE family protein 8 positively mediates brassinosteroid signaling through interacting with the GSK3-like kinase in rice. PLoS Genet. 12, e1006118 (2016).
Che, R. et al. Control of grain size and rice yield by GL2-mediated by brassinosteroid responses. Nat. Plants 2, 15195 (2015).
Hu, Z. et al. A novel QTL qTGW3 encodes the GSK3/SHAGGY-like kinase OsGSK5/OsSK41 that interacts with OsARF4 to negatively regulate grain size and weight in rice. Mol. Plant 11, 736–749 (2018).
Ma, M. et al. Control of grain size in rice by TGW3 phosphorylation of OsIAA10 through potentiation of OsIAA10-OsARF4-mediated auxin signaling. Cell Rep. 42, 112187 (2023).
Ding, S., Zhang, B. & Qin, F. Arabidopsis RZFP34/CHYR1, a ubiquitin E3 ligase, regulates stomatal movement and drought tolerance via SnRK2.6-mediated phosphorylation. Plant Cell 27, 3228–3244 (2015).
Schneider, C., Rasband, W. & Eliceiri, K. NIH image to imageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Acknowledgements
We would like to thank Prof. Lei Wang (Institute of Botany) for sharing the plasmid vectors pBI-2YN and pBI-2YC, and Prof. Yuxin Hu (Institute of Botany) for sharing the plasmid vectors pSuper1300-GFP and pSuper1300-Myc. We also thank Dr. Xiuping Xu, Dr. Zhuang Lu, and Dr. Jingquan Li (Institute of Botany) for technical assistance. This work was supported by STI 2030-Major Projects (2023ZD0406902), the Strategic Priority Research Program of Chinese Academy of Sciences (XDA24010101-2), the National Key Research and Development Program of China (2016YFD0100402), and the National Natural Science Foundation of China (91735302, 91435113, and 31471466).
Author information
Authors and Affiliations
Contributions
X.-J.S. designed the study. C.B., G.-J.W., X.-H.F., and Q.G. performed most of the experiments and analyzed the data. W.-Q.W., R.X., S.-J.G., S.-Y.S., M.M., W.-H.L., C.-M.L, and Y.-H.L. conducted some of the experiments. C.B. and X.-J.S. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Jian Zhang, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bai, C., Wang, GJ., Feng, XH. et al. OsMAPK6 phosphorylation and CLG1 ubiquitylation of GW6a non-additively enhance rice grain size through stabilization of the substrate. Nat Commun 15, 4300 (2024). https://doi.org/10.1038/s41467-024-48786-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-48786-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
