
Abstract
Plants extract mineral nutrients from the soil, or from interactions with mutualistic soil microbes via their root systems. Adapting root architecture to nutrient availability enables efficient resource utilization, particularly in patchy and dynamic environments. Root growth responses to soil nitrogen levels are shoot-mediated, but the identity of shoot-derived mobile signals regulating root growth responses has remained enigmatic. Here we show that a shoot-derived micro RNA, miR2111, systemically steers lateral root initiation and nitrogen responsiveness through its root target TML (TOO MUCH LOVE) in the legume Lotus japonicus, where miR2111 and TML were previously shown to regulate symbiotic infections with nitrogen fixing bacteria. Intriguingly, systemic control of lateral root initiation by miR2111 and TML/HOLT (HOMOLOGUE OF LEGUME TML) was conserved in the nonsymbiotic ruderal Arabidopsis thaliana, which follows a distinct ecological strategy. Thus, the miR2111-TML/HOLT regulon emerges as an essential, conserved factor in adaptive shoot control of root architecture in dicots.
Similar content being viewed by others

Introduction
Root systems are the main contact point of land plants with soluble nutrients. Adapting the root surface area to nutrient availability in the substrate is thus a key aspect of endogenous resource management in land plants.
Consistently, the perception of both restrictive and sufficient levels of nitrate, a frequently limiting macronutrient, induces root architectural adaptations. To this end, plants can enhance lateral root growth, and thus root surface area, either under deficient nitrate conditions or within local, nitrogen-rich patches. This process is termed foraging1. Within a given root segment, several factors have been suggested to be involved in regulating nitrate foraging locally. In Arabidopsis thaliana (Arabidopsis), the nitrate transceptor protein NRT1.1 controls biosynthesis and transport of auxin, thereby mediating local repression of lateral root development where perceived nitrate levels are low2. Downstream of NRT1.1, the GRAS transcription factor NIN LIKE PROTEIN 7 (NLP7) was shown to induce expression of the MADS-box gene ANR1, mediating further transcriptional changes that specifically promote lateral root elongation in nitrate rich soil patches3. Signalling via CLE (CLAVATA3/ESR) peptides and the leucine-rich repeat receptor kinase CLAVATA1 (CLV1) was further shown to be involved in nitrate-dependent local regulation of lateral root emergence in Arabidopsis4. A similar role has been assigned to the putative CLV1-ortholog HYPERNODULATION ABERRANT ROOT FORMATION1 (HAR1) in Lotus japonicus (Lotus)5. Balancing need and availability of nutrients is a challenge concerning the plant as a whole. Adaptations to nutrient stress thus require communication not only within, but also across plant organs, suggesting that they involve systemic signalling circuits linking above- and belowground tissues. Grafting experiments demonstrated that both shoot and root expression of HAR1 is required for nitrate-dependent adaptation of lateral root growth in Lotus5. This suggests a dual root-specific as well as systemic role of the CLE-HAR1 signalling node. Consistently, Lotus CLE-RS peptides were shown to be competent of xylem-based root-shoot mobility following arabinosylation, and can directly bind to HAR16. In addition, both C-terminally encoded peptide (CEP) hormones1,2 as well as cytokinins3,4 act as systemic root-shoot factors signalling low or high root nitrate content, respectively. In Arabidopsis, upon CEP perception by the CEP receptors CEPR1/2, shoot-produced CEP Downstream (CEPD) and CEPD LIKE (CEPDL) peptides translocate to roots to regulate nitrate uptake via transcriptional as well as post-translational regulation of NRT2.17,8. In the legume Medicago truncatula (Medicago), the putative CEPR1 orthologue COMPACT ROOT ARCHITECTURE 2 (CRA2) similarly steers NRT2.1 dependent nitrate uptake by mediating CEP1 dependent expression regulation9,10.
