
Abstract
The Epidermal Growth Factor Receptor (EGFR) is frequently found to be mutated in non-small cell lung cancer. Oncogenic EGFR has been successfully targeted by tyrosine kinase inhibitors, but acquired drug resistance eventually overcomes the efficacy of these treatments. Attempts to surmount this therapeutic challenge are hindered by a poor understanding of how and why cancer mutations specifically amplify ligand-independent EGFR auto-phosphorylation signals to enhance cell survival and how this amplification is related to ligand-dependent cell proliferation. Here we show that drug-resistant EGFR mutations manipulate the assembly of ligand-free, kinase-active oligomers to promote and stabilize the assembly of oligomer-obligate active dimer sub-units and circumvent the need for ligand binding. We reveal the structure and assembly mechanisms of these ligand-free, kinase-active oligomers, uncovering oncogenic functions for hitherto orphan transmembrane and kinase interfaces, and for the ectodomain tethered conformation of EGFR. Importantly, we find that the active dimer sub-units within ligand-free oligomers are the high affinity binding sites competent to bind physiological ligand concentrations and thus drive tumor growth, revealing a link with tumor proliferation. Our findings provide a framework for future drug discovery directed at tackling oncogenic EGFR mutations by disabling oligomer-assembling interactions.
Similar content being viewed by others
Introduction
Epidermal growth factor receptor (EGFR) is a transmembrane tyrosine kinase receptor at the heart of signals for cell survival, growth and division1. EGFR signals for growth and division are regulated on the cell surface by the binding of cognate growth factor ligands, which promote the assembly of a two-liganded back-to-back ectodomain dimer (B2Bectdimer)2. This dimer underpins across the membrane the formation of an asymmetric kinase dimer (Asymkindimer)3, which is fundamental to catalyze EGFR auto-phosphorylation in C-terminal tyrosine residues1 (Fig. 1a). In the context of ligand-bound oligomers, EGFR auto-phosphorylation triggers EGFR-dependent canonical downstream signaling pathways, like PI3K/Akt and Ras-MAPK, which promote cell growth and proliferation4,5,6.

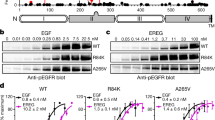
a Left, Cartoon of a ligand-free tethered ectodomain (subdomains numbered) linked to a kinase monomer; the ATP-binding site, T766M mutation, and C-terminal tail are marked. Right, A two-liganded (EGF) extended back-to-back ectodomain dimer (B2Bectdimer) structurally coupled across the plasma membrane80 to an asymmetric kinase dimer (Asymkindimer)3, in which an “activator” kinase (teal) stabilizes a “receiver” kinase (dark green) in the active conformation. b Left, a ligand-free head-to-head ectodomain dimer (H2Hectdimer) sub-unit linked to two kinase monomers36. Middle, a ligand-free back-to-back ectodomain dimer (B2Bectdimer) sub-unit81 coupled to a head-to-head kinase dimer (H2Hkindimer) sub-unit35. Right, a ligand-free stalk-to-stalk ectodomain dimer (St2Stectdimer) sub-unit36 coupled to the Asymkindimer sub-unit36.
Non-small cell lung cancer (NSCLC) has a 5-year relative survival rate of 28%7,8. Important examples of oncogenic drivers of NSCLC are somatic EGFR mutations in exons 18-21, frequently identified (10-60% of lung adenocarcinomas9,10,11) among patients successfully treated with first-generation ATP-competing tyrosine kinase inhibitors (TKIs)12, and in-frame insertions of three or more base pairs in exon 20 (Ex20Ins), unresponsive to TKIs and accounting for 4-10% of all EGFR mutations in NSCLC13. In TKI-responsive patients, resistance nevertheless emerges with high frequency14,15,16,17 through the development of a dominant secondary T766M mutation in exon 20 at the entrance of the ATP-binding site (also known as T790M when the 24 amino acid signal peptide is included) (Fig. 1a), that increases ATP-binding affinity18. Partial sensitivity is maintained with second-generation irreversible TKIs19, like Afatinib20, that form covalent bonds with a cysteine residue in the ATP-binding pocket, but their potency against wild-type (WT)-EGFR induces severe epithelium-based toxicity21. This limitation was surmounted with highly selective, third-generation irreversible TKIs, such as Osimertinib22,23. However, initial efficacy is overcome by the acquisition of mutations in residues that form covalent bonds with irreversible inhibitors, like C773S24,25,26. Mutant-selective allosteric drugs (so-called fourth-generation) like EAI04527 overcome resistance to third-generation TKIs by preventing the kinase domain from adopting its active conformation when combined with Cetuximab to also block ligand-induced EGFR dimerization27,28,29. However, toxicity-related concerns from off-target effects of Cetuximab limit therapeutic potential30,31.
Current approaches target either the ATP-binding pocket (Fig. 1a), the conformation of the kinase monomer and/or ligand-induced dimerization. One yet untested strategy is to interfere with the poorly characterized ligand-independent (or constitutive) kinase-active state of EGFR, which elicits non-canonical, EGFR-dependent signals for cell survival32,33. This state is selectively amplified by NSCLC mutations, typically resulting in a few-fold increase in ligand-independent autophosphorylation34, and could therefore underpin ligand-dependent tumor proliferation. However, the associated mechanisms remain poorly characterized. The best understood ligand-free structures have inactive kinases, namely EGFR monomers and autoinhibited dimers made up of a ligand-free B2Bectdimer structurally coupled to a symmetric head-to-head kinase dimer (B2Bect/H2Hkindimer)35 (Fig. 1b). Exploiting super-resolution Fluorophore Localization Imaging with Photobleaching (FLImP)6,36,37,38,39,40, a molecular ruler that measures inter-receptor separations in specifically-labeled dimers and oligomers on cells (Fig. 2), guided by protein structures and molecular dynamics (MD) simulations, we previously made progress towards understanding further the ligand-independent state of an autoinhibited oligomer EGFR architecture based on repeats of a head-to-head ectodomain dimer linked to non-interacting kinase monomers (H2Hect/2xkinmonomers)36 (Fig. 1b). We also previously detected the FLImP-separation signatures of two full-length ligand-free dimer conformers. One is consistent with the structure of the B2Bect/H2Hkindimer35 (Fig. 1b). In the other, the Asymkindimer is structurally coupled to a ligand-free stalk-to-stalk ectodomain dimer (St2Stect dimer)36 (Fig. 1b). However, in the St2Stect/Asymkindimer, which could account for ligand-independent signals, the Asymkindimer is disfavored by the WT-EGFR kinase41,42. Moreover, when the Asymkindimer is artificially joined to two ligand-free ectodomains, the dimer is disordered in electron micrographs43,44. Intriguingly, the T766M mutation, despite increasing EGFR autophosphorylation in the absence of ligand, counterintuitively destabilizes the catalytic Asymkindimer in favor of the inactive H2Hkindimer36 (Fig. 1b). Given this, we conjectured that the St2Stect/Asymkindimer might be an obligate oligomer sub-unit chaperoned by the B2Bect/H2Hkindimer in heteroconf-oligomers. However, the previous ~5 nm resolution of FLImP prevented us from testing the stated hypothesis because we could not resolve some dimer separations from each other and/or from those that arise from the interaction between dimer sub-units in an oligomer.

a EGFR is a transmembrane protein that forms higher-order associations on the scale of 0-70 nm. EGFR is amenable to fluorescent labeling at several sites, in this case a specific site of the extracellular domain (DIII) (the intracellular domain is not depicted); the distances (d1-dn) provide a detailed structural signature. b When visualized under TIRF microscopy in cells, the size of labeled-EGFR dimers and oligomers is smaller than the diffraction limit of the microscope. Therefore, the fluorescent tags associated to EGFR dimers and oligomers emit light within a diffraction-limited point-spread function (PSF), which is also the microscope image of a single molecule (yellow spots). To resolve the positions of multiple EGFR molecules within a single diffraction limited spot, a video acquisition (FLImP raw data) is taken as the fluorophores photobleach. Single-molecule feature detection and tracking of these videos reveals integrated intensities through time and a subset of these spots which have multistep photobleaching (c) is identified (track selection); such single molecules bleach in single steps, and this can be used to estimate the number of fluorophores emitting light (red) in the spot as a function of time. By combining this information with prior knowledge of PSF shape, we can then fit the selected spots through time with a varying sum of Gaussian PSFs model to determine the positions of the emitting fluorophores (FLImP localization fit) with associated uncertainties (d). Multiple such measurements in the form of empirical posteriors (e) can be pooled (summed) into a FLImP signature of the structure (f, gray histogram). The green vertical line in e is the mean and the yellow shading represents a 70% confidence interval. Finally, a Bayesian decomposition of the separation measurement set (e) is performed to determine the most likely set of unique discrete separations present within the structure (f, colored components). In f the legend and bars above colored component distributions give the median and most-compact 68% confidence interval for each. Legend also gives median proportion of measurements assigned to clutter. (More in Supplementary Note 3).
Here we implement a higher-resolution FLImP version which, combined with large-scale simulations of various membrane-embedded dimer interfaces, allowed us to build an experimentally backed model of all the relevant interactions required to assemble St2Stect/Asymkindimer sub-units within ligand-free EGFR oligomers. We show that WT-EGFR, T766M-EGFR, and Ex20Ins-EGFR share a ligand-free heteroconf-oligomer structure in which scaffolds made of H2Hect/2xkinmonomers and B2Bect/H2Hkindimer sub-units held by a transversal transmembrane interface45 cantilever into position the extracellular portion of the St2Stect/Asymkindimer under the regulation of the ectodomain tethered conformation46. Within these heteroconf-oligomers, St2Stect/Asymkindimer sub-units are positively and negatively regulated via two hitherto functionally-orphan kinase interfaces (PDB IDs:3VJO [https://www.rcsb.org/structure/3VJO]47 and in 5CNO [https://www.rcsb.org/structure/5CNO]48). Stabilization by T766M and Ex20Ins NSCLC mutations of the ancillary kinase interfaces leads to an increase in the number and stability of St2Stect/Asymkindimer sub-units, which accounts for mutation-dependent increases in ligand-free phosphorylation. Excitingly, our finding that St2Stect/Asymkindimer sub-units are epidermal growth factor (EGF) ligand high affinity binding sites, which are the sites previously proposed to drive tumor growth under pM physiological ligand concentrations49, explains how the dysregulation of the ligand-free kinase active state by NSCLC mutations translates into their ability to potentiate EGFR-dependent tumor growth in vivo.
Results
FLImP reports oligomer size and sub-unit conformers
Photobleaching imaging correlation spectroscopy50 analysis of CHO cells expressing ~105 copies/cell of WT-EGFR and EGFR mutations indicates that ~20–30% of EGFRs are incorporated in dimers and ~15–40% in oligomers (Supplementary Fig. 1a), consistent with previous results in fixed36 and live cells51. FLImP measures the lateral pairwise separations on the scale of 0–70 nm between fluorescent probes specifically bound to the extracellular domain of protomers in non-monomer structures on the cell surface, and their relative abundance6. Mimicking the viewpoint of FLImP microscopy, we can assume the orthogonal projection onto the cell surface (xy-plane) of a heteroconf-hexamer assembled by two dimer conformers (Fig. 3a). We created a dataset of synthetic FLImP separation probability distributions simulating measurements of individual separations between fluorophores bound to the heteroconf-hexamer, including spurious fluorophore localizations (clutter), and noise, which we summed (Fig. 3b; gray background).

