
Abstract
Transcriptional regulator MtrR inhibits the expression of the multidrug efflux pump operon mtrCDE in the pathogenic bacterium Neisseria gonorrhoeae. Here, we show that MtrR binds the hormonal steroids progesterone, β-estradiol, and testosterone, which are present at urogenital infection sites, as well as ethinyl estrogen, a component of some hormonal contraceptives. Steroid binding leads to the decreased affinity of MtrR for cognate DNA, increased mtrCDE expression, and enhanced antimicrobial resistance. Furthermore, we solve crystal structures of MtrR bound to each steroid, thus revealing their binding mechanisms and the conformational changes that induce MtrR.
Similar content being viewed by others
Introduction
The pathogen Neisseria gonorrhoeae is a Gram-negative diplococcus that colonizes genital, rectal, and oral mucosa1. As a strict human pathogen, N. gonorrhoeae is finely adapted for survival in its sole natural host and infects over 85 million individuals (both sexes) each year worldwide2 and infections in the US are far more prevalent in women than men3. No vaccine is available, and reinfection by identical gonococcal strains is possible1,4. After successful colonization by gonococci (GC), a female patient may experience complications arising from infection including pelvic inflammatory disease, ectopic pregnancy, and infertility1. Neonatal blindness caused by maternal transmission during childbirth can also occur without appropriate preventative measures1.
Since the late 1930’s with the availability of sulfonamides followed by penicillin, antibiotics have been used to treat gonococcal infections, but their effectiveness has decreased with the rise of multidrug resistance (MDR)5. To date, N. gonorrhoeae has developed resistance to sulfonamides, macrolides, aminoglycosides, beta-lactams, tetracyclines and fluoroquinolones1,6. Currently, the sole empiric treatment for gonococcal infections in the US is ceftriaxone but alarmingly, ceftriaxone-resistant strains have been identified in Denmark, France, Japan, Thailand, and the United Kingdom7,8,9,10,11,12. This consistent rise in MDR indicates the possibility of untreatable strains of N. gonorrhoeae and has prompted the CDC (Centers for Disease Control and Prevention) to flag this bacterium as an urgent public health threat. Thus, a detailed molecular understanding of gonococcal MDR mechanisms and their regulation are crucial to inform public health decisions7.
N. gonorrhoeae, like most pathogens, contains a number of mechanisms that lead to antibiotic resistance including β-lactam antibiotic resistance by antibiotic modifying proteins13, ribosome protective proteins14, mutations in penicillin binding proteins6, and reduced permeability of cell walls by outer membrane proteins6,15. Significantly, the direct export of antibiotics and toxic molecules by multidrug efflux pumps is a major contributor to gonococcal survival and MDR. Multidrug efflux pumps decrease the cytosolic and periplasmic concentration of antibiotics and cytotoxins by pumping them into the extracellular milieu16. These systems can recognize and efflux a wide variety of chemically and structurally dissimilar molecules16. The best characterized multidrug efflux system of N. gonorrhoeae is the MtrCDE tripartite system, a member of the Resistance-Nodulation-Division (RND) family1. Studies investigating MDR in N. gonorrhoeae report significant increases in gonococcal susceptibility to antibiotics when the MtrCDE system is impaired or deleted17. Overexpression of the MtrCDE efflux system due to cis- or trans-acting regulatory systems is a leading cause of gonococcal resistance to hydrophobic agents (HAs), fatty acids, and even human antimicrobial peptides such as LL-3717,18,19. However, the energetic cost of synthesizing efflux pumps is significant, and their expression is regulated tightly leading to optimal cell viability20.
The mtrCDE operon is regulated by the multiple transferable resistance repressor, MtrR, which also acts as a global transcriptional regulator in N. gonorrhoeae17. Beyond its regulation of the expression of the mtrCDE operon, MtrR directly represses rpoH, which encodes a secondary sigma factor that controls the stress response in gonococci21,22,23. MtrR can regulate these genes by binding a degenerate consensus sequence upstream of their coding regions thus blocking access of RNA polymerase to the promotor17,21. Upon binding to cytotoxic molecules and likely sensing oxidative stress, MtrR undergoes a conformational change to relieve repression of its target genes (Fig. 1A)18.

A MtrR bound to the mtrCDE operator site blocks recognition of the −35 and −10 elements by RNA polymerase thereby precluding transcription of the mtrC, mtrD, and mtrE genes, which encode the periplasmic accessory, efflux, and outer membrane channel proteins, respectively, of the MtrCDE efflux system. The dark blue arrows depict the coding regions of each gene. The transcription start site is represented by a bent arrow. Transcriptional repression is shown as a red X, cytotoxic molecules are represented as red spheres, and MtrR is represented as a blue surface. B The entirely helical structure of MtrR is shown as cylinders and the biochemical and biological functions of each helix is described. C Structure of the homodimeric MtrR bound to the mtrCDE operator site with the helices colored as in B (PDB: 7JU3)25.
A member of the ubiquitous TetR family, MtrR is entirely alpha helical and comprises a helix-turn-helix (HTH) DNA binding motif and a C-terminal ligand binding/dimerization domain (Fig. 1B, C)18. Mutations in the mtrR gene or its cognate promoters are present in many clinical isolates resistant to antimicrobials and antibiotics24. Recent structural and biochemical studies on the MtrR-rpoH and MtrR-mtrCDE operator complexes reveal the basis of loss of function mutations in the HTH DNA binding motif that abolish regulation of these genes by MtrR25.
Despite these initial studies, little is known about the induction and ligand recognition mechanisms of MtrR by innate human inducers. Previous work from our laboratories has shown that MtrR binds and is induced by the bile salts chenodeoxycholate (CDCA) and taurodeoxycholate (TDCA), found at extra-urogenital GC infection sites, but not glycocholate18. However, physiologically relevant inducers of MtrR that are found at urogenital sites, which form the vast majority of the sites of gonococcal infection, are not known, posing a significant gap in our understanding of how MtrR contributes to MDR and gonococcal survival in the reproductive tract. To address this critical lack of knowledge, we carried out a series of structural, biochemical, and cellular experiments to elucidate the binding and induction mechanisms of MtrR by female and male steroidal hormones as they are physiologically relevant molecules that would confront gonococci during infection. In the female reproductive tract, gonadol hormones are responsible for the coordination of immune cell function, vaginal microbiome, and epithelial cell architecture during menstration to optimize maternal protection against pathogen infection and also fetal implantation and survival26,27,28. This results in a “window of vulnerability” where the immune protection is dampened in the reproductive tract in order to optimize sperm and fetal survival, which can be taken advantage of by pathogens26,27. Thus, pathogens can utilize hormones to regulate gene expression involved in cytotoxic stress defenses allowing host infection26,29,30,31,32,33,34. Mechanisms for sensing and coping with cytotoxic stresses are crucial for survival of all organisms, particularly obligate human pathogens like N. gonorrhoeae that are routinely exposed to cytotoxic attacks by the host and inhabit an environment that is well equipped with antimicrobial defense mechanisms. Elucidation of the functions of MtrR will contribute to the understanding of MDR in N. gonorrhoeae, and transcriptional regulation that promotes pathogen survival. Here, we show that binding of human steroidal hormones results in a comformational change in the MtrR structure rendering it incompatible with DNA binding, thus revealing the structural mechanism of MtrR induction.
Results
Steroidal hormones bind MtrR and reduce its DNA-binding affinity
We first investigated the ability of MtrR to bind selected ligands. Because bacterial regulatory proteins of efflux systems are commonly induced by the substrates of the corresponding pump, allowing increased expression and resistance under cytotoxic stresses, we hypothesized that some ligands of MtrCDE are MtrR inducers35,36,37. Previous work has shown that mutations of the mtrD gene result in a significant decrease in the minimal inhibitory concentration (MIC) of progesterone (PTR) in N. gonorrhoeae38. Other physiologically relevant candidates of MtrR inducers include the gonadal steroids β-estradiol (EST) and testosterone (TES), which, in addition to PTR, have been shown to arrest N. gonorrhoeae growth39. Of these three, PTR is the most effective in growth inhibition, highlighting this compound as a potent protective agent against gonococcal infection. The effect of PTR may also explain the higher occurrence of asymptomatic gonorrheal infection in women than men39. Mutations in the mtrR gene also confer increased susceptibility to PTR in plated cells as well as decreased survival in the lower genital tract of female mice40. Additionally, female mice models are most susceptible to gonococcal infection during periods of the menses cycle when EST levels are highest41, and mice treated with EST showed significantly increased susceptibility to disseminated gonococcal infection42.