While shoot-root mobile CEPD and CEPDL signals systemically regulate root nitrate uptake, systemic shoot factors mediating nitrate-dependent root growth adaptations are so far unknown. We previously observed that a shoot-derived, phloem-mobile micro RNA, miR2111, regulates the formation of symbiotic infections and nitrogen-fixing nodule organs in Lotus japonicus (Lotus) roots inoculated with rhizobial bacteria11. miR2111 post-transcriptionally targets the root-expressed F-Box Kelch-repeat gene TOO MUCH LOVE (TML), which represses symbiosis11,12. Both CLE-RS/HAR111 and CEP/CRA2 signalling nodes12 regulate miR2111 abundance. Shoot miR2111 accumulation is repressed in the presence of sufficient nitrate as well as of compatible rhizobia11, releasing TML mRNA from posttranscriptional regulation and restricting symbiosis progression. Interestingly, miR2111 and TML are not restricted to plants establishing root nodule symbiosis, but are conserved across dicot lineages. The Arabidopsis genome contains two MIR2111 precursor gene loci13, both encoding a single miR2111 isoform that specifically targets the F-box Kelch-repeat gene At3g2715013, a TML homolog14 of unknown function. In comparison, the Lotus genome contains seven MIR2111 loci encoding three different isoforms11,15. On this basis, we hypothesized that miR2111 may have a conserved role in regulating lateral organ formation in roots also in nonsymbiotic settings, and have undergone functional diversification in nodulating lineages. Our work identifies miR2111 as a missing link signalling shoot nitrogen status to root organs and regulating adaptive root growth responses in a nitrate-dependent manner.
Results and discussion
miR2111 is a shoot factor regulating root architecture
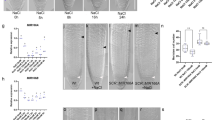
To investigate possible conserved, symbiosis-independent functions of miR2111 in root system architecture control, we analysed root systems of plants mis-expressing the miRNA. Indeed, Lotus plants expressing a pUBQ1::MIR2111-3 transgene resulting in overabundance of mature miR2111 (Fig. 1a) generated less lateral roots than wild type plants (Fig. 1b). miR2111 is produced primarily in shoots, and is proposed to translocate to roots via the phloem11,15. Phloem-mobile miRNAs were recently suggested to translocate as fully processed duplices, rather than as pri- or pre-miRNA precursors16. Consistently, we could trace plant specific mature miR2111 transcripts in aphids (Planococcus citri) feeding on Lotus, indicating its presence in the phloem sap (Supplementary Fig. 1a–c). To investigate whether shoot-derived miR2111 is indeed functional in Lotus roots and sufficient to regulate lateral root number, we grafted pUBQ1::MIR2111-3 expressing shoots onto wild type root stocks (Fig. 1c). Roots of chimeric plants showed enhanced levels of miR2111 (Fig. 1d), and fewer emerged lateral roots compared to control grafts (Fig. 1c, e), confirming that shoot miR2111 indeed translocates to roots to steer lateral root numbers.

a miR2111 abundance fold change compared to Gifu wild type (wt(G)) plants, and (b) LR count in transgenic pUBQ1::MIR2111-3 (2111ox) expressing lines (#3, 4) compared to wt(G). Line #3 was used for further analysis. c–f 2111ox / wt(G) (shoot / root) grafts compared to wt(G) / wt(G) control grafts. c Example of grafted plants. Scale bars equal 1 cm. d miR2111 levels, (e) LR numbers and (f) TML levels in roots of respective grafts. a, d, f qRT-PCR analyses. RNA levels are relative to those of two reference genes. RNA was extracted from root (d, f) or shoot samples (a). a Adult plants grown in soil. b–g Plants were grown at 0 mM nitrate and evaluated or harvested after two weeks of cultivation. Student’s t-test (**p ≤ 0.01; ***p ≤ 0.001) (d–f) or analysis of variance (ANOVA) and post-hoc Tukey testing (p ≤ 0.05) (b), with distinct letters indicating significant differences. All experiments were in ecotype Gifu B-129 (wt(G)). Sample size, replicates and exact p-values are listed in the Source Data file. Dotplots show individual data points and a line indicating their average value. Boxplot central line shows median value, box limits indicate the 25th and 75th percentile. Whiskers extend 1.5 times the interquartile range, or to the last datapoint. Individual datapoints are represented by dots.