a Cartoon dimer conformers assembling a heteroconf-hexamer. Separations between bound fluorophores (red circles) are: two 1st-order interfaces (D1, D2), short and long diagonals (O1, O2), and 2nd-order vertical (O3). b FLImP analysis of 100 synthetic pairwise heteroconf-hexamer separations (inset) (Supplementary Note 3). Sum of posteriors of individual separations between fluorophores (gray background) and abundance-weighted probability distributions of individual components of decomposed separation distribution (colored peaks). The area under each peak is weighted according to the estimated proportion of measurements attributed to that peak (“abundance”). Plot legend and bars above colored component distributions give the median and most-compact 68% confidence interval for each. Legend also gives median proportion of measurements assigned to clutter. Stars and dots below show individual separations assigned to peaks (colored) or clutter (black). c As (b) for the homo-trimer formed by the Dimer 2 conformer. d Top, map of mutations and treatments superimposed on the B2Bectdimer81, an Asymkindimer3, and H2Hkindimer54 sub-units. Mutations colored according to the different dimer conformers they inhibit or disrupt. Bottom, EGFR sequence diagram. e Comparisons between decomposed separation probability distributions between datasets. The continuous lines show the marginalized separation posterior for each condition, the sum of the abundance-weighted peaks in (b and c). Fluctuations around each continuous line arise from variations derived from FLImP decompositions for 20 bootstrap-resampled datasets to assess errors due to finite number of measurements. f Wasserstein MDS analysis of FLImP decompositions. This measures the work needed to convert a decomposed separation set into another, thereby estimating similarities and differences between whole FLImP separation decompositions. Similarities or dissimilarities between the 21 separation sets of different conditions (one main FLImP decomposition plus 20 bootstrap-resampled decompositions) are compared; in this case for the heteroconf-hexamer (navy) (b) and homo-trimer (cyan) (c). The plot axes are components C1 and C2. C1 represents the dimension that captures the largest amount of data variance; C2 represents the second-largest amount of variance orthogonal to C1. The ellipse centers (95% confidence range) mark the positions of the main FLImP decompositions. Crosses mark the positions of individual bootstrap-resampled separation sets.
If individual dimeric units of different conformations are present on the cell surface, FLImP will report one separation per type. If, as shown in Fig. 3a, heteroconf-oligomers are assembled by two dimer conformers, the FLImP peaks report 1st, 2nd, 3rd, etc., neighbor separations in the oligomer structure (Fig. 3b; color peaks). With the help of mutations, changes in intensity and position of these peaks can be used to investigate the structural sub-units assembling an oligomer. As an example, we simulated synthetic FLImP separations for the homo-trimer that forms when the dimer 1 conformer is disrupted (Fig. 3c, gray background). FLImP decomposes from these data the remaining 1st and 2nd-order vertical separations from the homo-trimer (Fig. 3c, color peaks), revealing which 1st order separation belongs to the disrupted interface and which higher-order peaks are dependent on heteroconf-interactions. The mutations and treatments that we used to dissect the interactions assembling ligand-free oligomers are mapped-out in Fig. 3d.
FLImP samples a finite population of separations, and this introduces errors. We, therefore, used bootstrap-resampling52 to estimate how this affects the decomposition. Figure 3e illustrates that bootstrap-resampling can capture changes caused by the transition from heteroconf-hexamer to homo-trimer and evaluate significance above finite sampling errors. These synthetic results suggest a resolution of <3 nm, which was experimentally validated in cells with a known membrane protein structure (Supplementary Fig. 1b).
Given enough resolution and with the help of mutations/treatments, separation peaks from 1st order interfaces are typically amenable to be assigned, but assigning higher order peaks can be harder. We interrogate the effect of mutations on unassigned higher order peaks by folding them into a multidimensional scaling (MDS) Wasserstein metric53. As shown in the example (Fig. 3f), we also include the bootstrap-estimated errors associated with finite sampling and calibrate changes to report oligomer growth direction.
The B2Bect/H2Hkin dimer and St2Stect/Asymkin dimer are oligomer sub-units
The color peaks in Fig. 4a show the most likely positions and intensities of pairwise separations between CF640R fluorophores specifically conjugated to anti-EGFR Affibodies bound to DIII of cell surface WT-EGFR ectodomains. All FLImP separation sets have hereafter the median positions of the WT-EGFR separations superimposed (dashed lines) to facilitate comparisons. FLImP measurements require immobilizing cell surface receptors by chemical fixation via a method demonstrated not to introduce detectable artefacts6. Nevertheless, predictions arising from the results of chemically fixed cells were validated in live cells using single particle tracking (Supplementary Fig. 1c), and by controls described below.

a, c–f, h FLImP analysis of 100 separation probability distributions between Affibody-CF640R pairs in the conditions indicated: Sum of posteriors of individual separations between fluorophores (gray background) and abundance-weighted probability distributions of individual components of decomposed separation distribution (colored peaks). Plot legend and bars above colored component distributions give the median and most-compact 68% confidence interval for each. Legend also gives median proportion of measurements assigned to clutter. Stars and dots below show individual separations assigned to peaks (colored) or clutter (black). The median peak positions marked by dashed lines are hereafter also superimposed on all the FLImP separation diagrams to facilitate comparisons with WT-EGFR. FLImP separations for all mutants and treatments are summarized in Supplementary Table 3. b Left, cartoon of a B2Bect/H2Hkindimer sub-unit labeled with two fluorescent anti-EGFR Affibody bound to the two DIII of the ectodomains. The ED/RK mutation inhibits the B2Bect/H2Hkindimer sub-unit. Right, a labeled St2Stect/Asymkindimer sub-unit. Erlotinib binds the ATP pocket of the kinase stabilizing the St2Stect/Asymkindimer sub-unit. g, i Comparisons between decomposed separation probability distributions between FLImP datasets. The continuous lines show the marginalized separation posterior, i.e. the sum of the abundance-weighted peaks, for each condition in the inset. Fluctuations around each continuous line arise from variations derived from FLImP decompositions for 20 bootstrap-resampled datasets to assess errors due to the finite number of measurements. Median peak positions for WT-EGFR marked by dashed lines, as in a. j Wasserstein MDS analysis of FLImP decompositions for the conditions in the inset. Similarities or dissimilarities between the 21 separation sets of different conditions (one main FLImP decomposition plus 20 bootstrap-resampled decompositions) are compared. The plot axes are components C1 and C2. C1 represents the dimension that captures the largest amount of variance in the data, while C2 represents the second-largest amount of variance that is orthogonal to C1. The ellipse centers (95% confidence range) mark the positions of the main FLImP decompositions. The crosses mark the positions of individual bootstrap-resampled separation sets.
Interpreting a FLImP separation set requires assigning 1st order peaks to the structural sub-units assembling the underlying dimers/oligomers. In previous work36, we linked the 10.8–13.5 nm peak (Fig. 4a, red dashed line) to the extracellular part of the B2Bect/H2Hkindimer, which could also exist as an oligomer sub-unit (Fig. 4b). The link between the 10.8–13.5 nm peak and the B2Bect/H2Hkindimer, either alone and/or as a protomer, is supported here by the observation that FLImP no longer indicates separation density between 8.1–14.8 nm when a previously proposed double E981R/D982K (ED/RK) mutation in the C-terminus54 that destabilizes electrostatic interactions at the heart of the intracellular portion of the B2Bect/H2Hkindimer is introduced (Fig. 4c; red dashed line). By contrast, separation density at 9.2–13.2 nm increases when the H2Hkindimer is stabilized by the T766M mutation36 (Fig. 4d; red dashed line), further supporting the assignment of the 10.8-13.5 nm peak in the WT-EGFR separation set to the B2Bect/H2Hkindimer conformer.
We previously linked a <9 nm separation to the extracellular portion of the St2Stect/Asymkindimer36, which could also exist as an oligomer sub-unit (Fig. 4b). Our enhanced FLImP method decomposes two <9 nm components in the WT-EGFR separation set (Fig. 4a; cyan and black dashed lines). A body of previous EGF-binding experiments suggested that EGFR is displayed on the cell surface in two forms, a minority (2–5%) of high-affinity EGFRs (KD = 10-100 pM) and a majority (95-98%) of low-affinity EGFRs (KD = 2–5 nM)55. Biophysical experiments suggested that these two affinity forms arise from two different dimer structures56. Given this, we speculated that if the two components of <9 nm arise from two different dimer conformers, we might be able to assign at least one according to ligand affinity. We pre-treated WT-EGFR-expressing cells with the conformation-selective monoclonal antibodies mAb-2E957 or mAb-10858, which select for high and low-affinity EGF binding, respectively. As the selectivity of these mAbs is bona fide against EGF binding, mAb-treated cells were next probed with an EGF-CF640R derivative. Here cells were fixed after mAb treatment but before probing with EGF-CF640R to avoid ligand-induced conformational changes. Results show that EGF-CF640R binds well to fixed cells at similar sites to Affibody-CF640R (Supplementary Fig. 1d, e).
High-affinity EGFRs have been proposed to be mostly responsible for ligand-dependent EGFR’s signaling57. Confocal results using mAb-2E9 indicate that EGF binds cell surface St2Stect/Asymkindimer conformers with high affinity (Supplementary Fig. 1f). The FLImP separation set from mAb-2E9-treated cells suggests two components of <9 nm (4.9–6.8 nm and 7.2–8.9 nm) (Fig. 4e; cyan and black dashed lines), which are consistent with those in the WT-EGFR separation set. In contrast, no separation density at 6.8–9.9 nm is apparent in the set from cells treated with mAb-108, which blocks high-affinity binding (Fig. 4f; black dashed line). To assess the robustness of this result, because incompletely resolved components cannot be perfectly separated, we compared the evidence for separations in bootstrap-resampled datasets after pooling the individual components (marginalized probability). This analysis indicates that the absence of separation density in the vicinity of ~8 nm associated to the mAb-108 treatment is robust to finite sampling errors (Fig. 4g). We therefore assigned the 7.0–8.6 nm peak in the WT-EGFR separation set to high-affinity EGF-binding St2Stect/Asymkindimer sub-units. Further validation of this assignment is provided below.
Despite the T766M mutation destabilizing the Asymkindimer36, the separation density at ~8 nm is similar between WT-EGFR and T766M-EGFR (Fig. 4a, d). We hypothesized that the T766M mutation underpins the St2Stect/Asymkindimer via interactions with the inactive B2Bect/H2Hkindimer, which is stabilized by the T766M mutation36. To test this, WT-EGFR expressing cells were treated with Erlotinib59, a TKI that binds to EGFR’s kinase ATP-binding pocket, stabilizing the St2Stect/Asymkindimer60. With our previous poorer resolution, we found that Erlotinib treatment enhances a broad peak encompassing separations 1.6-10.2 nm36. Our higher resolution method decomposes three components under this peak (Fig. 4h). Interestingly, Erlotinib treatment recapitulates effects induced by the T766M mutation, most notably on separations of <20 nm associated with Erlotinib-treated WT-EGFR and T766M-EGFR (Figs. 4h, d). Quantified in Fig. 4i, both induce a decrease in the 5.8–7.1 nm component, the assignment and function of which is discussed below, and an increase at ~9–12 nm. These results argue that the stabilizing the St2Stect/Asymkindimer directly by Erlotinib binding shares characteristics of stabilizing the B2Bect/H2Hkindimer via the T766M mutation, suggesting that the B2Bect/H2Hkindimer stabilizes the St2Stect/Asymkindimer within heteroconf-oligomers.
Further evidence that the B2Bect/H2Hkindimer and St2Stect/Asymkindimer are structural sub-units in ligand-free heteroconf-oligomers is shown in Fig. 4j. This includes the increase (decrease) in oligomer size induced by the T766M (ED/RK) mutations and the different oligomer sizes associated with mAb-2E9 and mAb-108. Notably, Erlotinib, as previously found36, does not significantly increase oligomer size. These results assign the role of underpinning oligomer growth to the H2Hkindimer.
H2Hect dimer/2xkin monomers are sub-units in heteroconf-oligomers
In a previous study36, we proposed a third ligand-free dimer conformer based on a lattice contact in an X-ray structure of the tethered ectodomain monomer (PDB ID:4KRP [https://www.rcsb.org/structure/4KRP]61), in which the monomers are held by ectodomain interactions (H2Hectdimer) (Fig. 5a). This model was supported by the FLImP results associated with ΔC-EGFR, a mutant in which the intracellular domains are deleted, here reanalyzed with the higher resolution decomposition (Fig. 5b). Results show peaks at almost a fixed interval consistent with the previously proposed homo-oligomers of repeating extracellular H2Hectdimer units36. The presence of H2Hectdimer/2xkinmonomers is further supported by the finding that the 1st-order separation of the truncated H2Hectdimer of ΔC-EGFR shares position with the shortest component in the separation set for WT-EGFR (Fig. 5b; cyan dashed line). Taken together, previous and current evidence confirms the presence of H2Hectdimer/2xkinmonomers as a third ligand-free dimer conformer.