We conducted isothermal titration calorimetry (ITC) studies on EST, PTR, and TES to determine their affinities for MtrR. These studies revealed that MtrR bound these steroids with equilibrium dissociation constants (Kd) of 2.75 ± 0.7 µM (PTR), 1.67 ± 1.1 µM (EST), and 2.26 ± 0.3 µM (TES) (Fig. 2A), whilst the chemical precursor cholesterol, the stress hormone cortisol, and the antibiotic azithromycin (AMR), show no binding (Fig S1). The stoichiometry for EST and TES is one molecule of steroid per MtrR monomer and intriguingly one molecule of PTR per MtrR dimer. In addition to these three gonadal steroids, we also tested the binding of ethinyl estradiol (NDR), which is a synthetic estrogen used in some hormonal contraceptives43, and found that it binds MtrR with a Kd of 0.94 ± 0.6 µM (Fig. 2A). These results show the substantial affinity and specificity of MtrR for steroidal hormones. To determine if these identified host ligands result in the biochemical induction of MtrR, we measured the binding affinity of MtrR for the rpoH and mtrCDE operators in the presence and absence of 125 µM PTR, EST, TES, or NDR (in 1% methanol (MeOH)) using a DNA binding fluorescence polarization-based assay (FP). Results from these FP experiments showed up to a 13-fold and 3-fold decrease in binding affinity of MtrR for the rpoH and mtrCDE operator, respectively, in the presence of each steroid compared to binding of MtrR to these sites in the presence of 1% MeOH alone (Fig. 2B, S2). At higher concentrations, MtrR begins to bind nonspecifically to the DNA, possibly explaining why MtrR shows a more modest fold change in binding affinity for the mtrCDE operator compared to rpoH. These data support the hypothesis that gonadal steroids are physiologically relevant inducers of MtrR regulated genes.

A Representative Isothermal titration calorimetry thermograms and binding isotherms of each natural steroid and ethinyl estradiol. At least three experimental measurements, technical and some biological replicates, were averaged for each reported binding constant. B Fluorescence polarization-based DNA-binding isotherms reveal significant decreases in the binding affinity of MtrR for the rpoH and mtrCDE operators in the presence of each gonadal steroid. Data are represented by the mean values (point) +/- SEM (error bar) of at least three independent experimental measurements and were averaged for each reported binding constant.
Structures of MtrR-steroid complexes
We carried out crystallographic studies to determine the binding mechanisms and structural basis of induction of MtrR bound to progesterone, testosterone, β-estradiol, and ethinyl estrogen. The structures of MtrR bound to cognate DNA and an “apo” form have been previously solved in our laboratory18,25. The structure of MtrR bound to EST was determined at a resolution of 2.37 Å using de novo phasing by single wavelength anomalous dispersion after selenomethionine substitution of the protein. Structures of MtrR bound to PTR, TES, and NDR were subsequently solved by molecular replacement at resolutions of 2.31, 2.22, and 3.20 Å, respectively. The Rwork and Rfree values for each structure are 19.2% and 24.4% (MtrR+EST), 20.8% and 26.1% (MtrR+TES), 21.0% and 25.1% (MtrR+PTR), and 20.1% and 27.7% (MtrR+NDR), respectively. The asymmetric unit (ASU) of all crystals contained two homodimers. The root mean square deviation (RMSD) of the pairwise alignments of all corresponding Cα atoms of the dimers within each ASU is 0.29 Å, 0.30 Å, 0.31 Å, and 0.48 Å for the MtrR-EST, MtrR-TES, MtrR-PTR and MtrR-NDR structures, respectively.
MtrR is composed of 210 residues and exists as a biochemical dimer of 48 kDa as confirmed by gel filtration chromatography18,25. Consistent with the apo structure of MtrR, the first 8 to 10 residues and a stretch of 5 to 11 residues between helices α4 and α5 are not observed in the crystal structures18. In all structures, the last observed carboxy-terminal residue is 209 or 210. In agreement with previous structures and other TetR family members, these MtrR-ligand structures are comprised of homodimers, composed of nine α-helices. The residues of each α-helix are: α1: 10–26, α2: 33–39, α3: 44–50, α4: 53–78, α5: 84–102, α6: 103–111, α7: 123–148, α8: 158–180, α9: 185–203 (Fig. 3A). The first three helices form the DNA binding domain which contains a HTH motif (Fig. 3A). Helices α8-α9 form the majority of the dimerization interface18,25. The “induced” conformation of MtrR bound to each steroid is structurally identical as the RMSDs of the alignment of all Cα atoms of each dimer in all steroid-bound structures reveals essentially the same structure: MtrR+EST:MtrR+TES = 0.43 Å, MtrR+EST:MtrR+PTR = 0.22 Å, MtrR+TES:MtrR+PTR = 0.37 Å, MtrR+EST:MtrR+NDR = 0.46 Å, MtrR+NDR:MtrR+TES = 0.50 Å, and MtrR+NDR:MtrR+PTR = 0.47 Å (Fig. 3A).

A Overlay of hormonal steroid-bound MtrR structures with progesterone-colored charcoal, β-estradiol blue, testosterone pale green, and ethinyl estrogen cyan. The helices of one subunit are labeled B Overlay of MtrR bound to β-estradiol, the induced conformation, (blue), and MtrR bound to the mtrCDE operator site, the repressor conformation, (white). C View of MtrR binding to progesterone, β-estradiol, testosterone, and ethinyl estradiol (PTR, EST, TES, and NDR). Selected binding residues are displayed as sticks. A nearby phosphate ion is shown as a sphere. D Electron density maps of each ligand in the MtrR ligand binding pocket looking both parallel and perpendicular to the four-membered ring of each steroid. Oxygen atoms are colored red. The composite 2Fo-Fc omit map contoured to 2.0 σ is shown in gray mesh.
The MtrR-steroid crystal structures reveal that MtrR binds one ligand per protomer as observed in many TetR family members, including the founding member TetR, which binds one molecule of Mg2+-tetracycline per subunit36. However, not all members of the TetR family bind one molecule per protomer including QacR, which notably binds one molecule per dimer35. The stoichiometry of MtrR bound to EST, TES and NDR is 1 ligand:1 protomer. By contrast, only one PTR is unambiguously assigned to bind the MtrR dimer. Inspection of the binding pocket of the non PTR bound-protomers reveals poor density that is possibly CAPS, a component of the crystallization buffer or a second weakly bound PTR. Interestingly, the second dimer in the asymmetric unit contains a progesterone molecule in each binding pocket. The stoichiometry for this MtrR-PTR complex is inconsistent with the ITC experiments (n = 0.5) and the structures (n = 1). This could be in part because MtrR is crystallized in an excessive concentration of steroids which does not reflect its more physiological binding, the apparent progesterone concentration in the ITC experiments is not correct, or that we are observing partial occupancy of each binding site in the crystal structure. Regardless, the MtrR-PTR complex assumes an induced conformation.
It should also be noted that the electron density for PTR was the weakest of the steroidal hormones, possibly indicating more than one binding pose for this steroid. Furthermore, β-estradiol and testosterone are positioned with the five-membered ring pointing towards the dimerization surface whilst progesterone is positioned with the five-membered ring positioned towards the lateral side of the protein (Fig. 3C, D). This indicates a binding pocket that is malleable, allowing different ligands of the same class to bind in multiple orientations, a finding which is consistent with other TetR multidrug binding proteins such as QacR of Staphylococcus aureus and RamR of Salmonella enterica35,44.