miR2111 regulates lateral root initiation through its target TML
miR2111 was proposed to directly target TML for posttranscriptional regulation in Lotus as well as Medicago11,12. Consistently, roots of pUBQ1::MIR2111-3/wild type (shoot/root) grafts had significantly lower TML levels than wild type/wild type controls (Fig. 1f). tml knockout mutants developed less lateral roots than wild type plants (Fig. 2a, b), and were phenotypically indistinguishable from pUBQ1::MIR2111-3 plants (Fig. 2a, b), suggesting that TML is the main target of miR2111 activity in lateral root control. Interestingly, this was equally the case when all initiated lateral roots, including pre-emerged root primordia as well as emerged lateral roots, were considered (Fig. 2c, Supplementary Fig. 2). Lateral root initiations were also reduced in grafted plants expressing pUBQ1::MIR2111-3 in their shoots compared to wild type/wild type control grafts (Fig. 2d). Using Crispr-CAS9 technology, we generated line mir2111-3-1, which possesses a 12 bp deletion in the stem-loop region of the MIR2111-3 locus in immediate proximity to the mature miRNA2111a sequence (Supplementary Fig. 3a–c). mir2111-3-1 plants showed significantly lower miR2111 abundance than wild type plants (Supplementary Fig. 4a) and, consistently, higher TML transcript levels (Supplementary Fig. 4b). In line with the observed reduced primordium formation in miR2111 overexpressors (Fig. 2c, d), lateral root initiation numbers were higher in mir2111-3-1 plants compared to wild type plants (Fig. 2e), and grafts of mir2111-3-1 shoots on wild type root stocks equally showed an enhanced lateral root initiation compared to control grafts. This is consistent with a shoot specific expression pattern of the MIR2111-3 locus (Fig. 2f, g), and confirms that shoot miR2111 is required for lateral root initiation control. Taken together, these data suggest that shoot-derived miR2111 is both sufficient and necessary for modulating lateral root initiations via TML (Fig. 2c–e, Supplementary Fig. 3a–c). Interestingly, this is in addition to the described function of shoot-derived miR2111 in systemic nodule number control in the context of symbiosis autoregulation (Supplementary Fig. 4c)11,15.

a Root phenotype of Gifu wild type (wt(G)), tml-6 and pUBQ1::MIR2111-3 plants (2111ox). Scale bars equal 1 cm. b Emerged LR numbers in wt, tml-6, tml-5 and 2111ox plants. c–e Number of LR initiations (emerged and primordial stages combined) in wt(G), 2111ox and tml-6 plants (c), on 2111ox / wt(G) (shoot / root) grafts compared to wt(G) / wt(G) control grafts (d), in mir2111-3-1 compared to wt(G) plants (e) and on mir2111-3-1 / wt(G) (shoot / root) grafts compared to wt(G) / wt(G) control grafts (f). f light grey dots represent data points not considered in the statistical analysis due to strong divergence of primary root length in the respective plants from the mean. g Plants expressing pMIR2111-3:GUS show pronounced GUS activity in leaf veins, while roots are free of visually traceable activity. Scale bar equals 1 cm. A total of 30 tested plants showed a similar expression pattern. b Datapoints are identical to datapoints at 0 mM nitrate in Fig. 3a. a–e Plants grown at 0 mM nitrate. Comparisons used Student’s t-test (*p ≤ 0.05; **p ≤ 0.01) (d, e) or analysis of variance (ANOVA) and post-hoc Tukey testing (p ≤ 0.05) (b, c), with distinct letters indicating significant differences. All experiments were in ecotype Gifu B-129 (wt(G)). a–g Plants were evaluated or harvested after two weeks of cultivation (a–c, e & g) or grafting (d, f). Sample size, replicates and exact p-values are listed in the Source Data file. Boxplot central line shows median value, box limits indicate the 25th and 75th percentile. Whiskers extend 1.5 times the interquartile range, or to the last datapoint. Individual datapoints are represented by dots.
The miR2111/TML regulon controls root branching in response to nitrate levels
Since root nodulation symbiosis, a known activity context of miR2111-TML, is an adaptation to nitrogen limitation, we hypothesized that this regulon may also help adapting root architecture to nitrogen availability.