a Left, cartoon of full-length H2Hect/2xkinmonomers sub-unit with two fluorescent anti-EGFR Affibodies (blue with red spot) bound to ectodomain’s DIIIs. The H2Hectdimer is linked to two non-interacting kinase monomers36. ΔC-EGFR deletion mutant length is marked. DIV-binding tether-disruptive mutations (e.g. H566F) interfere with the H2Hectdimer conformation. Right, a labeled St2Stect/Asymkindimer sub-unit. Lapatinib binds the kinase ATP-binding pocket disrupting the Asymkindimer. b, d, f FLImP analysis of 100 separation probability distributions between Affibody-CF640R pairs in the conditions indicated: Sum of posteriors of individual separations between fluorophores (gray background) and abundance-weighted probability distributions of individual components of decomposed separation distribution (colored peaks). Plot legend and bars above colored component distributions give the median and most-compact 68% confidence interval for each. Legend also gives median proportion of measurements assigned to clutter. Stars and dots below show individual separations assigned to peaks (colored) or clutter (black). WT-EGFR median peak positions marked by dashed lines. c Wasserstein MDS analysis of FLImP decompositions for conditions in the inset. Similarities or dissimilarities between the 21 separation sets of different conditions (one main FLImP decomposition plus 20 bootstrap-resampled decompositions) are compared. The axes are components C1 and C2. C1 represents the dimension that captures the largest amount of data variance, while C2 represents the second-largest amount of variance that is orthogonal to C1. The ellipse centers (95% confidence range) mark the positions of the main FLImP decompositions. The crosses mark the positions of individual bootstrap-resampled separation sets. e Comparisons between decomposed separation probability distributions between FLImP datasets. The continuous lines show the marginalized separation posterior, i.e. the sum of the abundance-weighted peaks, for each condition in the inset. Fluctuations around each continuous line arise from variations derived from FLImP decompositions for 20 bootstrap-resampled datasets to assess errors due to finite number of measurements. Note that the 8 nm separation corresponding to the St2Stect/Asymkindimer sub-unit is not significantly decreased by the H566F mutation (orange), consistent with the tether-disrupting mutations not inhibiting phosphorylation55 (Supplementary Fig. 3e). Dashed lines mark WT-EGFR median peak positions. Colored arrows show shift from WT.
Higher resolution unmasked differences beyond the 1st order separation between the ΔC-EGFR and WT-EGFR sets (Fig. 5c). Given this, we speculated that H2Hectdimer/2xkinmonomers conformers might participate in heteroconf-oligomerization. To investigate this, we introduced mutations that disrupt the H2Hectdimer. Our MD simulations suggest that inhibition of the tether via two well-understood DIV mutations in EGFR’s ectodomain, H566F and G564P55, disrupts the H2Hectdimer but not the B2Bectdimer (Supplementary Fig. 2 and Supplementary Tables 1 and 2). Consistent with previous data55, our MD simulations also suggest that H566F, but not G564P, rearranges the DI of EGFR’s ectodomain in a way that makes the binding site less accessible (Supplementary Fig. 2). Using FLImP we found that H566F decreases oligomer size (Fig. 5c) and induces a shift in the shortest separation component assigned to H2Hectdimer/2xkinmonomers conformers from a median position of 6.5 nm to 4 nm, which implicates the ectodomain tethered conformation in the formation of H2Hectdimer/2xkinmonomers (Figs. 5d, e; blue and orange). Analogous results for G564P are in Supplementary Fig. 3a–d.
Because MD simulations suggest that the WT-EGFR H2Hectdimer can explore a separation range of 3–7 nm between the center of mass of the two DIIIs (Supplementary Fig. 2c, Supplementary Note 1), the shift in the separation component annotated to H2Hectdimer/2xkinmonomers conformers from a median position of 6.5 nm to 5.2 nm introduced by the ED/RK mutation (Fig. 5e; blue and green) suggests that inhibiting the B2Bect/H2Hkindimer changes H2Hectdimer/2xkinmonomers conformation. The ED/RK mutations also induce a comparable oligomer size reduction to that of the H566F mutation (Fig. 5c). Together these results suggest that B2Bect/H2Hkindimer sub-units interact with H2Hectdimer/2xkinmonomers, and hence that the latter is also a sub-unit in ligand-free heteroconf-oligomers.
From this, it follows that H2Hectdimer/2xkinmonomers might interact with the St2Stect/Asymkindimer. To test this, WT-EGFR expressing cells were treated with Lapatinib, a TKI that binds to EGFR’s kinase ATP-binding site breaking the Asymkindimer62. As expected, Lapatinib induced a significant reduction around the 8 nm position assigned to the St2Stect/Asymkindimer conformer (Fig. 5e; blue and red, and 5f)60. This is also consistent with our finding that St2Stect/Asymkindimer sub-units display high-affinity because Lapatinib additionally decreases EGF-binding affinity60. The reduction in the ~8 nm component is accompanied by a shift in the component assigned to the H2Hectdimer/2xkinmonomers conformer, from a median position of 6.5 nm to 4.2 nm (Fig. 5e, f; cyan dashed line), arguing that inhibiting the St2Stect/Asymkindimer changes the conformation of H2Hectdimer/2xkinmonomers, and hence that H2Hectdimer/2xkinmonomers and St2Stect/Asymkindimer sub-units interact. Interestingly, Lapatinib does not decrease oligomer size (Fig. 5c). This will be discussed later with more data (Fig. 9).
Ligand-free conformers interact via transmembrane contacts
The conformation of the three ligand-free dimer sub-units suggest heteroconf-oligomer assembly might be mediated by transmembrane interactions. Therefore, we next considered the Lzip transmembrane dimer named after its leucine zipper-like interactions (Fig. 6a)45. In principle, the Lzip interface could mediate interactions between transmembrane monomers and dimers (Fig. 6b). To investigate this possibility, we mutated all three Lzip dimer transmembrane helix amino acids (V635, L638, L642) to either serine or alanine, named Lzip3S and Lzip3A, respectively. Based on previous literature, Lzip3S mutations would be expected to strongly inhibit the Lzip interaction, unlike the more conservative Lzip3A63.

a Previously proposed transmembrane dimers45. Lzip has leucine zipper-like interactions (V635xxL638xxxL642) and could establish contacts with Nter+/Nter and Cter+/Cter to assemble oligomers. b Top, a speculative tetramer (yellow) from two monomers interacting through Lzip contacts with one Cter interface. A Cter dimer model (magenta) is placed on top of the tetramer showing that the residues in the Cter interface are distinct from Lzip. Bottom, orthogonal projection on xy-plane. (Equivalent for Lzip contacts with one Nter interface in Supplementary Fig. 3f). c, f, g FLImP analysis of 100 separation probability distributions between Affibody-CF640R pairs in the conditions indicated: Sum of posteriors of individual separations between fluorophores (gray background) and abundance-weighted probability distributions of individual components of decomposed separation distribution (colored peaks). Plot legend and bars above colored component distributions give the median and most-compact 68% confidence interval for each. Legend also gives median proportion of measurements assigned to clutter. Stars and dots below show individual separations assigned to peaks (colored) or clutter (black). The median peak positions marked by dashed lines are those of WT-EGFR. d Comparisons between decomposed separation probability distributions between FLImP datasets. The continuous lines show the marginalized separation posterior, i.e. the sum of the abundance-weighted peaks, for each condition in the inset. The fluctuations around each continuous line arise from variations derived from FLImP decompositions for 20 bootstrap-resampled datasets to assess errors due to the finite number of measurements. Dashed lines mark WT-EGFR median peak positions. Colored arrows show shift from WT. e Wasserstein MDS analysis of FLImP decompositions for the conditions in the inset. Similarities or dissimilarities between the 21 separation sets of different conditions (one main FLImP decomposition plus 20 bootstrap-resampled decompositions) are compared. Plot axes are components C1 and C2. C1 represents the dimension that captures the largest amount of variance in the data, while C2 represents the second-largest amount of variance that is orthogonal to C1. The ellipse centers (95% confidence range) mark the positions of the main FLImP decompositions. The crosses mark the positions of individual bootstrap-resampled separation sets.
We found that short separations become poorly resolved in the Lzip3S-EGFR set (Fig. 6c). Notable effects include increased separation density at 2.2-4.6 nm (Fig. 6c, d), suggesting the conformation of H2Hectdimer/2xkinmonomer sub-units has changed, alongside an oligomer size reduction (Fig. 6e). The Lzip3A mutations did not show these effects (Supplementary Fig. 3g–i). Together, these results implicate Lzip contacts in heteroconf-oligomer assembly.
To evaluate the consistency of the FLImP results, we reasoned that simultaneously inhibiting the H2Hkindimer and Asymkindimer, and thus all intra-dimer intracellular interactions, together with Lzip contacts should recapitulate the ΔC-EGFR results. This hypothesis was evaluated in two stages. First, by combining the ED/RK mutations with L680N, a kinase N-lobe mutation that inhibits the kinase domain from acting as receiver54, and thus the St2Stect/Asymkindimer. Then the Lzip3S mutations were added. Reassuringly, the ED/RK + L680N mutations induce a shift in the peak assigned to H2Hectdimer/2xkinmonomers sub-units comparable to the Lzip3S mutations alone, confirming that the conformation of the H2Hectdimer/2xkinmonomers sub-units depends on heteroconf-oligomer interactions (Fig. 6d, f). Also reassuringly, combining ED/RK + L680N + Lzip3S mutations recapitulated the results for ΔC-EGFR (Fig. 6e, g), e.g., pseudo-periodic separations, oligomer size increase, and the loss of the 2.8–4.3 nm peak, suggesting that Lzip contacts inhibit the formation of the homo-oligomers of repeating extracellular H2Hectdimer interfaces.
Extracellular structure of ligand-free heteroconf-oligomers
Based on the above data, we constructed a model of the orthogonal projection on the cell surface of the heteroconf-oligomer based on the known shape and dimensions of the three ligand-free dimer sub-units (Fig. 1b, Supplementary Fig. 4). H2Hectdimer/2xkinmonomers and B2Bect/H2Hkindimer could form a tetramer in which, remarkably, the Lzip interface aligns one ectodomain of the H2Hectdimer/2xkinmonomers so it can link with another tetramer and form the extracellular portion of the St2Stect/Asymkindimer (Fig. 7a).

a Top left, Tetramer assembled by an H2Hect/2xkinmonomers sub-unit and a B2Bect/H2Hkindimer sub-unit. A transmembrane monomer of an H2Hect/2xkinmonomers interacts with the transmembrane dimer of the B2Bect/H2Hkindimer via an Lzip interface. The Lzip interface is related to the transmembrane dimers in the B2Bect/H2Hkindimer sub-unit and St2Stect/Asymkindimer sub-unit by a rotation of the helix along the long axis45. Bottom left, two tetramers form a St2Stect/Asymkindimer sub-unit. Note that the latter could in principle form also from one H2Hect/2xkinmonomers sub-unit forming an Lzip interface with a tetramer made of one H2Hect/2xkinmonomers sub-unit and a B2Bect/H2Hkindimer sub-unit. (Attempts to join a H2Hectdimer/2xkinmonomers sub-unit and a St2Stect/Asymkindimer sub-unit via Lzip contacts led to steric clashes. More details in Supplementary Fig. 4). Right, Annotated interfaces in the model. b–h Seven distinct optimally grouped triangles from 2D FLImP separations determined between EGF-CF640R probes bound to cell surface T766M-EGFR (Supplementary Fig. 5). The side lengths of each triangle are annotated in each inset in the colors used in 1D FLImP decompositions when found in the separation sets of either T766M-EGFR or WT-EGFR (Fig. 4a, d), and colored and underlined if found in both. Separations >70 nm are outside the range of 1D FLImP. The abundance of each triangle (T1-7) is stated bottom right. i Ligand-free heteroconf-oligomer model extended as described in a. The approximate positions where EGF-CF640R would bind DIII of the ectodomains are marked (red circles). Four versions of (b), the experimentally-optimized triangle 1 within errors (T1a-T1d), are superimposed. Inset, table of the side lengths of the superimposed triangles. j Top, ligand-free heteroconf-oligomer model with the triangle groups T2-T4 (c–e) superimposed. Bottom, heteroconf-oligomer model with the largest triangles T5-T7 (f–h) superimposed. Inset, table of the side lengths of the superimposed triangles.