The ligand binding pockets of each protein-steroid complex are similar in size with volumes of ~1040 Å3 (EST), ~1130 Å3 (TES), ~1000 Å3 (PTR), and ~900 Å3 (NDR) and consist of a tunnel between helices 4 and 7 that leads to a cavity surrounded by helices 4–9 (Fig S3)45. The ligand binding pocket is mainly comprised of hydrophobic residues with several amino acid residues in proximity to the steroids to provide Van der Waals contacts. Notably, the aromatic side chain of W136 stacks over the rings of each steroid (Fig. 3C). Interestingly, residue W136 in the MtrR-TES structure is found in two conformations within different binding pockets in the ASU, but maintains Van der Waals contacts to the steroid in each pose. Analysis of the composite omit maps of the MtrR-TES crystal structure reveal that these conformations are distributed equally in the asymmetric unit indicating that there is no significant preference for either indole ring conformation. No positively charged side chains are found in the core of the binding pocket, however, three residues, K167, R176’ (‘ indicating the residue is from the other protomer of the dimer), and R137 coordinate a phosphate ion outside the ligand binding pocket in all subunits, although the density is ambiguous in the MtrR-NDR structure, likely due to the limited resolution of the structure, and was thus omitted in the refinement step for this structure. W136 and R176 were previously shown to be important in binding the bile salt chenodeoxycholate18. One negatively charged amino acid residue, D171 on helix α8 near the dimerization surface is positioned ~3.4 Å from the hydroxyl group on the five-membered ring of testosterone and estradiol and ~3.8 Å away from the carbonyl on C3 of progesterone. The proximity of the side chain of D171 and the hydroxyl groups of estradiol and testosterone, indicated a role for this MtrR residue in ligand specificity. In addition to D171, the side chain of glutamine residue, Q133, adopts multiple conformations that allow it to engage in hydrogen bonds to the carbonyl 1 of progesterone and the hydroxyl group of testosterone (Fig. 3C). As noted, the ligand binding pocket is predominantly formed by non-polar and aromatic residues that engage in hydrophobic contacts with the ligands. Residues that are involved in contact with all four steroids are F95, W136, I174, T91, I140, L170, and L92 (Fig S4).
Superposition of the structures of steroid-bound MtrR and MtrR bound to the rpoH operator sequence reveals a significant conformational change in the DNA binding domain (Fig. 3B). The center-to-center distance between the “recognition” helices of the HTH motifs in the DNA bound structure is 36.8 Å. This distance is increased to 45.0, 45.2, 45.6, and 45.2 Å for MtrR bound to PTR, TES, EST, and NDR, respectively. The RMSD for the overlay of these structures to DNA-bound MtrR are 1.10 (EST:DNA), 1.68 (TES:DNA), 0.96 (PTR:DNA), and 1.98 (NDR:DNA). The alignment of MtrR-EST and MtrR-DNA structures reveals three major movements that result in the induction of the protein (Fig S5 and Supplementary Movie 1)46. The first movement is a 20.8° rotation of the DNA binding domain (α1 through α3), which shifts up and away from the DNA major groove in all MtrR-steroid structures (Fig. 3B). The second significant movement is a 10° rotation and 4.5 Å shift upward of helix α4. This translation of helix α4 coincides with the upward movement of the HTH domain, away from the DNA, as well as a widening of the ligand binding pocket in the induced form. Last, the flexibility in the loop formed by residues 114–122, between helices α6 and α7, allows a 17.2° outward rotation in helix α7. This movement also aids in the enlargement of the ligand binding pocket as well as a movement of residue W136, which is positioned to obstruct the entrance to the ligand binding pocket in the DNA bound form (Fig S3). Previously the structure determination of an “apo” structure of MtrR revealed that it took the identical induced conformation likely due to the presence of CAPS, a component of the crystallization buffer, in the ligand binding pocket18. Superposition of this “apo” structure and the steroid-bound structures results in an RMSD of 0.35 Å (apo-EST), 0.36 Å (apo-TES), 0.28 Å (apo-PTR), and 0.54 Å (apo-NDR).
Ligand binding to MtrR
To confirm the modes of binding of each steroid in the ligand pocket, we made single point mutations in the binding pocket of several residues and tested the ability of each substituted protein to bind each steroid (Fig. 4, S6). Residue D171 on helix α6 appears particularly important for binding and ligand specificity because its carboxy side chain was positioned within proximity to the hydroxy groups on the five-membered ring of TES, EST and NDR. The point mutation D171A was made and binding to each steroid was determined using ITC. Additionally, based on our previous results that showed residues W136 and R176 were important for bile salt binding18, we made and tested the binding properties of two additional mutants, W136A, and R176E. We also tested MtrR W136L, and a Q133A substitution because of the latter’s observed side chain flexibility that allowed the carboxamide group to interact with both testosterone and progesterone (Fig S6A). Of all mutations, D171A affected MtrR binding most significantly resulting in a 2.9-fold increase in Kd for progesterone binding, a 7.0-fold decrease in testosterone affinity, and no observable binding to β-estradiol or ethinyl estradiol (Fig. 4, S6A, B). Thus, this residue is important for ligand specificity, which is further buttressed by the finding that neither cholesterol nor cortisol, which do not have a polar atom in a similar position, bind to MtrR (Fig S1). The MtrR W136A mutant bound β-estradiol and testosterone similar to wild type but did not bind progesterone. We have posited that progesterone may take on multiple binding modes in the ligand binding pocket of MtrR, based on our crystallographic data. This may be the reason why this W136A mutant tolerates binding of testosterone, β-estradiol or ethinyl estradiol, but not progesterone. Similarly, the Kd of MtrR W136L to progesterone is increased by 40-fold compared to WT but binds to β-estradiol and testosterone with comparable affinity to WT MtrR (Fig S6). MtrR R176E binds testosterone and β-estradiol with similar affinity to WT MtrR but binds progesterone with an increased Kd of ~3 fold (Fig S6). MtrR Q133A bound all ligands with Kd values comparable to WT MtrR. Intriguingly, MtrR Q133A bound to progesterone in an endothermic reaction, unlike all other binding events, which are exothermic (Fig S6).

Isothermal titration calorimetry thermograms and resulting binding isotherms for binding reactions between MtrR W136A, R176E, or D171A with progesterone, β-estradiol, and testosterone.
Hormonal effects on gonococcal cells
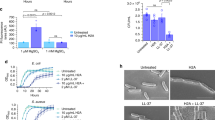
Based on the capacity of the above-described ligands to bind MtrR and abrogate the formation of a complex with the mtrCDE operator sequence we tested whether sub-lethal levels of selected hormones could induce gonococcal resistance to antimicrobials in an MtrR-dependent fashion. First, we determined the minimum inhibitory concentration (MIC) of the steroidal hormones in the gonococcal strain FA19 (Table S1) and a hormone concentration range that does not inhibit the gonococcal growth rate in liquid culture (Fig S7). We found that the MICs for PTR, EST, and TES are 127, >1175 and 555 µM respectively. Deletion of mtrD encoding the inner membrane component of the efflux pump lowered the MIC for PTR, EST, and TES, which indicated that these gonadal hormones are substrates of the MtrCDE pump (Table S1). This is in accord with a previous study that reported the decreased MIC for PTR in the same ∆mtrD strain38. In this regard, a concentration of up to 50 µM for each hormone did not retard the growth rate when gonococci were grown in GC broth (Fig S7). Thus, we found that in the presence of sub-lethal levels of PTR and EST wild-type strain FA19 could express elevated levels of resistance to azithromycin, a known substrate for the MtrCDE efflux pump38, while a genetic derivative lacking MtrR did not show change in antibiotic resistance (Fig. 5A). Intriguingly, we show that MtrR does not bind azithromycin directly (Fig S1), suggesting an alternative mode of action by which MtrR induction contributes to azithromycin resistance. One possibility under investigation is that antibiotics can promote formation of reactive oxygen species that contribute to cellular death47,48. As mentioned, MtrR also represses directly the expression of rpoH, which encodes an oxidative stress response sigma factor that is critical in the defense against reactive oxygen species (ROS) present at gonococcal infection sites21,22,23. Thus, MtrR may be sensing ROS produced as a consequence of antibiotic treatment to induce the efflux pump. Similarly, when these strains were exposed to EST, strain FA19 displayed increased resistance to the model cationic antimicrobial peptide polymyxin B (Fig. 5B).

Cultures (5 × 105 CFU) and 1:5 serial dilutions of wild type FA19 and mutant strain JF1 (ΔmtrR) were spotted on GC agar plates containing progesterone or β-estradiol and azithromycin A or polymyxin B B. Antimicrobial susceptibility was assessed visually at 24 h.