Lotus showed enhanced lateral root numbers under nitrogen starvation (Fig. 3a). This was ecotype-independent (Fig. 3a, b) and is consistent with the nitrate foraging responses reported in other plants17. Notably, this trend was only apparent for emerged lateral roots. Lateral root primordia, on the contrary, were more abundant under nitrate sufficient conditions compared to deficiency. This results in a positively nitrate-correlated (Fig. 3c, d) or nitrate-independent (Supplementary Fig. 5a) sum of initiated roots.

a, b Emerged LRs in (a) Gifu B-129 wild type (wt(G)), tml-6, tml-5 and pUBQ1::MIR2111-3 (2111ox), and in (b) MG20 wildtype (wt(M)) and tml-1 plants. c Number of LR initiations (emerged plus primordial stages) in wt(M) and tml-1 plants. d Simplified model of nitrate dependency of LR initiations in wt and tml mutant plants. e, f Relative mature miR2111 levels in shoots (e) and roots (f). g Relative TML levels in same wild type root systems as in (f). h Simplified model outlining nitrate dependency of miR2111 and TML levels, and root architectural responses. I, j Split root experiments. Relative miR2111 (i) and TML (j) levels in secondary roots of wt(M) plants. e–g, i, j qRT-PCR analyses. RNA levels are relative to those of two reference genes. a, b, e–g, i, j Tissue harvest / analysis after two weeks and (c) 10 days of cultivation. Comparisons used analysis of variance (ANOVA) and post-hoc Tukey. testing (p ≤ 0.05), with distinct letters indicating significant differences and additional Student’s t-test (i, j) comparing only the split roots (n.s. p > 0.05). c–h Trendlines are simplified and not to scale. Plants grown at indicated nitrate concentrations. Sample size, replicates and exact p-values are listed in the Source Data file. Dotplots show individual data points and a line indicating their average value. Boxplot central line shows median value, box limits indicate the 25th and 75th percentile. Whiskers extend 1.5 times the interquartile range, or to the last datapoint. Individual datapoints are represented by dots.
Following a likewise trend, mature miR2111 levels were negatively correlated with nitrate availability in a dosage-dependent manner in both shoots and roots (Fig. 3e, f), indicating an involvement in systemic nitrogen response signalling. The levels of TML transcripts, which were only detected in roots, showed a complementary, inverse pattern (Fig. 3g), suggesting TML suppression by systemic miR2111 under nitrogen starvation conditions. Indicating ecotype specific differences, this pattern was particularly apparent in the Lotus ecotype MG-20, consistent with a more pronounced responsiveness of lateral root initiations to nitrate availability compared to Gifu B-129 (Supplementary Fig. 5a–d).
Compared to wild type plants, both miR2111 overexpressors and tml mutants had a consistently lower number of lateral roots, and emerged lateral root number was independent of nitrate availability (Fig. 3a, b). The same was true for lateral root initiation, not only in the ecotype Gifu B-129, but also in MG-20, where a nitrate-dependent increase in lateral root primordia is more strongly pronounced than in Gifu B-129 (Fig. 3c, d and Supplementary Fig. 5a). Shoot specific overexpression of MIR2111-3 using heterografting experiments induced a loss of nitrate responsive lateral root initiation in chimeric plants with MG-20 root stocks (Supplementary Fig. 6a, b), suggesting that shoot-derived miR2111 efficiently represses this response. On this basis, we predicted that nitrate-independent TML transcript levels in Gifu B-129 (Supplementary Fig. 5d) may prevent adaptive primordia formation in this ecotype. Indeed, increased TML transcript levels in mir2111-3-1 compared to wild type plants (Supplementary Fig. 4b) resulted in a positive nitrate response of lateral root initiation numbers in the ecotype Gifu B-129 as well (Supplementary Fig. 6c), indicating that miR2111 mediated TML control is necessary for the ecotype-specific attenuation of root system response to nitrate observed in Gifu B-129. The combined phenotypic and molecular data suggests a role of the miR2111-TML regulon in lateral root initiation and adaptive emergence in response to nitrate, with miR2111 systemically repressing TML. Contrasting with their respective roles in symbiosis, our data identify miR2111, a positive regulator of nodule numbers, as a repressor of root primordia, and TML as a root primordial activator (Fig. 3h). The data further reveal that nitrate dependent regulation of primordia emergence into full lateral roots strictly requires the presence of functional TML (Fig. 3a–c), but does not correlate with TML transcript abundance (Fig. 3g, h, Supplementary Fig. 5d). This suggests involvement of additional factors in regulating nitrate responsive emergence of TML-dependent lateral root primordia.