To test this model and its relevance to the dysregulated ligand-free state, we implemented a 2D version of FLImP that reports triangular arrangements between probes bound to EGFR structures (Supplementary Fig. 5). The 3-fold higher probe concentration required for 2D FLImP was better suited to the less sticky EGF-CF640R derivative, as Affibody-CF640R at this higher concentration began to show signs of non-specific binding on the glass supporting the cells64.
As an example, we probed cells expressing T766M-EGFR with EGF-CF640R after chemical fixation to avoid ligand-induced conformational changes, as discussed above. The resulting 2D FLImP triangle dataset was optimally grouped into seven distinct triangles (Fig. 7b–h). Reassuringly, the 1D separations in the triangles are found as components decomposed by 1D FLImP for T766M-EGFR and WT-EGFR (Fig. 4d, a), arguing that T766M-EGFR and WT-EGFR share heteroconf-oligomer structure.
To evaluate whether the 2D FLImP data supports the proposed model, the model was expanded to the size required by the triangles and the positions expected for EGF-CF640R bound to EGFR’s ectodomain DIII marked. We found that the smallest triangle (T1) accounts, within errors, for four triangular probe motifs in the model (Fig. 7i).Triangles T2-T7 each account for one motif (Fig. 7j). To further validate the applicability of the heteroconf-oligomer structure to T766M-EGFR, we verified that combining the T766M mutation with the tether-disrupting H566F mutation or the Lzip3S mutations also disrupts the heteroconf-oligomers (Supplementary Fig. 6). This, together with the excellent results of superimposing the triangles from 2D FLImP data collected from CHO cells expressing T766M-EGFR indicate that the model is an accurate representation of the extracellular portion of the ligand-free heteroconf-oligomers. Interestingly, this model predicts that the smallest oligomers that could bear one St2Stect/Asymkindimer sub-unit are a hexamer assembled from two H2Hectdimer/2xkinmonomers sub-units and one B2Bect/H2Hkindimer and an octamer assembled from two of each (Fig. 7a). Consistent with this, mutations that preserve phosphorylation are at least hexamer in size (Supplementary Table 3).
A ligand-independent mechanism of Ex20ins-induced activation
The model predicts that inhibiting the B2Bect/H2Hkindimer conformer would result in the tetramer shown in Fig. 8a. Inhibiting the Lzip interface should have the same effect, but separations found when B2Bect/H2Hkindimer sub-units are inhibited by the ED/RK mutations are inconsistent with those found when Lzip contacts are inhibited via Lzip3S mutations (Fig. 8b). This hinted at the possibility that we had not yet accounted for all heteroconf-oligomer assembling interactions.

a Tetramer that would form if B2Bect/H2Hkindimer sub-units or Lzip contacts were inhibited (interfaces annotated). b, f, k Comparisons between decomposed separation probability distributions between FLImP datasets. Continuous lines show the marginalized separation posterior sum of the abundance-weighted peaks, for each condition in the inset. Fluctuations around each continuous line arise from variations from FLImP decompositions for 20 bootstrap-resampled datasets to assess errors from finite measurements number. c Structure and cartoon of the Bb2Bbkindimer formed mainly through N-to-C lobe interactions (PDB ID 3VJO [https://www.rcsb.org/structure/3VJO])47. A kinase monomer can dock in the Bb2Bbkindimer to form an Asymkindimer, as shown, with the position of the A-loop marked. The K946E mutation breaks the Bb2Bbkininterface interface (Supplementary Fig. 7a, Supplementary Note 1). d Western blot showing phosphorylation in the absence of ligand in transfected CHO cells. (Blot quantification in Supplementary Fig. 7b, representative of n = 3 blots). e, h, j FLImP analysis of 100 separation probability distributions between Affibody-CF640R pairs. The sum of posteriors of individual separations (gray background) and abundance-weighted probability distributions of individual components of decomposed separation distribution (colored peaks). Plot legend and bars above colored component distributions give the median and most-compact 68% confidence interval for each. Stars and dots below show individual separations assigned to peaks (colored) or clutter (black). Legend also gives median proportion of measurements assigned to clutter. Dashed lines mark WT-EGFR median peak positions (also in (k)). g Wasserstein MDS analysis of FLImP decompositions. Similarities or dissimilarities between the 21 separation sets of different conditions (one main FLImP decomposition plus 20 bootstrap-resampled decompositions) are compared. The plot axes are components C1 and C2. C1 represents the dimension that captures the largest amount of data variance, while C2 represents the second-largest amount of variance orthogonal to C1. The ellipse centers (95% confidence range) mark the positions of the main FLImP decompositions. The crosses mark those of individual bootstrap-resampled separation sets. i The Bb2Bbkindimer sub-unit47 (side and top-view). Inserted residues of the Ex20Ins mutation on the αC/β4 loop shown (yellow circles). Source data provided as a ‘Source data’ file.
We considered a functionally orphan symmetric backbone-to-backbone kinase interface (Bb2Bbkininterface), revealed by X-ray crystallography47 (Fig. 8c). Because a kinase monomer could dock into a Bb2Bbkindimer to form an Asymkindimer, we speculated that the Bb2Bbkininterface might be involved in strengthening the St2Stect/Asymkindimer sub-units. Giving credence to this possibility, two non-naturally occurring charge-reversal R938E and K946E mutations, which compromise the Bb2Bbkindimer, as reported by MD simulations (Supplementary Fig. 7a), decrease receptor phosphorylation (Fig. 8d).
Remarkably, we found that the separation set associated with K946E-EGFR is almost indistinguishable from that of ED/RK + L680N-EGFR (Fig. 8e–g). This reveals that disrupting the Bb2Bbkininterface via the K946E mutation recapitulates the effect of jointly inhibiting St2Stect/Asymkindimer and B2Bect/H2Hkindimer sub-units via the combined ED/RK + L680N mutations. This is consistent with St2Stect/Asymkindimer and B2Bect/H2Hkindimer conformers being obligate heteroconf-oligomers. Adding the K946E mutation to T766M-EGFR also reduces oligomer size and decreases receptor phosphorylation (Fig. 8d, g, h, Supplementary Fig. 7c). Together, results suggest that the Bb2Bbkininterface reinforces the St2Stect/Asymkindimer sub-units, and thereby their interaction with the B2Bect/H2Hkindimer in the heteroconf-oligomers.
If the above interpretation is correct, one would expect that stabilizing the Bb2Bbkininterface should mirror the effects of stabilizing the St2Stect/Asymkindimer conformer. We conjectured that Ex20Ins might increase the number of contacts between the kinase domains of the Bb2Bbkininterface, stabilizing that interface (Fig. 8i). This was supported by MD simulations of the WT and D770-N771insNPG (insNPG), a mutant chosen because structural data is available65 (Supplementary Fig. 8a, b). In support of the notion that the Bb2Bbkininterface underpins the stability of St2Stect/Asymkindimer conformers, the separation set for insNPG-EGFR recapitulates the effects of the Erlotinib treatment in WT-EGFR (Fig. 8j, k, Supplementary Fig. 8c), revealing that the mechanism by which insNPG-EGFR dysregulates ligand-independent phosphorylation is by strengthening the Bb2Bbkininterface in heteroconf-oligomers.
A ligand-independent mechanism of T766M-induced activation
After incorporating the Bb2Bbkininterface (Fig. 9a), another prediction is that disrupting Lzip contacts should have an analogous effect on phosphorylation to inhibiting the B2Bect/H2Hkindimer via the ED/RK mutations, but only the latter increases phosphorylation (Fig. 8d). We speculated this might be explained if the B2Bect/H2Hkindimer sequestered kinase monomers from stabilizing the Asymkindimer via the Bb2Bbkininterface (Fig. 9b). In the crystal lattice of the activator-impaired V924R H2Hkindimer48 we noticed a side-to-side interface (S2Skininterface) that could play such a role (Fig. 9c).

a Tetramer that would form after inhibiting B2Bect/H2Hkindimer sub-units or Lzip contacts with two Bb2Bbkininterfaces buttressing an Asymkindimer sub-unit. Ectodomains and kinases belonging to the same receptor in H2Hectdimer/2xkinmonomers sub-units are colored accordingly. b Ligand-free heteroconf-oligomer model incorporating four S2Skininterfaces. c Left, side-view of the S2Skindimer sub-unit maintained mainly through the β2-sheet and αD-helix of the one monomer and the αE- and αΙ-helices of another (PDB ID: 5CNO [https://www.rcsb.org/structure/5CNO])48. The position of the A-loop in the inactive configuration is marked. Right, cartoon representation of the top-view of an H2Hkindimer sub-unit flanked by two monomers via S2Skininterfaces. d, g, h FLImP analysis of 100 separation probability distributions between Affibody-CF640R pairs in the conditions indicated: Sum of posteriors of individual separations between fluorophores (gray background) and abundance-weighted probability distributions of individual components of decomposed separation distribution (colored peaks). Plot legend and bars above colored component distributions give the median and most-compact 68% confidence interval for each. Legend also gives median proportion of measurements assigned to clutter. The median peak positions marked by dashed lines are those of WT-EGFR. e Comparisons between decomposed separation probability distributions between datasets. The continuous lines show the marginalized separation posterior, i.e. the sum of the abundance-weighted peaks, for each condition in the inset. The fluctuations around each continuous line arise from variations derived from FLImP decompositions for 20 bootstrap-resampled datasets to assess errors due to finite number of measurements. Dashed lines marked WT-EGFR median peak positions (as in d). Colored arrows show shift from WT-EGFR. f Wasserstein MDS analysis of FLImP decompositions for the conditions in the inset. Similarities or dissimilarities between the 21 separation sets of different conditions (one main FLImP decomposition plus 20 bootstrap-resampled decompositions) are compared. The plot axes are components C1 and C2. C1 represents the dimension that captures the largest amount of variance in the data, while C2 represents the second-largest amount of variance that is orthogonal to C1. The ellipse centers (95% confidence range) mark the positions of the main FLImP decompositions. The crosses mark the positions of individual bootstrap-resampled separation sets.
We previously reported that the non-naturally occurring I942E mutation inhibits ligand-bound oligomerisation36. Here we show that I942E decreases ligand-free EGFR phosphorylation (Fig. 8d). The I942 residue lies at the heart of the S2Skininterface, and modelling of the WT and I942E mutant suggested that the latter stabilizes the S2Skininterface (Supplementary Fig. 9a, Supplementary Note 1). The separation set associated with I942E-EGFR is consistent with the results obtained for WT-EGFR-expressing cells treated with Lapatinib, which inhibits the Asymkindimer (Fig. 9d, e). These results argue that S2Skininterface stabilization via the I942E mutation also inhibits the St2Stect/Asymkindimer, indicating that the S2Skininterface plays an autoinhibitory role. Combining the T766M and I942E mutations support this notion (Supplementary Fig. 9b, c).
If the B2Bect/H2Hkindimer sequestered kinase monomers via the S2Skininterface preventing these from reinforcing St2Stect/Asymkindimer sub-units via the Bb2Bbkininterface, then inhibiting the Bb2Bbkininterface should make more kinase monomers available to stabilize the B2Bect/H2Hkindimer via the S2Skininterface, thereby growing larger oligomers as found when B2Bect/H2Hkindimer sub-units are stabilized by the T766M mutation (Fig. 4j). To test this possibility, we need to inhibit the Bb2Bbkininterface whilst preserving St2Stect/Asymkindimer and B2Bect/H2Hkindimer sub-units so they can compete for kinase monomers. However, when the Bb2Bbkininterface-inhibitory K946E mutation is introduced in WT-EGFR, the mutation also disassembles the St2Stect/Asymkindimer and B2Bect/H2Hkindimer conformers (Fig. 8f). In contrast, partially disassembled Lzip3S-EGFR oligomers preserve their phosphorylation (Fig. 8d) and some density around 12 nm (Fig. 8b), suggesting that Lzip3S-EGFR oligomers retain some St2Stect/Asymkindimer and B2Bect/H2Hkindimer sub-units. Thus, we reasoned that adding K946E to Lzip3S mutations should stabilize the B2Bect/H2Hkindimer and increase oligomer size. The results are consistent with this notion (Fig. 9f). We also found that the separation sets of Lzip3S + K946E and Lzip3S + L680N are similar below ~35 nm (Fig. 9g, h), suggesting that inhibiting the St2Stect/Asymkindimer and the Bb2Bbkininterface are almost equivalent, and explaining why disrupting the St2Stect/Asymkindimer, either directly via Lapatinib or indirectly via the I942E mutation, does not decrease oligomer size, as both stabilize the S2Skininterface and thereby the H2Hkindimer counterbalancing the effect of disrupting St2Stect/Asymkindimer sub-units (Fig. 9f).