To ascertain if the elevated antimicrobial resistance (AMR) expressed by strain FA19 in the presence of sub-lethal levels of EST and PTR was due to increased expression of mtrCDE we examined the in vivo expression of mtrC and an additional MtrR-repressed gene (rpoH). For this purpose, we used strains producing a wild-type MtrR, MtrR-negative strains and genetic complementation strains expressing wild-type MtrR and mutant MtrR D171A from a copy of the mtrR promoter and translational signals located in trans in the chromosome. We found that in the presence of increasing levels of EST, PTR, and TES expression of mtrC and rpoH could be induced in the complemented (MtrR + ) strains but this induction of gene expression was abrogated due to loss of MtrR or to the production of the MtrR-D171A mutant protein (Fig. 6, S8). Importantly, genes not subjected to MtrR regulation (lptA and ngo1249) were not induced by these ligands (Fig S9). In addition to the MtrR-D171A variant, similar results were obtained with FA19 strains producing the MtrR mutant proteins with single amino acid changes at positions 136 or 176 when grown in the presence of TES, EST or PTR (Fig S10). We observed less induction by the MtrR R176E mutant compared to the MtrR W136L (Fig S10). While MtrR W136L shows almost comparable levels of activation to MtrR WT in the presence of β-estradiol (for either mtrC and rpoH) the R176E mutant shows a clearly weaker activation with β-estradiol for both genes (Fig S10). In the presence of progesterone, both mutants show almost no activation for either mtrC or rpoH, consistent with weak binding reported with ITC (Fig. 4, S6). In the presence of testosterone, MtrR W136L shows a weaker activation for mtrC and comparable activation to the MtrR WT for rpoH, while MtrR R176E shows significantly less activation for both genes (Fig S10). Lastly, we found that ethinyl estrogen could induce expression of mtrC and rpoH but only when the exposed strain produced a wild-type MtrR (Fig S11).

108 CFU/mL of wild-type (WT) mtrR bearing strains (JC106 and JC108), mtrR deletion mutants (JC107 and JC109) and complemented strains in trans with WT mtrR (JC89 and JC91) and mutant allele mtrR D171A (JC101 and JC102) were grown in GC-broth to late exponential phase in increasing concentrations of progesterone (PTR, A, C) or β-estradiol (EST, B, D), statically in 96-well plates (37 °C, 5% v/v CO2). Expression of mtrC (A, B) and rpoH (C, D) was measured from transcriptional and translational lacZ fusions and expressed as a corresponding β-galactosidase activity in Miller units (MU) as described in the Methods. Data are represented by the mean (bar) + SEM (error bar) of n = 4 biological samples in the bar graphs and as fold change in MU relative to the zero-hormone control in the line graphs. MU values are annotated within each bar. The legends shown for the bar and line graphs in A apply to all graphs in C, D. The experiments were performed at least thrice with reproducible results. Statistics: ANOVA test and a Dunnett’s Multiple Comparison post-test (*statistically different from 0 µM at p < 0.05).
Discussion
N. gonorrhoeae is a human pathogen and the etiological agent of the sexually transmitted infection gonorrhea. Many globally isolated strains of N. gonorrhoeae are now multidrug-resistant, and thus has been identified as an urgent public health threat by the CDC7. Understanding the mechanism(s) of MDR in pathogenic bacteria such as N. gonorrhoeae is paramount to combating the prospect of untreatable infections. The overexpression of multidrug efflux pumps that can export myriad antibiotics and other cytotoxic molecules from the cell is an important mechanism for N. gonorrhoeae survival and its development of MDR. By tightly regulating the expression of these efflux pumps, the bacteria can respond quickly to drug treatment or attacks by the host whilst minimizing the energetic cost of efflux pump biosynthesis. Here, we identify the gonadal steroid hormones testosterone, β-estradiol, and progesterone, as well as ethinyl estrogen, a component of some hormonal birth control pills, as physiologically relevant inducers of the MtrCDE efflux and RpoH stress response systems through their binding to MtrR. Although the serum concentrations of these gonadal steroids is in the nM range49,50, the concentrations of progesterone and β-estradiol are significanly higher, low µM range, in the female reproductive tract at certain stages of menses27. Thus, it is likely that the local concentration of testosterone is similiarly higher at the sites of infection where production of these steroids occurs. Previous in vitro work has shown that these steroids arrest gonococcal growth39. During gonococcal colonization of the urogenital tract, this bacterium will encounter potentially toxic local concentrations of these steroidal hormones but which induces the MtrCDE and RpoH systems to allow its survival by efflux and upregulation of stress response genes.
This interbacterial communication between the host-produced hormones and bacterial transcriptional regulators is consistent with other systems34. For example, the locus of enterocyte effacement (LEE) pathogenicity island in Enterohemorrhagic Escherichia coli can be activated by the mammalian hormone epinephrine29. The presence of estradiol significantly increases the ability of Chlamydia trachomatis to adhere to HeLa cells32, and progesterone increases susceptibility to infection in female rat models33. β-estradiol replacement in gonadectomized female and male rat models was reported to have protective effects against Vibrio vulnificus infection31. In prostate cell models, uropathogenic E. coli infection was surpressed by testosterone51.
Interestingly, an early report by James and Swanson revealed phenotypic differences between gonococcal strains that were isolated from human female and male patients, as well as from strains isolated from women at different times during the menstrual cycles52. They showed that strains isolated from female patients tended to have higher resistance to trypsin growth inhibition52. Trypsin susceptibility greatly depended on the phase of the menstrual cycle when isolates were collected, with lowest resistance occuring during the luteal phase when the concentration of progesterone is hightest52. This study highlights the effects of different hosts environments on Neisserial biology52. Hormonal changes during the menstrual cycle influences the protective immune response in the female reproductive tract to optimize conditions for defending against pathogen infection and also repressing immune reponses whilst remodeling tissue to enable implantation and pregnancy26,27. This creates a “window of vulnerability” wherein pathogens can enter and infect the reproductive tract when immune protection is dampened in order to optimize pregnancy26,27. This is consistent with our findings that these gonadal steroids induce stress responses in gonococcal cells that are invading and colonizing a highly dynamic environment.
The structures of MtrR bound to these steroids reveal the ligand-recognition and induction mechanisms and structural consequences of ligand binding. These structures elucidate the highly hydrophobic/aromatic binding pocket of MtrR and the presence of a key aspartate residue that accommodates steroid binding. Site directed mutagenesis of selected residues within the ligand binding pocket reveal varying effects on binding. MtrR D171 is perhaps the most critical contributor to hormonal steroid binding. Additionally, the interaction between D171 and a polar atom of each steroid appears to be a key determinant in discriminating against other steroids such as cholesterol and cortisol, which lack an appropriate proximal polar group. Such a binding pocket is consistent with other multiligand binding proteins such as QacR from S. aureus35, TtgR from Pseudomonas putida53, CmeR from Campylobacter jejuni54and AcrR from E. coli55, the drug-binding sites of which consist mostly of aliphatic and aromatic residues with only a few, but essential, polar residues.
The structures of the MtrR-steroids complexes represent the induced form of MtrR as the center-to-center distance between the recognition helices of the helix-turn-helix DNA binding motifs is not suitable for specific recognition of the nucleobases of two consecutive major grooves when compared to the DNA bound conformation of MtrR25. Such an increase in this distance is consistent with other induction mechanisms of TetR family members35,56,57,58,59. Although members of the TetR family show strong structural homology in their DNA binding domains, the ligand binding pockets vary in composition and volume. For example, the ligand binding pocket of QacR, is unusual for a multiligand binding protein as it utilizes glutamates that become available upon ligand binding due to a conformational change, as well as polar residues to facilitate the binding of multiple drugs, but maintains critical aromatic interactions with each compound35. Binding of bile salts to RamR is facilitated by four hydrogen bonding residues, whilst relying only on π-π stacking to bind compounds such as dequalinium, ethidium, and rhodamine 6G44,60. Here, we show that MtrR utilizes only one polar residue, D171, to bind specifically to gonadal steroids. Like QacR, MtrR recruits a key surface-exposed residue, W136, upon ligand binding thereby allowing the indole ring to cap the pocket35.