Nitrate perception and nitrogen starvation have been found to trigger local and systemic responses involving physiological and morphological adaptations17. We thus performed split root assays to identify the trigger underlying miR2111 regulation under asymbiotic conditions. Roots growing on nitrogen starvation medium contained low miR2111 levels if other roots of the same plant experienced nitrate sufficiency (Fig. 3i). This suggests that miR2111 accumulation is not triggered by roots experiencing nitrate starvation, but rather is systemically repressed by roots exposed to nitrate sufficiency (Fig. 3i), implying that miR2111 levels are regulated through nitrate supply rather than deficiency. TML levels in these roots were complementary yet intermediate (Fig. 3j). Consistent with previous observations, TML abundance is thus likely subject to additional regulatory factors11.
Apart from nitrate, rhizobial infection triggers changes in miR2111 and TML transcript abundance (ref. 11; Supplementary Fig. 7 a–c), and miR2111 acts as a positive regulator of nodule organogenesis by repressing the nodulation inhibitor TML11. We thus wondered how miR2111-TML dynamics affect lateral root formation under symbiotic conditions. Interestingly, in both wild type and tml-6 mutant plants, symbiotic infection led to a decrease of lateral root initiations (Supplementary Fig. 7d), implying that an additional TML-independent regulation of lateral root initiation overlays miR2111-TML dependent primordium control under symbiotic conditions.
Systemic root control by miR2111/TML is phylogenetically conserved
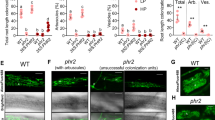
Root architecture adaptation to abiotic stimuli is an ancient necessity and a core developmental feature of land plants that is phylogenetically widespread17 and thus precedes the evolution of nitrogen-fixing nodulation symbiosis. Consistently, the miR2111-TML regulon is conserved in non-nodulating plants, including the nonsymbiotic plant Arabidopsis thaliana (Arabidopsis)11,14. Arabidopsis possesses two MIR2111 precursor loci generating a single isoform identical to LjmiR2111a (Supplementary Fig. 8a, b)11. Phylogenetic analysis revealed one putative TML orthologue, which we named HOMOLOGUE OF LEGUME TML (HOLT), featuring a miR2111 complementary site in the coding sequence11,14 (Supplementary Fig. 9a, b). Consistent with the expression pattern of MIR2111 loci in Lotus, Arabidopsis pMIR2111a/b:GUS expressing lines showed predominant GUS activity in leaf vein phloem cells (Fig. 4a, b and Supplementary Fig. 10a, b), suggesting systemic mobility18. These observations are in line with organ specific expression data19 (Supplementary Table 1). Similar to what has been previously observed for selected Lotus MIR2111 precursor genes15, moderate pMIR2111a/b:GUS activity was evident in mature root parts of Arabidopsis as well (Fig. 4a and Supplementary Fig. 10a), although MIR2111a/b precursor transcripts were not traceable in publicly available RNAseq datasets19 (Supplementary Table 1). Like in Lotus, Arabidopsis lateral root initiation numbers depend on nitrogen supply, peaking around 1 mM nitrate under long day conditions in plate-grown wild type plants (Fig. 4c). In comparison, plants grown under starvation or saturating conditions show reduced numbers of lateral root initiations (Fig. 4c). The observed increase of lateral root numbers under moderately deficient (1 mM nitrate) as compared to sufficient (10 mM nitrate) nitrogen supply has previously been associated with nitrogen dependent root architectural adaptations commonly referred to as foraging response1. To evaluate a possible role of the miR2111/HOLT regulon in nitrogen foraging related lateral root initiation in Arabidopsis, we generated transgenic lines overexpressing miR2111 under the control of a Cauliflower Mosaic Virus 35 s promoter fragment, showing a concomitant reduction in HOLT levels (Supplementary Fig. 11a, b). All tested lines showed reduced lateral root initiation compared to wild type plants in the T2 generation (Supplementary Fig. 11c). We chose a representative line, #3, for further propagation, as it showed stable overabundance of miR2111 and a corresponding reduction of TML transcript abundance in the T3 generation (Fig. 