Results, therefore, show that destabilizing the St2Stect/Asymkindimer, either directly by the L680N mutation or indirectly by inhibiting the Bb2Bbkininterface, increases oligomer size. In WT-EGFR this depends on the S2Skininterface at the expense of phosphorylation. The T766M mutation directly stabilizes the B2Bect/H2Hkindimer, thus promoting the formation of larger and more stable36 oligomers that can bear more St2Stect/Asymkindimer sub-units, thus accounting for the increase in T766M-induced ligand-independent phosphorylation (Supplementary Fig. 7c).
Tumor growth depends on heteroconf-oligomerisation
To test the relevance of the proposed ligand-free oligomer structure and assembling mechanisms in vivo, we carried out cellular transformation assays using the IL3-dependent murine lymphoid Ba/F3 cell system with the aim of using these cells to establish tumor xenografts. Ba/F3 cells fail to survive and multiply in the absence of IL366,67, but this phenotype can be rescued by the ectopic expression of a constitutively active receptor tyrosine kinase, like for example T766M-EGFR, which allows survival signaling in the absence of IL366.
We generated Ba/F3 cell lines stably expressing WT-EGFR (Ba/F3 + WT), T766M-EGFR (Ba/F3 + T766M), and EGFR mutants at near equal levels using the PiggyBac system and tested the ability of the transformed cells to grow in the absence of IL3 (Supplementary Fig. 10a). To minimize mice number, from the mutations that disrupt heteroconf-oligomer structure but not phosphorylation we focused on the T766M + H566F and omitted the apparently equivalent T766M+Lzip3S (Supplementary Fig. 6). From the mutations that disrupt oligomer structure and phosphorylation we chose T766M + K946E over T766M + I942E because unlike the I942E mutation, K946E does not interfere with the ligand-bound state (Supplementary Fig. 10b).
Next, we compared the ability of these Ba/F3 cell lines to establish and grow tumors. Notably, all these cell lines also stably expressed GFP, exploited to image growing tumors. We found a lag-time prior to the onset of palpable tumors of around 20 days in all cohorts, after which the tumors formed in animals that had received Ba/F3 + T766M and Ba/F3 + T766M + H566F cells and, with an additional delay, in the Ba/F3 + T766M + K946E cohort (Fig. 10a). Importantly, mirroring the growth pattern observed in vitro, Ba/F3 + T766M tumors grew best throughout, followed by the double mutant tumors (Ba/F3 + T766M + H566F > Ba/F3 + T766M + K946E), while WT-EGFR tumors did not establish or grow. The lack of tumor cell survival in the Ba/F3 + WT cohort was already strongly suggested by day 21 when we observed no GFP reporter signals at the injection site (Supplementary Fig. 10c–e). The experimental endpoint was on day 41 when we imaged all animals by IVIS (Fig. 10b, c) to determine tumor fluorescence as an independent measure of tumor growth between cohorts. We found significant differences in fluorescence signals between groups, in line with caliper measurements (Fig. 10a), except for Ba/F3 + T766M vs. Ba/F3 + T766M + H566F tumors. The latter highlights the limitations of epifluorescence imaging, which did not reflect accurately the signal differences between larger tumors due to limited tissue penetration and absorption within thicker tissues. Harvested tumors were first qualitatively imaged under daylight (Fig. 10d) and then weighed (Fig. 10e), upon which the significant growth differences between the Ba/F3 + T766M vs. Ba/F3 + T766M + H566F tumors were evident, as were the other differences already seen in vivo in Fig. 10a. Moreover, we subjected harvested tumor tissues to histology to demonstrate tumor morphology by H&E (Fig. 10f) and pan-EGFR expression by anti-EGFR staining (Fig. 10g vs. staining control in Supplementary Fig. 10f). All tumors were positive for EGFR with plasma membrane localization clearly visible in stained tissues (Fig. 10g). Receptor copy numbers of ~2×105 per cell were estimated by comparison with tumor xenografts from other cell lines (Supplementary Fig. 10g). Our results are consistent with previous intravital fluorescence imaging data that showed predominant localization of EGFR at the plasma membrane consistent with very low ligand binding49.

a Tumor growth measurements by calipers: immunodeficient (NSG) mice were used to establish tumors in their flanks and tumor establishment and cumulative growth was monitored using calipers. N = 6 biologically independent animals per cohort, mean values plotted with error bars representing standard deviation (SD). b IVIS imaging of all animals from (a) at the experimental endpoint (day 41) showing epi-fluorescence signals from animal tumors. Radial efficiency was calculated as 1010(p/s/cm2/sr)/(μW/cm2). c GFP fluorescence signal quantification of tumors corresponding to N = 6 biologically independent animals in b. Statistical analysis by 1-way ANOVA (α = 0.05 and Tukey’s multiple comparison correction). * = 0.012; ** = 0.0029; *** = 0.0001-0.0006 (Supplementary Table 5). Data are presented as mean values; error bars are SD. d Harvested tumors from animals with established tumors were photographed under daylight. No tumors had established in the cohort that had received Ba/F3 + WT cells. e Tumor weights from harvested tumors of N = 6 biologically independent animals. Statistical analysis by 1-way ANOVA with Tukey’s multiple comparison correction: * = 0.0416; ** = 0.0013; **** <0.0001 (Supplementary Table 5; data are presented as mean values; error bars are SD). f Hematoxylin and eosin staining of tumors; scale bars are 100 μm. g Immunohistochemistry staining using a pan anti-EGFR antibody; corresponding background control staining is shown in Supplementary Fig. 10f. Scale bars are 100 μm. Source data are provided as a ‘Source data’ file.
Notably, the T766M + H566F mutant, which does not reduce basal phosphorylation compared with T766M (Supplementary Fig. 7c), also handicaps tumor growth. An explanation is that T766M + H566F expressing Ba/F3 cells are unable to phosphorylate and activate AKT signaling to the same extent as T766M (Supplementary Fig. 7c).
Discussion
We combined super-resolution FLImP imaging with in silico modeling and mutagenesis to identify the interfaces that assemble ligand-free, kinase active EGFR oligomers. The <3 nm resolution achieved allowed us not only to fingerprint previously proposed ligand-free dimer conformers but, crucially, show how interactions between kinase inactive dimers assemble ligand-free heteroconf-oligomers that bear active St2Stect/Asymkindimer sub-units. From this knowledge and the known shapes of the ligand-free dimer conformers we derived a structural model of ligand-free heteroconf-oligomerization that explains how EGFR can achieve ligand-independent auto-phosphorylation. In addition, the extracellular part of this model was validated by implementing a 2D FLImP version.
The tethered conformation of EGFR’s ectodomain appeared during the evolution of vertebrates68 and was suggested to have evolved to prevent crosstalk between the different EGFR homologs in the vertebrate EGFR family69. Our MD simulations and FLImP results propose another biological role for the tethered conformation, which is to regulate the formation of the H2Hectdimer, and through it, heteroconf-oligomer size, receptor activation, and, in the pathological context of T766M-EGFR, tumor formation.
Comparing models and data revealed that a previously orphan Bb2Bbkininterface reported by X-ray crystallography47 plays a regulatory role in ligand-free heteroconf-oligomerization and activation. The role of the Bb2Bbkininterface is to buttress the Asymkindimer, leading to activation once the B2Bect/H2Hkindimer cantilevers H2Hectdimer/2xkinmonomers into position to form the St2Stect/Asymkindimer. This mechanism explains how the Asymkindimer can form in the absence of ligand-induced conformational changes that are typically required to overcome the activation barrier associated with the formation of the asymmetric interface between activator and receiver kinases42. Importantly, our work shows that Ex20Ins mutations stabilize the Bb2Bbkininterface, providing a breakthrough in our understanding of these mutations, which so far have been studied only at the monomeric level.
Comparing models and data also suggested a second S2Skininterface that we identified from a crystal contact in a structure of the H2Hkindimer54. This interface allows the H2Hkindimer to hijack kinase monomers, preventing formation of the stimulatory Bb2Bbkininterface, and down-regulating activation. The S2Skininterface stabilizes the H2Hkindimer, explaining its autoinhibitory role. We propose that the regulation of the activation of ligand-free heteroconf-oligomers rests on balancing the interactions between the Asymkindimer and the H2Hkindimer with the Bb2Bbkininterface and the S2Skininterface.
Beyond its autoinhibitory role, the H2Hkindimer also acts as a scaffold promoting the formation of larger heteroconf-oligomers. Thus, by stabilizing the H2Hkindimer, the T766M mutation decouples oligomer growth from its dependence on the autoinhibitory S2Skininterface. This indirectly underpins the formation of Asymkindimer units without the need to hijack monomers from the Bb2Bbkininterface, which can thereby still buttress Asymkindimer sub-units within larger oligomers, thus amplifying ligand-independent phosphorylation for the T766M receptor mutant.
As an example, we used T766M-EGFR-dependent cell growth to evaluate in vivo the effect of disrupting ligand-free heteroconf-oligomer structure by inhibiting the tethered conformation and the Bb2Bbkininterface. Excitingly, disrupting the Bb2Bbkininterface, which has a deleterious effect on oligomer size and phosphorylation, almost abolishes tumor growth. A clue to a possible mechanism is provided by previous in vivo intravital microscopy studies that proposed that the small pool of high-affinity EGFRs drives tumor xenograft growth when stimulated by the 17–100 pM ligand concentration surrounding the tumor cells49. We found that St2Stect/Asymkindimer sub-units are high-affinity ligand binding sites. According to our model, oligomers display ~25–30% high-affinity St2Stect/Asymkindimer sub-units. In CHO cells expressing ~105 EGFR copies/cell we estimated that ~15-40% of cell surface EGFRs are incorporated in ligand-free oligomers. Similar oligomer fractions would be expected for Ba/F3 + T766M tumor xenograft cells, which we found to express ~2 × 105 EGFR copies/cell70. Among the oligomer species, ~1/2 are hexamers and larger oligomers36 that can bear at least one St2Stect/Asymkindimer sub-unit. Thus, our model predicts that ~2–6% of the total sites will be high-affinity St2Stect/Asymkindimer sub-units, consistent with previous predictions71,72,73. T766M-EGFR oligomers are larger and more stable36, displaying more high-affinity St2Stect/Asymkindimer sites. The finding that inhibition of the Bb2Bbkininterface, which abolishes the assembly of high-affinity St2Stect/Asymkindimer sub-units, also abolishes tumor growth is therefore consistent with the premise that high-affinity sites drive tumor growth when stimulated by ligand. We note that the decreased accessibility of the ligand to its binding site in H566F-EGFRs might also explain the partial effect of this mutation in reducing tumor growth.
By revealing how tumor-driving high-affinity St2Stect/Asymkindimer sub-units assemble within oligomers, our work has also revealed an Achilles heel in drug-resistant NSCLC tumors that could be therapeutically targeted, counterintuitively, with drugs or small proteins that interfere with the tether, and/or can inhibit the Bb2Bbkininterface. Furthermore, these interfaces are away from the ATP-binding pocket mutational hotspot. Targeting protein-protein interactions is an alternative direction in treating diseases and an essential strategy for drug development74. Given the poor long-term efficacy of current treatments for NSCLC, the structural understanding from this work of how T766M-EGFR and Ex20Ins-EGFR amplify cell growth suggests a possible route for more effective therapies. Notably, mutant, and WT-EGFR share heteroconf-oligomer structure. It would be interesting to find out whether these principles apply to cancers driven by EGFR overexpression which currently have very limited therapeutic options.