In conclusion, we have elucidated the binding and induction mechanisms of hormonal steroids by MtrR, a transcriptional regulator that is a key player in the multidrug resistance arsenal of Neisseria gonorrhoeae. Significantly, the induction of the mtrCDE efflux genes and the gene encoding the rpoH stress response sigma factor by ethinyl estrogen suggests that steroid-based hormonal contraceptives might be co-opted by Neisseria gonorrhoeae to help colonize its human host, hence underscoring that, although these medicines are highly effective in preventing unwanted pregnancies, they do not protect against gonococcal infection.
Methods
Protein overexpression and purification
The gene encoding mtrR is codon optimized (GenScript) and synthesized for expression in E. coli. The gene was subsequently subcloned into the ampicillin resistant pMCSG7 vector in-frame with a T7 promotor, and contains an N-terminal Hexa-histidine tag and a tobacco etch virus (TEV) protease cleavage site. Rosetta Gami B(DE3) pLysS cells were transformed by the plasmid for protein expression. The cells were grown in Luria-Bertani broth containing ampicillin, chloramphenicol, and tetracycline at 37 °C. Cultures were grown to an optical density (OD600) of 0.5–0.6 and expression of MtrR was induced by the addition of 1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) and grown for an additional 3 h before harvesting. Cells were centrifuged, reconstituted in buffer containing 20 mM Tris-HCl pH 8.0, 200 mM NaCl, 10% glycerol, and 1 mM tris-2-carboxyethyl phosphine hydrochloride (TCEP) and lysed by sonication. MtrR was purified from clarified lysate by Ni2+-nitrilotriacetic acid (Ni-NTA) affinity chromatography followed by cleavage of the Hexa-histidine tag by TEV digestion. MtrR was further purified by size exclusion chromatography (S200 column). Selenomethionine modification of MtrR was achieved using the methionine-inhibitory pathway and purified as above61.
Crystallization and data collection
Crystals were grown at 25 °C using the hanging drop-vapor diffusion method. MtrR was co-crystalized with 2.5 mM progesterone, β-estradiol, testosterone, or ethinyl estradiol with 5% MeOH. Crystals formed within two weeks in crystallization solutions containing 1200 mM sodium phosphate monobasic, 800 mM potassium phosphate dibasic, 100 mM CAPS/ NaOH pH 10.5, and 200 mM lithium sulfate. Crystals were cryoprotected in this buffer with 25% glycerol added for flash freezing in liquid nitrogen. Density data were collected at the Advance Light Source (ALS) on beamline 5.0.2 (MtrR-EST and MtrR-TES), 5.0.1 (for MtrR-PTR), or 5.0.3 (for MtrR-NDR). The wavelengths for data collection were 0.9763 (MtrR-EST), 0.9786 (MtrR-TES), 0.9774 (MtrR-PTR), and 0.9765 (MtrR-NDR). Data were collected at 100 K. Crystals of selenomethionine-substituted MtrR in complex with β-estradiol were used for SAD data collection. Phenix Autosol (using Phenix version 1.19)62,63 was then employed to locate and refine Se heavy atom sites, determine initial phases, and carry out density modification. This produced an excellent experimental map, in which 76% of the structure was autotraced. Strong density for β-estradiol was observed in this map. The initial model was subjected to refinement in Phenix-refine and multiple cycles of rebuilding and refinement62. After the final refinement, a dimer of this structure was used in molecular replacement to solve the structures of MtrR bound to progesterone, testosterone, and ethinyl estrogen. Iterative cycles of the model building were done in COOT (version 0.9.6)64 and refinement using Phenix refine62,65,66. The stereochemistry was excellent with the favored percent Ramachandran for each structure: 98.82 (MtrR-EST), 98.02 (MtrR-TES), 98.25 (MtrR-PTR), and 87.86 (MtrR-NDR.) There were no Ramachandran outliers in any structure except for MtrR bound to NDR which had the lowest resolution. Clash scores for all structures were less than or equal to 6 except MtrR bound to NDR, which had a clashscore of 10.83. Mean I/sigma(I) values for the highest resolution shell were 0.7 (MtrR-EST), 2.2 (MtrR-TES), and 2.3 (for both MtrR-PTR and MtrR-NDR). Selected data collection and refinement statistics are listed in Table 1. Alignment analysis and protein visualization were done in Pymol (version 2.5.4).
Isothermal titration calorimetry
Purified MtrR was concentrated to 20 µM in purification buffer plus 1% MeOH. The addition of 1% methanol is necessary to dissolve each hormonal steroid, cortisol, and cholesterol in the purification buffer. Titrations with 20 or 5 µM MtrR in the sample cell and 125, 200, or 500 µM ligand in the syringe were performed using a VP-ITC microcalorimeter (Microcal Inc.). ITC experiments were conducted at 25 °C with a stirring speed of 199 rpm. Spacing between injections was 600 or 300 s (no difference in results found). Germane blanks were subtracted, and the data analyzed using ORIGIN 7.067. At least three experimental measurements, technical and some biological replicates, were averaged for each reported binding constant.
Fluorescence polarization based-DNA binding assays
Fluorescence polarization based-DNA binding data were collected using a Panvera Beacon 2000 fluorescence polarization system (Invitrogen) and analyzed with Prism (Graphpad Software 9, 10). Purified MtrR was buffer exchanged into “DNA” binding buffer (150 mM NaCl, 20 mM Tris-HCl pH 7.5, 2.5% glycerol, 1 mM TCEP). MtrR was titrated into a binding buffer solution that contained 1 nM 5’-fluoresceine-labeled DNA (containing either the rpoH or mtrCDE operator site), 1 µg of poly(dI-dC) DNA, 1 µg bovine serum albumin (BSA) and 125 µM progesterone, testosterone, or β-estradiol (or ethinyl estradiol, see Fig. S2) resulting in a final methanol concentration of 1%. The reported oligoduplexes containing the rpoH and mtrCDE binding site differ in lengths, 27mer and 21mer, respectively17,21. Previous work from our laboratories has shown that these differences in length have no impact on binding affinity between the two sites25. BSA and dI-dC are included as carriers and to control for nonspecific DNA binding. Samples were excited at 490 nm and polarized emissions measured at 530 nm. Controls were run with no protein (DNA \(\pm\) methanol, hormonal steroids) to ensure that millipolarization values were not affected by experimental conditions alone. The binding affinity of protein for DNA was determined using the following equation: P = Pf + (Pb – Pf) [M] / (Kd + [M]) where P is the measured polarization, Pf is the polarization of free DNA, Pb is the polarization of the completely bound DNA, Kd is the equilibrium dissociation constant, and M is the concentration of protein. Pb and Kd were determined by nonlinear least-squares analysis. At least three independent experimental measurements were averaged for each reported binding constant.
Strains and media
N. gonorrhoeae strains used in this study are described in Table S2. Gonococcal cultures were grown overnight at 37 °C in a 5% (v/v) CO2 incubator on GC agar (BD Difco) plates supplemented with Kellogg’s supplements I and II68. Growth in liquid medium was at 37 °C in GC broth (Proteose Peptone No.3 15 g/L, dipotassium phosphate 4 g/L, monopotassium phosphate 1 g/L, sodium chloride 5 g/L) containing Kellogg’s supplements I and II and 0.042% (w/v) sodium bicarbonate. E. coli TOP10 cells (Life Technologies, Carlsbad, CA) were used for cloning steps and grown on LB medium69.
Gonococcal genetic transformations
Plasmids and oligonucleotide primers used for gonococcal genetic transformations are described in Table S2 and S3 respectively. Genetic transformation of N. gonorrhoeae was achieved by homologous recombination using the spot agar transformation method described perviously70. A complete description of cloning and transformation steps of derivatives of gonococcal strain FA19 is described in the supplementary material.
Growth of gonococcal strains in 96-well plates and β-galactosidase assays
To determine the effect of the steroidal hormones on the expression of MtrR-regulated genes, gonococcal strains bearing the transcriptional and translational lacZ fusion constructs were grown in GC-broth in 96-well plates (Costar #3370 flat bottom, sterile, with lid). β-galactosidase activities were determined by hydrolysis of the colorimetric substrate ortho-Nitrophenyl-β-galactoside (ONPG) using a modified version of the J.H Miller71 protocol as described in the supplementary material.