4d, e). We further isolated Arabidopsis holt-1 and holt-2 mutants lacking a traceable full-length HOLT transcript (Supplementary Fig. 12a, b). holt-1, holt-2 and p35s::MIR2111b plants showed significantly reduced lateral root initiations at low and moderate nitrate concentrations compared to wild type plants (Fig. 4f). Notably, they failed to show a traceable foraging response (Fig. 4f,g). Wild type plants exposed to severe nitrogen limitation repress lateral root development, a response known as a survival strategy1, which is thought to involve the nitrate transporter NRT1.120 as well as locally induced lateral root inhibition through the CLAVATA3/CLAVATA1 signalling module4. Transcript abundance of NRT1.1 and other NRTs was not significantly altered in holt-1 or p35s::MIR2111b as compared to wild type plants (Supplementary Fig. 13a–d). Consistent with a HOLT independent mechanism, a successive reduction in lateral root initiation numbers at nitrate levels <1 mM was retained in holt-1, holt-2 and p35s::MIR2111b in a similar way as in wild type plants (Fig. 4f, g). In wild type plants, miR2111 levels correlate positively with nitrate concentration (Fig. 4h), consistent with low HOLT levels at high nitrate supply (Fig. 4i). The integration of phenotypic and molecular data reveals that, in line with observations in Lotus, HOLT levels positively correlate with lateral root initiations (Fig. 4j). To investigate whether shoot-derived miR2111 is sufficient to regulate root architecture in Arabidopsis as observed in Lotus, we analysed p35s::MIR2111b/Col-0 (shoot/root) grafts. These had significantly less lateral root initiations than Col-0/Col-0 control grafts (Fig. 4k), confirming miR2111 as a systemically acting, mobile regulator of lateral root initiation across dicot plant lineages.

a, b Stably transformed A. thaliana (Arabidopsis) plants expressing pMIR2111b:GUS show predominant GUS activity in the phloem of leaf veins. b Leaf cross section of pMIR2111b:GUS plants. ph, phloem and xy, xylem. c Numbers of LR initiations at different nitrate concentration in Arabidopsis wild type (Col-0) plants. d, e miR2111 (d) and HOLT (e) levels in Col-0 and p35s::MIR2111b expressing plants (2111ox) at 1 mM nitrate. f LR initiations in holt-1, holt-2 and 2111ox plants compared to Col-0. g Schematic model of nitrate responsiveness of Arabidopsis lateral root initiations. holt-1 and 2111ox plants lack a foraging response at moderate nitrate starvation. h, i miR2111 (h) and HOLT (i) levels of Col-0 at varying nitrate concentrations. j Simplified model outlining nitrate dependency of miR2111 and HOLT levels, and of root architectural responses in Arabidopsis between 1 and 10 mM nitrate. k Number of LR initiations on 2111ox / Col-0 (shoot / root) grafts compared to Col-0 / Col-0 control grafts at 1 mM KNO3. l Combined simplified model of Arabidopsis and L. japonicus (Lotus) root responses to varying nitrate supply. HOLT positively correlates with LR initiations in both species, but nitrate dependent abundance patterns of both LR initiations and TML/HOLT levels are opposite. d, e, h, i qRT-PCR analyses. RNA levels are relative to those of two reference genes, whole plant tissue harvested 10 days after germination. c, f, k Analysis seven days after germination (c, f) or after graft regeneration (k). Comparisons used analysis of variance (ANOVA) and post-hoc Tukey testing (p ≤ 0.05), with distinct letters indicating significant differences (c, f, h, i) or Student’s t-test (*p ≤ 0.05) (d, e, k). a, b Scale bars equal 200 µm (a) or 100 µm (b). a, b all 21 tested plants of 3 independent lines showed a similar expression pattern. Analysis of three independent lines showed similar results. c, f, h, i Plants were grown at indicated nitrate concentrations using ½ strength MS media free of other nitrogen sources. c, f–j, l Trendlines are simplified and not to scale. Sample size, replicates and exact p-values are listed in the Source Data file. Dotplots show individual data points and a line indicating their average value. Boxplot central line shows median value, box limits indicate the 25th and 75th percentile. Whiskers extend 1.5 times the interquartile range, or to the last datapoint. Individual datapoints are represented by dots.