Methods
Reagents
Antibodies and reagents were purchased as follows: mAb-2E9 (Abcam ab8465, RRID: AB_2096462), Anti-EGFR Affibody® Molecule (Abcam ab95116; RRID: AB_11156238), Anti-EGFR (D38B1, Cell Signaling Technology (CST) 4267; RRID: AB_2246311), Anti-Phospho-Akt (Ser473, D9E, CST 4060; RRID: AB_2315049), Anti-beta-Actin Monoclonal Antibody, HRP Conjugated (13E5 CST 5125; RRID: AB_1903890), Anti-Phospho-EGF Receptor (Tyr1068, D7A5 CST 3777; RRID: AB_2096270), Anti-EGFR (R & D Systems, AF231; RRID: AB_355220), Anti-EGFR (phospho Y992, EM-12, Abcam ab81440; RRID: AB_1658463), Anti-mouse IgG-HRP (Jackson ImmunoResearch 715-035-150; RRID: AB_2340770), Anti-rabbit IgG-HRP (Jackson ImmunoResearch 711-035-152; RRID: AB_10015282), Anti goat IgG-HRP (Jackson ImmunoResearch 705-035-147; RRID: AB_2313587), Anti-Rabbit-HRP (Dako P044801, RRID: AB_2617138), Recombinant Murine EGF (Peprotech, 315-09), Recombinant Murine IL-3 (Peprotech, 213-13), Erlotinib Hydrochloride (Biovision, 1558-100), Lapatinib Ditosylate (Biovision, 1642-25), 4% Paraformaldehyde (EM Grade, Electron Microscopy Service (EMS), 157-4), 25% Glutaraldehyde (Grade I, Sigma G5882), FluoSpheres™ (carboxylate modified, 0.1 μm, infrared (715/755), Invitrogen F8799).
Cell culture
mAb-108 expressing hybridoma cells (ATCC, HB-9764) were grown in high glucose DMEM media, no phenol red (Gibco, 31053044) supplemented with 10% fetal bovine serum (FBS – Gibco, 10270-106) and 1 mM sodium pyruvate (Gibco, 11360070). Chinese Hamster Ovary (CHO) cells (gift from Prof. Peter Parker at The Francis Crick Institute, UK) were grown in DMEM/F12 with no phenol red (Gibco, 21041-025) + 10% (v/v) FBS + 1% penicillin/streptomycin (Gibco, 15140148) and when made to stably express EGFR or mutants the media was supplemented with 4 µg/mL puromycin (Gibco, A1113803) to maintain expression. CHO cells expressing ΔC-EGFR (gift from Prof. Linda Pike, Washington University School of Medicine, USA) were grown in phenol-red-free DMEM supplemented with 10% FBS, 2 mM glutamine (Gibco, 25030081), 1% penicillin/streptomycin, 100 µg/mL hygromycin (Gibco, 10687010) and 100 µg/mL geneticin (Gibco, 10131035). Ba/F3 cells (Creative Biogene, CSC-C2045) stably expressing EGFR or EGFR mutants were grown in RPMI1640, no phenol red (Gibco, 11835063), with 2 mM L-glutamine and without HEPES + 10% heat-inactivated FBS (Gibco, 10500064) + 10 ng/mL mouse IL3 (Peprotech, 213-13) + 1% penicillin/streptomycin + 2 µg/mL puromycin. All cell lines were verified negative for mycoplasma before use and tested routinely.
Plasmid construction & mutagenesis
Point mutations in EGFR plasmids were introduced using Quikchange Lightning Site-directed mutagenesis kit (Agilent Technologies – cat. no. 210518-5) using primer pairs listed in Supplementary Table 4. All constructs were verified by sequencing the whole coding sequence of EGFR. To generate EGFR PiggyBac plasmids, WT-EGFR was PCR amplified from WT-EGFR/pcDNA3 plasmid using primer pairs with NheI and NotI restriction enzyme sites (Supplementary Table 4) and inserted into PB513B-1 vector using standard molecular biology techniques.
Purification of mAb-108 from cell culture media
mAb-108 antibody was purified from mAb-108 hybridoma cell culture supernatant using a mouse TCS antibody purification kit (Abcam, ab128749). Purified antibody was quantified using a Nanodrop and stored at 4 °C.
Generation of EGFR CHO stable cell lines
CHO cells expressing one of the following were generated: WT-EGFR, ED/RK-EGFR, T766M-EGFR, T766M + K946E-EGFR or T766M + I942E-EGFR. CHO cells were transfected with a mix of 0.1 μg of Super Piggybac Transposase expression vector (PB210PA-1, System Biosciences) and 1 μg of the appropriate endotoxin-free EGFR plasmid DNA (in PB513B-1 vector, System Biosciences) using FuGENE HD at 1:3 DNA:FuGENE HD ratio according to manufacturer’s instructions. The cells were selected in fresh media containing 4 μg/mL puromycin for 7-10 days. Surviving clones of cells were pooled and checked for EGFR expression by western blotting and confocal imaging.
Generation of Ba/F3 stable cell lines
Ba/F3 cell lines were electroporated using a Neon transfection kit (Invitrogen, MPK10096) and MicroPorator device (Invitrogen). On the day of electroporation, cells were washed once in PBS and resuspended in buffer R at a final concentration of 1.5 × 107 cells/mL. 100 μL cells ( = 1.5 × 106 cells) were electroporated with a mix of 0.5 μg of Super Piggybac Transposase expression vector and 5 μg of the appropriate endotoxin-free EGFR plasmid DNA according to the manufacturer’s instructions. Cells were selected in media containing 2 μg/mL Puromycin at a density between 0.3 to 1 × 106 cells/mL for 7–10 days to obtain polyclonal cell populations stably expressing EGFR.
Ba/F3 IL3-independent growth assay
Ba/F3 cells stably expressing EGFR or EGFR mutants were washed once in PBS and resuspended in Ba/F3 media without IL3 and puromycin. Cells were grown in the absence of IL3 for 5 days, then seeded at a density of 20,000 cells (in 100 μL of media) per well in white-bottomed 96-well plates in triplicate. As a positive control for cell growth, additional cultures of each cell line were maintained in complete media (including 10 ng/mL recombinant murine IL3). These cells were seeded as above in the presence of IL3. Cell viability was measured every day for 4 days for the positive controls, or 10 days for the cells without IL3, using CellTiter-Glo Luminescent Assay (Promega, G7572) and a CLARIOstar plus microplate reader according to manufacturer’s instructions.
FLImP sample Affibody labeling
CHO cells stably expressing WT-EGFR or mutant EGFR were seeded in 3 of the central 4 wells of μ-Slide 8 well high glass bottom slides (ibidi, 80807), coated with 1% BSA, at 1.8 × 104 cells per well and allowed to grow for 2 days. The 4th well (top left) was coated with poly-L-Lysine (PLL – Sigma P4707-50ml) only. ΔC-EGFR-expressing CHO cells were grown in the presence of 50 ng/ml doxycycline (ThermoFisher Chemicals, J67043.AD). Cells were cultured for 48 h prior to labeling.
Transient transfections of WT CHO cells using FuGENE HD (Promega, E2312) and 600 ng plasmid DNA, at 1:3 DNA:FuGENE ratio, were necessary for the following EGFR mutants: H566F, Lzip3S, ED/RK + L680N, ED/RK + L680N + Lzip3S, K946E, insNPG, G564P, G564P + ED/RK, Lzip3A, T766M + Lzip3S and T766M + I942E. Transfections were carried out 24 h after seeding and left a further 24 h prior to labeling.
Samples were starved in low serum medium (0.1% FBS), with 1 µM Lapatinib or Erlotinib if necessary, for 2 h before rinsing with PBS (PBS without Ca2+ and Mg2+ was used throughout). Samples were then chilled on ice at 4 °C for 10 min in PBS and labeled with 8 nM HER1 Affibody-CF640R, with 1 µM Lapatinib or Erlotinib if necessary, for 1 h on ice at 4 °C. Cells were rinsed and fixed with 3% paraformaldehyde plus 0.5% glutaraldehyde for 15 min on ice and 15 min at room temperature (RT).
Cells were rinsed with PBS and stained with 1 µg/mL Hoechst (Invitrogen, H21492) in PBS at RT for 10 min then rinsed again with PBS. Samples were stored at 4 °C and prior to imaging slides were brought up to RT. PLL was removed from the top left well and PBS from the samples wells. In all wells, 300 µLs of FluoSpheres™, 0.1 µm, infrared (715/755) (Invitrogen F8799) were added (1/50,000 dilution of stock in PBS) to be used as fiducials.
Samples were loaded onto the ONI nanoimager microscope so that the central dividing cross between the central 4 wells of the slide corresponded to position 0,0. The slide was warmed to 34 °C prior to sample collection and the sample was brought into initial focus by the operator.
FLImP sample EGF labeling
Samples were labeled as above except for the following: for mAb-108 and mAb-2E9 treatment, CHO cells expressing WT-EGFR were starved of serum for 2 h then treated with 200 nM mAb on ice at 4 °C for 2 h. For 2D FLImP triangles, CHO cells expressing T766M-EGFR were serum starved for 2 h. Cells were fixed in 3% paraformaldehyde in PBS for 15 min at RT and rinsed. WT-EGFR cells and T766M-EGFR cells were labeled with 10 or 20 nM EGF-CF640R respectively for 1 h at RT and rinsed with PBS. The samples were fixed again for 15 min at RT with 3% paraformaldehyde plus 0.5% glutaraldehyde.
FLImP data acquisition
Image acquisition was performed using an Nanoimager S (ONI Oxford, UK) single molecule imaging microscope with a 1.49 N oil immersion objective, operating NanoImager software (Version: 1.7.3.10248–ef4ff2c0). Single frame FLImP acquisitions used an 8 mW 640 nm diode laser for 20 ms exposure, single frame Hoechst acquisitions used an 8 mW 405 nm diode laser with 20 ms exposure.
The TIRF angle was set, and the instrument temperature was maintained at 34 °C. Data acquisition was facilitated using the PythONI (Python API from ONI) included with the NanoImager software suite. Automated data acquisition procedures are described in Supplementary Note 2. Fiducials in the top left well of each plate were used to estimate PSF properties from a 3 × 3 frame region of interest (ROI). The focal plane was established using the provided ONI autofocusing software (followed by a focal polishing step as described in Supplementary Note 2).
Western blot
CHO cells, 24 h post-transfection with FuGENE HD as described above, were serum starved in media containing 0.1% FBS for 2 h at 37 °C. Cells were placed on ice, washed in cold PBS and lysed directly in 6-well dishes in cell lysis buffer [50 mM Tris/HCl (pH 7.4), 1 mM EDTA, 1 mM EGTA, 50 mM sodium fluoride, 5 mM sodium pyrophosphate, 10 mM sodium β-glycerol 1-phosphate, 1 mM dithiothreitol, 1 mM sodium orthovanadate, 0.27 M sucrose, 1% (v/v) Triton X-100, 1x Protease inhibitor]. Cell extracts were clarified by centrifugation, and the protein concentration was determined using Bradford assay. 10–20 µg of total cell lysate was resolved on an 8% Bolt Bris-Tris gel, and proteins were transferred to PVDF membrane. Membranes were blocked and probed with primary and secondary-HRP antibodies according to manufacturer’s instructions. Subsequently, membranes were incubated with Immobilon ECL Ultra Western HRP substrate solution (Millipore WBKLS0500) and imaged on Biorad Chemidoc MP Imager. Images were quantified in ImageLab software (Biorad) where intensity of each band was relatively quantified against WT-EGFR which was set to 1.
Single particle tracking
Cells were seeded on 1% BSA-coated 35 mm no. 1.5 (high precision) glass-bottomed dishes (MatTek) in 2 mL of media. After 24 h, transient transfections were performed using FuGENE HD as described above, and cells were grown for further 24 h. Prior to imaging, cells were starved for 2 h at 37 °C in 0.1% FBS. Cells were then rinsed twice with 0.1% FBS pre-heated at 37 °C and were labeled with a 1:1 mixture of 8 nM Affibody- Alexa 488 / Affibody- CF640R for 7 min at 37 °C. Cells were rinsed twice with low serum medium pre-heated at 37 °C and promptly imaged as described previously36,75. Typically, for each condition, at least 30 field of views comprising one or more cells were acquired from a total of at least 3 independent biological replicates.
All single-molecule time series data (for FLImP and single particle tracking) were initially analyzed using the multidimensional analysis software described previously76. The colocalization event duration analysis was performed as before36.
Confocal imaging
For all confocal experiments, cells expressing WT-EGFR, L680N-EGFR or ED/RK-EGFR were seeded, serum-starved for 2 h as described above and rinsed twice in ice-cold PBS and cooled down for 10 min on ice.