MIC and antimicrobial susceptibility testing
To determine the minimal inhibitory concentrations (MICs) by the steroidal hormones and their effect on antimicrobial susceptibility, 5 x 105 CFU and 1:5 serial dilutions of gonococcal strains were spotted on GC agar plates containing different concentrations of antibiotics and/or hormones as indicated. Antimicrobial susceptibility was assessed after 24 h incubation.
Site directed mutagenesis
The gene encoding the MtrR protein was codon optimized and synthesized for expression in E. coli (GenScript). The gene was subsequently subcloned into the ampicillin resistant pMCSG7 vector in-frame with a T7 promotor, N-terminal Hexa-histidine tag, and tobacco etch virus (TEV) protease cleavage site. This was used as the template for site directed mutagenesis. Site directed mutagenesis was performed with complementary oligonucleotides containing the desired mutations, obtained from Integrated DNA Technologies (IDT), as previously descrIbed18. Protein was expressed and purified as described in Materials and Methods.
Plate-based fluorescence polarization DNA-binding assay
To analyze the effect of ethinyl estradiol on DNA binding by MtrR, fluorescence polarization DNA-binding data were collected in 96-well plates (Costar #3915 black, flat bottom). with a SpectraMax M5e Microplate Reader (Molecular Devices) and analyzed with Prism (Graphpad Software, version 10.0.0). Purified MtrR was buffer exchanged into “DNA” binding buffer (150 mM NaCl, 20 mM Tris-HCl pH 7.5, 2.5% glycerol, 1 mM TCEP). MtrR was titrated into a binding buffer solution that contained 10 nM 5’-fluoresceine-labeled DNA (encompassing either the rpoH or mtrCDE operator site), 1 µg of poly(dI-dC) DNA, 12 µg bovine serum albumin (BSA) and 125 µM ethinyl estradiol resulting in the final methanol concentration of 1%. The binding affinity of protein for DNA was determined using the Morrison’s quadratic equation72. BSA and dI-dC are included as carriers and to control for nonspecific DNA binding. Samples were excited at 485 nm and polarized emissions measured at 525 nm, with a 515 nm cutoff. Controls were run with no protein (DNA \(\pm\) methanol, ethinyl estradiol) to ensure that millipolarization values were not affected by experimental conditions alone. At least three independent experimental measurements were averaged for each reported binding constant.
Gonococcal genetic transformations
Plasmids and oligonucleotide primers used are described in Tables S2 and S3, respectively. To study the hormone effect on the expression of MtrR-regulated genes mtrC and rpoH, lacZ transcriptional and translational fusions were constructed using pLES9473 and integrated into the proAB locus of N. gonorrhoeae strains. Genes lptA and ngo1249, not present within the MtrR regulon, were used as negative controls. To obtain the mtrC-, rpoH- and ngo1249-lacZ fusions the promoter region and the initial 3 to 6 codons of the genes were amplified from FA19 gDNA using the primer pairs mtrC-lacZ-F/mtrC-lacZ-R, rpoH-lacZ-F/rpoH-lacZ-R and 1249-lacZ-F/1249-lacZ-R respectively. The lptA-lacZ fusion was constructed before74. The resulting PCR products were digested with BamHI, cloned into similarly digested pLES94 and transformed into TOP10 cells. Blue colonies were selected in LB agar containing chloramphenicol (10 µg/mL) and X-gal (20 µg/mL). The resulting recombinant plasmids were sequenced with primers proABFw and lacZRv. Plasmids bearing the expected sequences were linearized with HindIII and transformed into N. gonorrhoeae strains (Table S2) using a spot agar transformation method70 and selection on GC-agar containing chloramphenicol (0.5 µg/mL). Integration of the lacZ fusion at the proAB locus was confirmed by PCR with primers proABFw and lacZRv and by Sanger sequencing.
To complement mtrR expression in mtrR null mutant strain JF1, the promoter region and coding sequence (CDS) of mtrR was amplified from FA19 gDNA using primers pacImtrR-Fw and pmeImtrR-Rv. The PCR fragment was double digested with PacI and PmeI and ligated into similarly digested pGCC375. The resulting pGCC3-mtrR was sequenced with pGCC3Rv and mtrR-qRT-R and used to transform strains JC107 and JC109 to obtain strains JC89 and JC91, respectively. Selection of the transformant strains was carried out in GC-agar containing erythromycin (1.0 µg/mL). Correct integration of the plasmids into the lctP-aspC loci was confirmed by PCR with primers lctPqFw and pGCC3Rv and sequencing of the PCR products with pGCC3Rv and mtrR-qRT-R.
To complement mtrR expression with mutant alleles encoding the missense mutations in MtrR W136L, R176E, or D171A overlap extension PCRs were prepared to fuse a WT mtrR promoter region with mutant mtrR alleles. Briefly, a PCR fragment encoding the promoter region was obtained with primers pacImtrR-Fw and mtrR-midRv and FA19 gDNA; while the mutant alleles were amplified from site-directed mutagenesis created-vectors pGAB020RGB (mtrR W136L), pGAB018RGB (mtrR R176E) and pmtrRD171A (mtrR D171A) using primers mtrR-mid-Fw and PmeI-mtrR-Rv. The two resulting PCR fragments (promoter + mutant mtrR) were mixed and used as templates for the amplification of the fused promoter region-mutant mtrR alleles fragments with primers pacImtrR-Fw and PmeI-mtrR-Rv. These final PCR fragments were digested with PacI and PmeI and ligated into similarly digested pGCC3. Complementation vectors expressing the mutant mtrR alleles from their own transcriptional and translational signals were transformed into strains JC107 and JC109 as described above for pGCC3-mtrR.
Growth of gonococcal strains in 96-well plates and β-galactosidase assays
To determine the effect of the steroidal hormones on the expression of MtrR-regulated genes, gonococcal strains bearing the lacZ fusion constructs were grown in GC-broth in 96-well plates (Costar #3370 flat bottom, sterile, with lid). Cultures (100 µL) were adjusted to an initial optical density at 600 nm (OD600) of 0.1 containing serial dilutions of the hormones as indicated and were incubated statically at 37 °C in the 5% (v/v) CO2 incubator for 7 h until late-exponential growth phase. Stock solutions of the hormones were prepared to 3.3 mM in a 2:1 mix of methanol:H2O. The OD600 of the plates was determined in a plate reader and the plates were kept in ice until use. Next, cell lysis was achieved by resuspending 18.75 µL of the cultures in 56.25 µL of CelLytic B Cell lysis reagent (Sigma) in a new 96-well plate and incubating 30 min at 26 °C. Beta-galactosidase reactions started by adding 100 µL of a reaction mix containing 62.5 µL of buffer Z (60 mM Na2HPO4, 40 mM NaH2PO4 pH 7.0, 10 mM KCl, 1 mM MgSO4) and 37.5 µL of 4 mg/mL ortho-Nitrophenyl-β-galactoside (ONPG) (diluted in 0.1 M phosphate buffer pH 7.0). Reactions were incubated at 26 °C until a pale-yellow color developed, were stopped with 73 µL of 1 M Na2CO3 and the OD420 was read with a plate reader. A modified formula was used to calculate the Miller units71 of the cultures: Miller units = ((OD420 nm - OD420 nm blank) × 1.4925 × 1000)/(t(min) × V(mL) × 6.857 × OD600nm × 5.5897), where 1.4925 is the conversion factor to correct the OD420 obtained with the light path of a 248 µL volume in 96-well plates to the light path using cuvettes (1 cm) in a spectrophotometer, t is the incubation time of the assay in min, V is the volume of cells used for the assay (18.75 µL), 6.857 is to convert the volume of cells used in the microplate assay to the equivalent volume used in the original Miller protocol, OD600nm is the optical density of the cells after growth, and 5.5897 is the conversion factor to transform the OD600 of the cultures in 96-well plates (100 µL) to the OD600nm obtained using a spectrophotometer and a cuvette with a light path of 1 cm. In this formula the turbidity correction (1.75 × OD550nm) used in the original Miller formula was omitted due to the low turbidity of the reactions.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data in this study are included in the article. The coordinates and structure factors for the MtrR-PTR, MtrR-EST, MtrR-TES, and MtrR-NDR complexes have been deposited in the RCSB Protein Data Bank with the accession codes 8FW8, 8FW0, 8FW3, and 8SSH, respectively. Source data are provided in this paper.