In line with divergent habitat requirements and ecological strategies of the symbiotic Lotus21,22 and the asymbiotic ruderal Arabidopsis23, abundance patterns of lateral root primordia with respect to external nitrogen supply were distinct in these two species (Fig. 4l). Yet, consistent with a conserved positive role of TML/HOLT in nitrate-dependent lateral root initiation, TML/HOLT RNA levels were upregulated in both species under nitrate conditions triggering abundant lateral root primordia. Accordingly, in either species, miR2111 levels were low under such conditions, in line with a negative effect on TML/HOLT levels and lateral root initiation. The dynamic response pattern of the Arabidopsis root system reflected in integrating distinct and functionally overlapping regulatory nodes (refs. 4,20, this study) indicates its capacity to populate a wide variety of soils23. Our data suggests that Lotus, as a pioneer lineage that is primarily competitive on nitrogen poor soils24, initiates additional root primordia under starvation conditions that have a limiting effect on Arabidopsis root architecture (Fig. 4l). The lack of a strong nitrogen starvation response in Lotus could be explained by the formation of nitrogen fixing symbiosis, which prevents nitrogen starvation even on nitrogen poor soils.
An important role of the miR2111-TML/HOLT regulon in adapting plant root systems to their natural habitat is in line with the observed differences in lateral root abundance patterns between Lotus MG-20 and Gifu ecotypes (Fig. 3a–c, Supplementary Fig. 5a, Supplementary Fig. 6a–c). Lotus japonicus underwent intense diversification during evolution and encompasses more than 130 ecotypes that have adapted to a wide range of environmental conditions on the Japanese Islands21. A time course experiment revealed an increasing difference between wild type and tml-1 in lateral root numbers over time. Here, tml-1 plants showed significantly less biomass production in both below- and aboveground tissues compared to wild type (Supplementary Fig. 14a, b), suggesting that root architecture adaption plays an important role in plant productivity and fitness. We have no evidence for a direct involvement of the miR2111-TML/HOLT regulon in nitrate uptake, and mRNA levels of nitrate transporter genes NRT1.1, NRT1.5, NRT2.1 and NRT3.1 are unaltered in holt-1 and p35s::MIR2111b lines compared to wild type controls (Supplementary Fig. 13). This is in contrast to CEP/CEPD/CEPDL2 mediated regulation of nitrate uptake via NRT2.1regulation7,8, and suggests an indirect role of the miR2111-TML/HOLT regulon in nutrient uptake regulation by altering the extent of the root surface area (Supplementary Fig. 13).
Alteration of the root depletion zone also affects the uptake of other nutrients. Interestingly, in Arabidopsis, miR2111 was shown to be induced by phosphate starvation25, and Arabidopsis is known to adapt its root architecture to phosphate availability26. This could hint to a more general role of miR2111 in adapting root architecture to nutrient availability.
The presented data identify miR2111 and TML/HOLT as conserved factors in root architectural control, suggesting that they were evolutionarily co-opted by rhizobial nodulation symbiosis to regulate root responses to symbiotic bacteria, and organogenesis of nodule organs11. Consistent with this hypothesis, the transcription factors SCARECROW and SHORTROOT27, as well as ASYMMETRIC LEAVES2-LIKE1828,29 and STYLISH30 mediating auxin signalling hold dual roles in nodule organogenesis and root development, and comparative transcriptome analysis of lateral root and nodule primordia further supports generic ties between these organs28.
Our data reveal the miR2111-TML/HOLT regulon as a key factor in systemic control of root system architecture and lateral root organ number. An exciting future challenge will be determining the molecular activity of the TML/HOLT protein, a proposed component of the E3 Ubiquitin ligase complex in Arabidopsis31 with a possible role in mediating degradation of target transcription factors31. Determining downstream effectors will help us better understand how the miR2111-TML/HOLT regulon functionally integrates with hormonal networks and other regulators of root growth.