For anti-EGFR Affibody and EGF competition binding experiments, cells were then pre-treated with either 200 nM, 400 nM or 600 nM of ice-cold Affibody-CF640R in PBS or with mock treatment (PBS) for 1 h at 4 °C, rinsed with ice-cold PBS and fixed with 3% paraformaldehyde in PBS for 30 min at 4 °C. After fixation, cells were rinsed again with RT PBS and labeled with 400 nM EGF-Alexa488 in PBS for 1 h at RT, rinsed with RT PBS, then fixed with 3% paraformaldehyde + 0.5% glutaraldehyde for 15 min at RT.
For the mAb-2E9 (Abcam, ab8465, RRID: AB_2096462) binding experiments, after starvation cells were pre-treated with 200 nM of mAb-2E9-AF488 in PBS or with mock treatment (PBS) for 2 h at 4 °C, rinsed with ice-cold PBS and fixed with 3% paraformaldehyde in PBS for 15 min at 4 °C. For the EGF binding post-fixation test, cells were labeled with 200 nM EGF-CF640R for 2 h at 4 °C then fixed.
After fixation, cells were rinsed again with RT PBS and labeled with 200 nM EGF-CF640R in PBS for 1 h at RT, then rinsed with PBS, fixed with 3% paraformaldehyde + 0.5% glutaraldehyde for 15 min at RT and rinsed with PBS. All samples were stored in PBS at 4 °C until the time of acquisition, and allowed to pre-warm at RT, before loading on the microscope.
Image acquisition was performed on an Elyra PS1, using Zen Black v2.3 SP1 using 633 nm or 488 nm laser excitation.
Colocalization analyses were carried out on images of 600 μm optical slices and performed using Huygens software (Scientific Volume Imaging).
For the mAb-2E9 binding experiments, pixel-wise intensity or intensity ratio distributions were extracted from the data using Huygens (SVI). The non-parametric Kruskal-Wallis statistical test was performed and T-test post-hoc analysis with Bonferroni multiple comparison correction was applied to calculate P Values in Python.
Mice tumor models
In this study, 24 young adult male (6-7 weeks old, 24.6 ± 2.1 g) were used for all animal experiments (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ mice mice, Charles River UK, Strain code: 614; RRID:IMSR_JAX:005557). All mice were maintained within the King’s College London Biological Services Unit under specific pathogen-free conditions in a dedicated and licensed air-conditioned animal room (at 23 ± 2 °C and 40-60% relative humidity) under light/dark cycles lasting 12 h every day. They were kept in individually ventilated standard plastic cages (501cm2 floor space; from Tecniplast) including environmental enrichment and bedding material in the form of sterilized wood chips, paper stripes and one cardboard roll per cage. Maximum cage occupancy was five animals, and animals were moved to fresh cages with fresh environmental enrichment and bedding material twice per week. Sterilized tap water and food were available ad libitum; food was PicoLab Rodent Diet 20 (LabDiet) in the form of 2.5 × 1.6 × 1.0 cm oval pellets that were supplied at the top of the cages.
Male NSG mice were used to establish subcutaneous tumor models (in right flanks) with indicated stable Ba/F3 cell lines. Males were used because lung cancer has a higher incidence in human males. After acclimatization, mice were randomly allocated into four cohorts with six individuals each, shaved on their flanks, and then each received 2 × 106 tumor cells suspended in 100 μL PBS subcutaneously. Tumor growth was followed by calipers and tumor volumes calculated. Tumor models were grown to compare tumor growth between cohorts. The experimental endpoint was defined by the time the humane endpoint was reached for the cohort with the largest tumor growth, and then all animals were sacrificed. All experimental protocols were monitored and approved by the King’s College London Animal Welfare and Ethical Review Body in accordance with UK Home Office regulations (Project License PP4067431) under the Animals (Scientific Procedures) Act 1986 and UK National Cancer Research Institute (NCRI) Guidelines for the Welfare and Use of Animals in Cancer Research. The mice were treated according to the endpoints stated in the Project License. The mice were well throughout, and the tumor volumes were established using the formula for an ellipsoid, V = ½LxW2. The volumetric size limit is 1.5 cm3 which determined the end point of the experiment.
In vivo imaging of tumor models
In vivo GFP fluorescence imaging of superficial tumor models was performed to visualize tumor growth in some animals per group over time and to quantify tumor growth differences in all animals at the experimental endpoint. ROIs were manually drawn including the whole tumor (or the injection sites where no tumors were visible) and used to calculate the radiant efficiency. Prism software version 9 (GraphPad, La Jolla, USA) was used to calculate all statistical parameters as indicated. Generally, p-values were calculated using significance levels of α = 0.05. In-text numbers indicate means of pooled data ± standard deviation (SD) unless otherwise stated.
Tissue staining and histologic analysis
Formaldehyde-fixed paraffin-embedded (FFPE) tissues were prepared using standard methods as described in77. Morphologic analysis of tumor tissues was performed on hematoxylin- and eosin-stained sections. For antibody staining, sections were blocked (Dual Endogenous Enzyme Blocking Reagent, Dako, S200389-2) in 1% (w/v) BSA for 60 min at RT, incubated with primary antibodies (Anti-EGFR, D38B1, CST 4267; RRID: AB_2246311) at 4 °C overnight and secondary antibody (2 µg/mL Anti-Rabbit Ig-HRP, Dako P044801 in Tris Borate Saline - TBS) for 60 min at RT. Samples were developed (using Liquid DAB+ Substrate Chromogen System, Dako, K3467) and counterstained with hematoxylin before mounting. Slides were scanned using a Nanozoomer (Hamamatsu, Japan) with images being analyzed and processed by ImageJ.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
FLImP and single particle tracking data generated in this study are available within this paper and upon request from the Lead Contact. Source data are provided with this paper. The confocal data generated in this study to assess binding affinities have been deposited in the Zenodo database under accession code https://doi.org/10.5281/zenodo.1056724878 and the input files for the MD simulations are deposited in YARETA under accession code https://doi.org/10.26037/yareta:qtkuoibmhndc3jxcwtwzo7eeey79. Source data are provided with this paper.
Code availability
The code used in this study is available upon request from the Lead Contact.
References
Lemmon, M. A., Schlessinger, J. & Ferguson, K. M. The EGFR family: Not so prototypical receptor Tyrosine Kinases. Cold Spring Harb. Perspect. Biol. 6, a020768–a020768 (2014).
Lemmon, M. A. & Schlessinger, J. Cell signaling by receptor Tyrosine Kinases. Cell 141, 1117–1134 (2010).
Zhang, X., Gureasko, J., Shen, K., Cole, P. A. & Kuriyan, J. An allosteric mechanism for activation of the kinase domain of epidermal growth factor receptor. Cell 125, 1137–1149 (2006).
Wee, P. & Wang, Z. Epidermal growth factor receptor cell proliferation signaling pathways. Cancers 9, 52 (2017).
Liang, S. I. et al. Phosphorylated EGFR dimers are not sufficient to activate ras. Cell Rep. 22, 2593–2600 (2018).
Needham, S. R. et al. EGFR oligomerization organizes kinase-active dimers into competent signalling platforms. Nat. Commun. 7, 13307 (2016).
SEER*Explorer: An interactive website for SEER cancer statistics. https://seer.cancer.gov/statistics-network/explorer/ (2023).
SEER Incidence Data, November 2022 Submission (1975-2020), SEER 22 registries (excluding Illinois and Massachusetts). Expected Survival Life Tables by Socio-Economic Standards. (2022).
Mok, T. S. et al. Gefitinib or Carboplatin–Paclitaxel in pulmonary Adenocarcinoma. N. Engl. J. Med. 361, 947–957 (2009).
Rosell, R. et al. Screening for epidermal growth factor receptor mutations in lung cancer. N. Engl. J. Med. 361, 958–967 (2009).
Douillard, J.-Y. et al. First-line gefitinib in Caucasian EGFR mutation-positive NSCLC patients: a phase-IV, open-label, single-arm study. Br. J. Cancer 110, 55–62 (2014).
Gazdar, A. F. Activating and resistance mutations of EGFR in non-small-cell lung cancer: role in clinical response to EGFR tyrosine kinase inhibitors. Oncogene 28, S24–31 (2009).
Vyse, S. & Huang, P. H. Targeting EGFR exon 20 insertion mutations in non-small cell lung cancer. Signal Transduct. Target. Ther. 4, 1–10 (2019).
Kuiper, J. L. et al. Incidence of T790M mutation in (sequential) rebiopsies in EGFR-mutated NSCLC-patients. Lung Cancer 85, 19–24 (2014).
Wang, Z.-F., Ren, S.-X., Li, W. & Gao, G.-H. Frequency of the acquired resistant mutation T790 M in non-small cell lung cancer patients with active exon 19Del and exon 21 L858R: a systematic review and meta-analysis. BMC Cancer 18, 148 (2018).
Wu, S.-G. et al. The mechanism of acquired resistance to irreversible EGFR tyrosine kinase inhibitor-afatinib in lung adenocarcinoma patients. Oncotarget 7, 12404–12413 (2016).
Del Re, M. et al. Concise review: resistance to tyrosine kinase inhibitors in non-small cell lung cancer: the role of cancer stem cells. STEM CELLS 36, 633–640 (2018).
Yun, C.-H. et al. The T790M mutation in EGFR kinase causes drug resistance by increasing the affinity for ATP. Proc. Natl Acad. Sci. USA. 105, 2070–2075 (2008).
Carmi, C. et al. Irreversible inhibition of epidermal growth factor receptor activity by 3-Aminopropanamides. J. Med. Chem. 55, 2251–2264 (2012).
Harvey, R. D., Adams, V. R., Beardslee, T. & Medina, P. Afatinib for the treatment of EGFR mutation-positive NSCLC: a review of clinical findings. J. Oncol. Pharm. Pract. 26, 1461–1474 (2020).
Sequist, L. V. et al. Neratinib, an irreversible Pan-ErbB receptor tyrosine kinase inhibitor: Results of a Phase II trial in patients with advanced non–small-cell lung. Cancer J. Clin. Oncol. 28, 3076–3083 (2010).
Soria, J.-C. et al. Osimertinib in untreated EGFR-mutated advanced non-small-cell lung cancer. N. Engl. J. Med. 378, 113–125 (2018).
Carlisle, J. W. & Ramalingam, S. S. Role of osimertinib in the treatment of EGFR-mutation positive non-small-cell lung cancer. Future Oncol. Lond. Engl. 15, 805–816 (2019).
Le, X. et al. Landscape of EGFR-dependent and -independent resistance mechanisms to osimertinib and continuation therapy beyond progression in EGFR-mutant NSCLC. Clin. Cancer Res. J. Am. Assoc. Cancer Res.24, 6195–6203 (2018).
Ramalingam, S. S. et al. Mechanisms of acquired resistance to first-line osimertinib: Preliminary data from the phase III FLAURA study. Ann. Oncol. 29, viii740 (2018).
Papadimitrakopoulou, V. A. et al. Analysis of resistance mechanisms to osimertinib in patients with EGFR T790M advanced NSCLC from the AURA3 study. Ann. Oncol. 29, viii741 (2018).
Wang, S., Song, Y. & Liu, D. EAI045: The fourth-generation EGFR inhibitor overcoming T790M and C797S resistance. Cancer Lett. 385, 51–54 (2017).
Jia, Y. et al. Overcoming EGFR(T790M) and EGFR(C797S) resistance with mutant-selective allosteric inhibitors. Nature 534, 129–132 (2016).
Red Brewer, M. et al. Mechanism for activation of mutated epidermal growth factor receptors in lung cancer. Proc. Natl Acad. Sci. USA 110, E3595–604 (2013).
Bhullar, K. S. et al. Kinase-targeted cancer therapies: progress, challenges and future directions. Mol. Cancer 17, 1–20 (2018).
Pottier, C. et al. Tyrosine Kinase inhibitors in cancer: breakthrough and challenges of targeted therapy. Cancers 12, 731 (2020).
Chakraborty, S. et al. Constitutive and ligand-induced EGFR signalling triggers distinct and mutually exclusive downstream signalling networks. Nat. Commun. 5, 5811 (2014).
Guo, G. et al. Ligand-independent EGFR signaling. Cancer Res. 75, 3436–3441 (2015).
Paez, J. G. et al. EGFR mutations in lung cancer: correlation with clinical response to gefitinib therapy. Science 304, 1497–1500 (2004).
Arkhipov, A. et al. Architecture and membrane interactions of the EGF receptor. Cell 152, 557–569 (2013).