References
Unemo, M. et al. Gonorrhoea. Nat. Rev. Dis. Prim. 5, 79 (2019).
Rowley, J. et al. Chlamydia, gonorrhoea, trichomoniasis and syphilis: global prevalence and incidence estimates, 2016. Bull. World Health Organ 97, 548–562 (2019).
Kreisel, K. M. et al. Sexually transmitted infections among US women and men: prevalence and incidence estimates, 2018. Sex. Transm. Dis. 48, 208–214 (2021).
Rice, P. A., Shafer, W. M., Ram, S. & Jerse, A. E. Neisseria gonorrhoeae: drug resistance, mouse models, and vaccine development. Annu. Rev. Microbiol. 71, 665–686 (2017).
Quillin, S. J. & Seifert, H. S. Neisseria gonorrhoeae host adaptation and pathogenesis. Nat. Rev. Microbiol. 16, 226–240 (2018).
Unemo, M. & Shafer, W. M. Antimicrobial resistance in Neisseria gonorrhoeae in the 21st century: past, evolution, and future. Clin. Microbiol. Rev. 27, 587–613 (2014).
CDC. Antibiotic Resistance Threats in the United States 2019. Centers for Disease Control and Prevention Available at https://stacks.cdc.gov/view/cdc/82532 (Accessed 1 January 2020).
Wi, T. et al. Antimicrobial resistance in Neisseria gonorrhoeae: global surveillance and a call for international collaborative action. PLoS Med. 14, e1002344 (2017).
de Curraize, C. et al. Ceftriaxone-resistant Neisseria gonorrhoeae Isolates (2010 to 2014) in France characterized by using whole-genome sequencing. Antimicrob. Agents Chemother. 60, 6962–6964 (2016).
Kueakulpattana, N. et al. Multidrug-resistant Neisseria gonorrhoeae infection in heterosexual men with reduced susceptibility to ceftriaxone, first report in Thailand. Sci. Rep. 11, 21659 (2021).
Lee, K. et al. Clonal expansion and spread of the ceftriaxone-resistant Neisseria gonorrhoeae strain FC428, identified in Japan in 2015, and closely related isolates. J. Antimicrob. Chemother. 74, 1812–1819 (2019).
Terkelsen, D. et al. Multidrug-resistant Neisseria gonorrhoeae infection with ceftriaxone resistance and intermediate resistance to azithromycin, Denmark, 2017. Euro Surveill 22 https://doi.org/10.2807/1560-7917.Es.2017.22.42.17-00659 (2017).
Elwell, L. P., Roberts, M., Mayer, L. W. & Falkow, S. Plasmid-mediated beta-lactamase production in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 11, 528–533 (1977).
Morse, S. A., Johnson, S. R., Biddle, J. W. & Roberts, M. C. High-level tetracycline resistance in Neisseria gonorrhoeae is result of acquisition of streptococcal tetM determinant. Antimicrob. Agents Chemother. 30, 664–670 (1986).
Olesky, M., Zhao, S., Rosenberg, R. L. & Nicholas, R. A. Porin-mediated antibiotic resistance in Neisseria gonorrhoeae: ion, solute, and antibiotic permeation through PIB proteins with penB mutations. J. Bacteriol. 188, 2300–2308 (2006).
Piddock, L. J. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 19, 382–402 (2006).
Lucas, C. E., Balthazar, J. T., Hagman, K. E. & Shafer, W. M. The MtrR repressor binds the DNA sequence between the mtrR and mtrC genes of Neisseria gonorrhoeae. J. Bacteriol. 179, 4123–4128 (1997).
Beggs, G. A. et al. Structural, biochemical, and in vivo characterization of MTRR-mediated resistance to innate antimicrobials by the human pathogen Neisseria gonorrhoeae. J. Bacteriol. 201. https://doi.org/10.1128/JB.00401-19 (2019).
Shafer, W. M., Qu, X., Waring, A. J. & Lehrer, R. I. Modulation of Neisseria gonorrhoeae susceptibility to vertebrate antibacterial peptides due to a member of the resistance/nodulation/division efflux pump family. Proc. Natl Acad. Sci. USA 95, 1829–1833 (1998).
Nikaido, H. Multidrug resistance in bacteria. Annu Rev. Biochem 78, 119–146 (2009).
Folster, J. P. et al. MtrR modulates rpoH expression and levels of antimicrobial resistance in Neisseria gonorrhoeae. J. Bacteriol. 191, 287–297 (2009).
Gunesekere, I. C. et al. Comparison of the RpoH-dependent regulon and general stress response in Neisseria gonorrhoeae. J. Bacteriol. 188, 4769–4776 (2006).
Laskos, L., Ryan, C. S., Fyfe, J. A. & Davies, J. K. The RpoH-mediated stress response in Neisseria gonorrhoeae is regulated at the level of activity. J. Bacteriol. 186, 8443–8452 (2004).
Chen, S. C., Yin, Y. P., Dai, X. Q., Unemo, M. & Chen, X. S. Antimicrobial resistance, genetic resistance determinants for ceftriaxone and molecular epidemiology of Neisseria gonorrhoeae isolates in Nanjing, China. J. Antimicrob. Chemother. 69, 2959–2965 (2014).
Beggs, G. A. et al. Structures of Neisseria gonorrhoeae MtrR-operator complexes reveal molecular mechanisms of DNA recognition and antibiotic resistance-conferring clinical mutations. Nucleic Acids Res. 49, 4155–4170 (2021).
Collins, M. K., McCutcheon, C. R. & Petroff, M. G. Impact of estrogen and progesterone on immune cells and host-pathogen interactions in the lower female reproductive tract. J. Immunol. 209, 1437–1449 (2022).
Wira, C. R., Rodriguez-Garcia, M. & Patel, M. V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 15, 217–230 (2015).
Kwon, M. S. & Lee, H. K. Host and microbiome interplay shapes the vaginal microenvironment. Front Immunol. 13, 919728 (2022).
Sperandio, V., Torres, A. G., Jarvis, B., Nataro, J. P. & Kaper, J. B. Bacteria-host communication: the language of hormones. Proc. Natl Acad. Sci. USA 100, 8951–8956 (2003).
Rumbaugh, K. P. Convergence of hormones and autoinducers at the host/pathogen interface. Anal. Bioanal. Chem. 387, 425–435 (2007).
Merkel, S. M., Alexander, S., Zufall, E., Oliver, J. D. & Huet-Hudson, Y. M. Essential role for estrogen in protection against Vibrio vulnificus-induced endotoxic shock. Infect. Immun. 69, 6119–6122 (2001).
Bose, S. K. & Goswami, P. C. Enhancement of adherence and growth of Chlamydia trachomatis by estrogen treatment of HeLa cells. Infect. Immun. 53, 646–650 (1986).
Kaushic, C., Zhou, F., Murdin, A. D. & Wira, C. R. Effects of estradiol and progesterone on susceptibility and early immune responses to Chlamydia trachomatis infection in the female reproductive tract. Infect. Immun. 68, 4207–4216 (2000).
Hughes, D. T. & Sperandio, V. Inter-kingdom signalling: communication between bacteria and their hosts. Nat. Rev. Microbiol. 6, 111–120 (2008).
Schumacher, M. A. et al. Structural mechanisms of QacR induction and multidrug recognition. Science 294, 2158–2163 (2001).
Kisker, C., Hinrichs, W., Tovar, K., Hillen, W. & Saenger, W. The complex formed between Tet repressor and tetracycline-Mg2+ reveals mechanism of antibiotic resistance. J. Mol. Biol. 247, 260–280 (1995).
Lin, J. et al. Bile salts modulate expression of the CmeABC multidrug efflux pump in Campylobacter jejuni. J. Bacteriol. 187, 7417–7424 (2005).
Chitsaz, M., Booth, L., Blyth, M. T., O’Mara, M. L. & Brown, M. H. Multidrug Resistance in Neisseria gonorrhoeae: identification of functionally important residues in the MtrD Efflux Protein. mBio 10. https://doi.org/10.1128/mBio.02277-19 (2019)
Fitzgerald, T. J. & Morse, S. A. Alteration of growth, infectivity, and viability of Neisseria gonorrhoeae by gonadal steroids. Can. J. Microbiol. 22, 286–294 (1976).
Jerse, A. E. et al. A gonococcal efflux pump system enhances bacterial survival in a female mouse model of genital tract infection. Infect. Immun. 71, 5576–5582 (2003).
Kita, E., Matsuura, H. & Kashiba, S. A mouse model for the study of gonococcal genital infection. J. Infect. Dis. 143, 67–70 (1981).
Kita, E., Takahashi, S., Yasui, K. & Kashiba, S. Effect of estrogen (17 beta-estradiol) on the susceptibility of mice to disseminated gonococcal infection. Infect. Immun. 49, 238–243 (1985).
CDC. U.S. Selected Practice Recommendations for Contraceptive Use, 2013. adapted from the World Health Organization selected practice recommendations for contraceptive use, 2nd edition. MMWR Recomm. Rep. 62, 1–60 (2013).
Yamasaki, S. et al. Crystal structure of the multidrug resistance regulator RamR complexed with bile acids. Sci. Rep. 9, 177 (2019).
Stourac, J. et al. Caver Web 1.0: identification of tunnels and channels in proteins and analysis of ligand transport. Nucleic Acids Res. 47, W414–w422 (2019).
Veevers, R. & Hayward, S. Methodological improvements for the analysis of domain movements in large biomolecular complexes. Biophys. Physicobiol. 16, 328–336 (2019).
Kohanski, M. A., Dwyer, D. J., Hayete, B., Lawrence, C. A. & Collins, J. J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 130, 797–810 (2007).
Dwyer, D. J. et al. Antibiotics induce redox-related physiological alterations as part of their lethality. Proc. Natl Acad. Sci. USA 111, E2100–E2109 (2014).
Ferri, F. F. Progesterone (serum). (Elsevier, 2019).
J. Larry Jameson, L. J. D. G. Diagnosis and treatment of disorders of sexual development. Vol. 7th Chapter 119 (Elsevier Saunders, 2016).
Ho, C. H. et al. Testosterone regulates the intracellular bacterial community formation of uropathogenic Escherichia coli in prostate cells via STAT3. Int. J. Med. Microbiol. 310, 151450 (2020).
James, J. F. & Swanson, J. Studies on gonococcus infection. XIII. Occurrence of color/opacity colonial variants in clinical cultures. Infect. Immun. 19, 332–340 (1978).
Alguel, Y. et al. Crystal structures of multidrug binding protein TtgR in complex with antibiotics and plant antimicrobials. J. Mol. Biol. 369, 829–840 (2007).
Gu, R. et al. Crystal structure of the transcriptional regulator CmeR from Campylobacter jejuni. J. Mol. Biol. 372, 583–593 (2007).
Li, M. et al. Crystal structure of the transcriptional regulator AcrR from Escherichia coli. J. Mol. Biol. 374, 591–603 (2007).
Orth, P., Schnappinger, D., Hillen, W., Saenger, W. & Hinrichs, W. Structural basis of gene regulation by the tetracycline inducible Tet repressor-operator system. Nat. Struct. Biol. 7, 215–219 (2000).
Yeo, H. K., Park, Y. W. & Lee, J. Y. Structural basis of operator sites recognition and effector binding in the TetR family transcription regulator FadR. Nucleic Acids Res. 45, 4244–4254 (2017).
Le, T. B., Schumacher, M. A., Lawson, D. M., Brennan, R. G. & Buttner, M. J. The crystal structure of the TetR family transcriptional repressor SimR bound to DNA and the role of a flexible N-terminal extension in minor groove binding. Nucleic Acids Res. 39, 9433–9447 (2011).
Frénois, F., Baulard, A. R. & Villeret, V. Insights into mechanisms of induction and ligands recognition in the transcriptional repressor EthR from Mycobacterium tuberculosis. Tuberculosis (Edinb.) 86, 110–114 (2006).
Yamasaki, S. et al. The crystal structure of multidrug-resistance regulator RamR with multiple drugs. Nat. Commun. 4, 2078 (2013).
Doublié, S. Preparation of selenomethionyl proteins for phase determination. Methods Enzymol. 276, 523–530 (1997).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D. Biol. Crystallogr. 66, 213–221 (2010).
Terwilliger, T. C. et al. Decision-making in structure solution using Bayesian estimates of map quality: the PHENIX AutoSol wizard. Acta Crystallogr. D. Biol. Crystallogr. 65, 582–601 (2009).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D. Biol. Crystallogr. 60, 2126–2132 (2004).
Afonine, P. V. et al. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D. Biol. Crystallogr 68, 352–367 (2012).
Williams, C. J. et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 27, 293–315 (2018).
Origin and OriginPro. OriginLab Corporation Available at https://www.originlab.com/index.aspx?go=Products%2FOrigin#Apps_in_Origin (Accessed 18 June 2021).
Kellogg, D. S. et al. Neisseria gonorrhoeae. I. Virulence genetically linked to clonal variation. J. Bacteriol. 85, 1274–1279 (1963).
Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 62, 293–300 (1951).
Dillard, J. P. Genetic Manipulation of Neisseria gonorrhoeae. Current Protocols in Microbiology 23, 4A.2.1-4A2.24. https://doi.org/10.1002/9780471729259.mc04a02s23 (2011).
Miller, J. H. Experiments in molecular genetics. (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1972).
Copeland, R. A. Enezymes. A practical Introduciton to Structure, Mechanism, and Data Analysis. Second edn, 310–315 (John Wiley & Sons, 2000).
Silver, L. E. & Clark, V. L. Construction of a translational lacZ fusion system to study gene regulation in Neisseria gonorrhoeae. Gene 166, 101–104 (1995).
Kandler, J. L. et al. The MisR response regulator is necessary for intrinsic cationic antimicrobial peptide and aminoglycoside resistance in Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 60, 4690–4700 (2016).
Mehr, I. J. & Seifert, H. S. Random shuttle mutagenesis: gonococcal mutants deficient in pilin antigenic variation. Mol. Microbiol. 23, 1121–1131 (1997).
Acknowledgements
We would like to thank the beamline scientists at ALS BL 8.3.1 and BL 5.0.2 for their help with data collection. In addition, we acknowledge the Advanced Light Source at Lawrence Berkeley National Laboratory. National Institutes of Health [R35 GM130290 to M.A.S., R05 AI048593 to R.G.B., R01 AI021150 to W.M.S.]; W.M.S. is the recipient of a Senior Research Career Scientist Award from the Biomedical Laboratory Research and Development Service of the U.S. Department of Veterans Affairs; The Advanced Light Source is supported by the Director, Office of Science, Office of Basic Energy Sciences, Material Sciences Division, of the U.S. Department of Energy [DE-AC03-76SF00098]. Funding for open access charge: Duke University School of Medicine.
Author information
Authors and Affiliations
Contributions
G.M.H., J.C.A., W.M.S., and R.G.B. designed the experiments and analyzed the biochemical and cellular data. G.M.H., R.G.B., and M.A.S. analyzed the structural data. G.M.H. and G.A.B. generated purification constructs. G.M.H. purified proteins, determined their structures, and performed biochemical characterizations. M.A.S. and G.M.H. collected X-ray crystallography data and M.A.S. provided X-ray crystallography consulting and experimental input. G.M.H. J.C.A., W.M.S., and R.G.B. wrote the manuscript. J.C.A., C.L.H., and V.D. conducted cellular experiments. We also thank J.R.P. for insights into steroid and pathogen biology and for providing testosterone for the project. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Peer review
Peer review information
Nature Communications thanks Robert Nicholas and the other, anonymous, reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral concerning jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hooks, G.M., Ayala, J.C., Holley, C.L. et al. Hormonal steroids induce multidrug resistance and stress response genes in Neisseria gonorrhoeae by binding to MtrR. Nat Commun 15, 1153 (2024). https://doi.org/10.1038/s41467-024-45195-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-45195-1
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