Zanetti-Domingues, L. C. et al. The architecture of EGFR’s basal complexes reveals autoinhibition mechanisms in dimers and oligomers. Nat. Commun. 9, 4325 (2018).
Zanetti-Domingues, L. C. et al. Determining the geometry of oligomers of the human epidermal growth factor family on cells with 7 nm resolution. Prog. Biophys. Mol. Biol. 118, 139–152 (2015).
Webb, S. E. D. et al. Nanometric molecular separation measurements by single molecule photobleaching. Methods San. Diego Calif. 88, 76–80 (2015).
Needham, S. R. et al. Determining the geometry of oligomers of the human epidermal growth factor family on cells with <10 nm resolution. Biochem. Soc. Trans. 43, 309–314 (2015).
Needham, S. R. et al. Measuring EGFR separations on cells with ∼10 nm resolution via fluorophore localization imaging with photobleaching. PloS One 8, e62331 (2013).
Sutto, L. & Gervasio, F. Effects of oncogenic mutations on the conformational free-energy landscape of EGFR kinase. Proc. Natl Acad. Sci. 110, 10616–10621 (2013).
Shan, Y. et al. Oncogenic mutations counteract intrinsic disorder in the EGFR kinase and promote receptor dimerization. Cell 149, 860–870 (2012).
Lu, C., Mi, L.-Z., Schurpf, T., Walz, T. & Springer, T. a. Mechanisms for Kinase-mediated dimerization of the epidermal growth factor receptor. J. Biol. Chem. 287, 38244–38253 (2012).
Bessman, N. J., Bagchi, A., Ferguson, K. M. & Lemmon, M. A. Complex relationship between ligand binding and dimerization in the epidermal growth factor receptor. Cell Rep. 9, 1306–1317 (2014).
Lelimousin, M., Limongelli, V. & Sansom, M. S. P. Conformational changes in the epidermal growth factor receptor: role of the transmembrane domain investigated by coarse-grained metadynamics free energy calculations. J. Am. Chem. Soc. 138, 10611–10622 (2016).
Ferguson, K. M. et al. EGF activates its receptor by removing interactions that autoinhibit Ectodomain Dimerization. Mol. Cell 11, 507–517 (2003).
Yoshikawa, S. et al. Structural basis for the altered drug sensitivities of non-small cell lung cancer-associated mutants of human epidermal growth factor receptor. Oncogene 32, 27–38 (2013).
Kovacs, E. et al. Analysis of the role of the C-terminal tail in the regulation of the epidermal growth factor receptor. Mol. Cell. Biol. 35, 3083–3102 (2015).
Pinilla-Macua, I., Grassart, A., Duvvuri, U., Watkins, S. C. & Sorkin, A. EGF receptor signaling, phosphorylation, ubiquitylation and endocytosis in tumors in vivo. eLife 6, e31993 (2017).
Kozer, N. et al. Exploring higher-order EGFR oligomerisation and phosphorylation-a combined experimental and theoretical approach. Mol. Biosyst. 9, 1849–1863 (2013).
Saffarian, S., Li, Y., Elson, E. L. & Pike, L. J. Oligomerization of the EGF receptor investigated by live cell fluorescence intensity distribution analysis. Biophys. J. 93, 1021–1031 (2007).
Dixon, P. M. Bootstrap Resampling. in Encyclopedia of Environmetrics (eds. El-Shaarawi, A. H. & Piegorsch, W. W.) (John Wiley & Sons, Ltd, 2006). https://doi.org/10.1002/9780470057339.vab028.
Vaserstein, L. N. Markov processes over denumerable products of spaces, describing large systems of automata. Ross. Akad. Nauk Probl. Peredachi Informatsii 5, 64–72 (1969).
Jura, N. et al. Mechanism for activation of the EGF receptor catalytic domain by the juxtamembrane segment. Cell 137, 1293–1307 (2009).
Mattoon, D., Klein, P., Lemmon, M. A., Lax, I. & Schlessinger, J. The tethered configuration of the EGF receptor extracellular domain exerts only a limited control of receptor function. Proc. Natl Acad. Sci. USA 101, 923–928 (2004).
Tynan, C. J. et al. Human epidermal growth factor receptor (EGFR) aligned on the plasma membrane adopts key features of Drosophila EGFR asymmetry. Mol. Cell. Biol. 31, 2241–2252 (2011).
Defize, L. H. et al. Signal transduction by epidermal growth factor occurs through the subclass of high affinity receptors. J. Cell Biol. 109, 2495–2507 (1989).
Schmitz, K. R. & Ferguson, K. M. Interaction of antibodies with ErbB receptor extracellular regions. Exp. Cell Res. 315, 659–670 (2009).
Shepherd, F. A. et al. Erlotinib in previously treated non–small-cell lung cancer. N. Engl. J. Med. 353, 123–132 (2005).
Macdonald-Obermann, J. L. & Pike, L. J. Allosteric regulation of EGF receptor ligand binding by tyrosine kinase inhibitors. J. Biol. Chem. 293, 13401–13414 (2018).
Schmitz, K. R., Bagchi, A., Roovers, R. C., van Bergen en Henegouwen, P. M. P. & Ferguson, K. M. Structural evaluation of EGFR inhibition mechanisms for nanobodies/VHH domains. Structure 21, 1214–1224 (2013).
Mi, L.-Z. et al. Simultaneous visualization of the extracellular and cytoplasmic domains of the epidermal growth factor receptor. Nat. Struct. Mol. Biol. 18, 984–989 (2011).
Lecordier, L. et al. C-terminal mutants of apolipoprotein LI efficiently kill both Trypanosoma brucei brucei and Trypanosoma brucei rhodesiense. PLoS Pathog. 5, e1000685 (2009).
Zanetti-Domingues, L. C., Tynan, C. J., Rolfe, D. J., Clarke, D. T. & Martin-Fernandez, M. Hydrophobic fluorescent probes introduce artifacts into single molecule tracking experiments due to non-specific binding. PLoS ONE 8, e74200 (2013).
Yasuda, H. et al. Structural, biochemical, and clinical characterization of Epidermal Growth Factor Receptor (EGFR) Exon 20 insertion mutations in lung cancer. Sci. Transl. Med. 5, 216ra177–216ra177 (2013).
Chakroborty, D. et al. An unbiased in vitro screen for activating epidermal growth factor receptor mutations. J. Biol. Chem. 294, 9377–9389 (2019).
Collins, M. K. et al. Transfer of functional EGF receptors to an IL3-dependent cell line. J. Cell. Physiol. 137, 293–298 (1988).
Liu, P. et al. A single ligand is sufficient to activate EGFR dimers. Proc. Natl Acad. Sci. USA 109, 10861–10866 (2012).
Liu, P., Bouyain, S., Eigenbrot, C. & Leahy, D. J. The ErbB4 extracellular region retains a tethered-like conformation in the absence of the tether. Protein Sci. 21, 152–155 (2012).
Byrne, P. O., Hristova, K. & Leahy, D. J. EGFR forms ligand-independent oligomers that are distinct from the active state. J. Biol. Chem. 295, 13353–13362 (2020).
Ringerike, T. et al. High-affinity binding of Epidermal Growth Factor (EGF) to EGF receptor is disrupted by overexpression of mutant dynamin (K44A) *. J. Biol. Chem. 273, 16639–16642 (1998).
Rees, A. R., Gregoriou, M., Johnson, P. & Garland, P. B. High affinity epidermal growth factor receptors on the surface of A431 cells have restricted lateral diffusion. EMBO J. 3, 1843–1847 (1984).
Sorkin, A., Waters, C., Overholser, K. A. & Carpenter, G. Multiple autophosphorylation site mutations of the epidermal growth factor receptor. Analysis of kinase activity and endocytosis. J. Biol. Chem. 266, 8355–8362 (1991).
Lu, H. et al. Recent advances in the development of protein–protein interactions modulators: mechanisms and clinical trials. Signal Transduct. Target. Ther. 5, 1–23 (2020).
Clarke, D. T. et al. Optics clustered to output unique solutions: A multi-laser facility for combined single molecule and ensemble microscopy. Rev. Sci. Instrum. 82, 93705 (2011).
Rolfe, D. J. et al. Automated multidimensional single molecule fluorescence microscopy feature detection and tracking. Eur. Biophys. J. EBJ 40, 1167–1186 (2011).
Fruhwirth, G. O., Diocou, S., Blower, P. J., Ng, T. & Mullen, G. E. D. A whole-body dual-modality radionuclide optical strategy for preclinical imaging of metastasis and heterogeneous treatment response in different microenvironments. J. Nucl. Med. Publ. Soc. Nucl. Med. 55, 686–694 (2014).
Iyer, R. S. et al. Drug-resistant EGFR mutations promote lung cancer by stabilizing interfaces in ligand-free kinase active EGFR oligomers. Zenodo https://doi.org/10.5281/zenodo.10567249 (2024).
Gervasio, F. L. & Galdadas, I. Drug-resistant EGFR lung cancer mutations promote tumor growth by stabilizing interfaces in ligand-free signaling-competent EGFR oligomers. YARETA https://doi.org/10.26037/yareta:qtkuoibmhndc3jxcwtwzo7eeey (2024).
Endres, N. F. et al. Conformational coupling across the plasma membrane in activation of the EGF receptor. Cell 152, 543–556 (2013).
Lu, C. et al. Structural evidence for loose linkage between ligand binding and kinase activation in the epidermal growth factor receptor. Mol. Cell. Biol. 30, 5432–5443 (2010).
Acknowledgements
We thank Drs Esther Garcia-Gonzalez, Jana Harizanova, Christopher Tynan, Michael Hirsch, Jianguo Rao and Michalis Vrettas for technical support. We thank James Rosekilly and Dr Cheryl Gillett for providing the human tissue TMA that consisted of no/low/high human cancer tissues. We thank Prof Andrew Clayton for his support in collecting the pbICS data. We also thank Dr Michael Hirsch for comments on the manuscript. This work has been funded by grant Ref: ST/S000682/1 from the Science and Technology Facilities Council UK (R.S.I. and B.M.D.) and a joint-funded King’s College London-The University of Hong Kong PhD studentship (R.C.H.M.). M.L.M.-F., D.J.R., and B.M.D. are grateful for significant computing resources and support provided by STFC Scientific Computing Department’s SCARF cluster and its Data Services, Research Infrastructure and Cloud Operations Groups, with funding from STFC’s Ada Lovelace Centre and IRIS eInfrastructure consortium. F.L.G. and I.G. acknowledge the Swiss National Science Foundation and Bridge for financial support (project number: 200021_204795 and 40B2-0_203628). F.L.G. and I.G. also acknowledge the Swiss National Supercomputing Centre (CSCS) for large supercomputer time allocations, project IDs: s1107, s1169, s1228.
Author information
Authors and Affiliations
Contributions
R.S.I., S.R.N., I.G., B.M.D., and S.K.R. contributed equally to this work. Conceptualization, R.S.I., I.G., S.K.R., S.R.N., L.C.Z.-D., P.J.P., D.J.R., B.M.D., D.T.C., F.L.G., and M.L.M.-F.; molecular biology, R.S.I. and S.K.R.; in vitro data acquisition, R.S.I., S.R.N., S.K.R., L.C.Z.-D.; FLImP automation, D.J.R., B.M.D., S.R.N.; algorithm development and data analysis, D.J.R., B.M.D.; MD simulations, I.G. and F.L.G.; in vivo work R.C.H.M., G.O.F., R.S.I., and S.K.R.; writing – original draft, M.L.M.-F.; writing – review & editing, I.G., L.C.Z.-D., R.S.I., S.R.N., D.J.R., B.M.D., G.O.F., P.J.P., S.K.R., D.T.C., F.L.G., M.L.M.-F.; visualization, S.R.N., L.C.Z.-D., B.M.D., D.J.R., S.K.R., D.T.C., I.G., R.C.H.M., R.S.I.; funding acquisition M.L.M.-F., D.J.R., B.M.D., and F.L.G.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Iyer, R.S., Needham, S.R., Galdadas, I. et al. Drug-resistant EGFR mutations promote lung cancer by stabilizing interfaces in ligand-free kinase-active EGFR oligomers. Nat Commun 15, 2130 (2024). https://doi.org/10.1038/s41467-024-46284-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-46284-x
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