Methods
Plant and bacterial resources
Plants for root architecture analyses, qPCR assays and GUS stainings were Lotus japonicus L. ecotype Gifu B-129 (wild type, tml-5 (line ID 30013998), tml-6 (line ID 30086992) and pMIR2111-3:GUS11,32) and ecotype MG-20 (wild type and tml-133). Generation of stable transgenic plants expressing pUBQ1:MIR2111-334 and MIR2111-3 knockout lines followed a published procedure based on callus regeneration11,35 and made use of Agrobacterium tumefaciens AGL1 and L. japonicus ecotype Gifu B-129.
Further, A. thaliana Col-0 wild type, holt-1 (line ID SALK_044075.49.80.x) and holt-2 (line ID SALK_140092.27.55.x) were used. Generation of p35::MiR2111b expressing plants as well as pMIR2111a/b:GUS lines was done via floral dipping using A. tumefaciens GV3101. Cloning approaches made use of E. coli strains TOP10 or DB3.1. Plants were infected with M. loti MAFF30309936 expressing DsRED bacteria.
Construct generation
For p35s driven miRNA overexpression, the transcription start site upstream of the MIR2111b stemloop was predicted using the publicly available Softberry toolset with standard settings (http://www.softberry.com/berry.phtml?topic=tssplant&group=programs&subgroup=promoter). The entire precursor gene, including 80–100 bp downstream of the miRNA stem loop, was amplified from A. thaliana Col-0 genomic DNA using primers carrying overhangs for subsequent cloning into the gateway vector PGWB602. For the pMIR2111a/b:GUS lines, a three kb region upstream of the precited stem loop was cloned into the vector PMDC163 using gateway cloning. For Golden Gate technology-based generation of CRISPR/Cas9 constructs targeting the MIR2111-3 locus a codon optimized Cas9 endonuclease from the Streptococcus pyogenes containing the potato IV2 intron driven by a minimal 35 s promotor was used. Two gRNAs GGTAATCTGCATCCTG and GAGTCGGTATATATTGGGTC were predicted using CLC Main Workbench 8 (Qiagen). Primers can be found in Supplementary Table 2.
Lotus plant growth
For plant growth and phenotyping, L. japonicus seeds were surface scarified, sterilized using sodium hypochloride solution containing 1 g/l NaClO, imbibed in ddH2O and transferred to sterile ¼-strength B&D medium37 with 1% (w/v) phyto agar (Duchefa Biochemie). Following stratification for three days at 4 °C, seeds were germinated at 21 °C in constant darkness for two (MG-20) or three (Gifu B-129) days. For growth on plates, seedlings were transferred to 12 × 12 cm square plastic dishes containing 50 ml ¼-strength B&D / 1% (w/v) phyto agar medium supplemented with KNO3 at indicated concentrations. Plants were grown at long day conditions (16 h light, 21 °C / 8 h dark, 17 °C). Roots were shaded from direct light. For root architecture evaluation and quantification of molecular miR2111 and TML levels, plants were grown for twoweeks. For quantification of lateral root initiations plants were grown for ten days.
Arabidopsis plant growth
For plant growth and phenotyping, A. thaliana seeds were sterilized by 30 minutes incubation in a solution of 70% (v/v) ethanol and 0.05% (v/v) Triton X-100. Sterile seeds were transferred to 12×12 cm square plastic dishes containing 50 ml ½-strength MS medium38 without nitrogen / 1% (w/v) phyto agar medium supplemented with KNO3 at indicated concentrations and stratified for three days at 4 °C. Plants were grown at long day conditions (16 h light, 21 °C / 8 h dark, 17 °C). Roots were shaded from direct light. For quantification of molecular miR2111 and HOLT levels, plants were grown for ten days. For quantification of lateral root initiations plants were grown for seven days.
Lotus grafting
Plants were treated and germinated as described above. After germination plants were transferred to ¼-strength B&D / 1 mM KNO3 / 1% (w/v) phyto agar medium. Plates were kept in darkness for two days, then eight days in long day conditions. For graftings, seedlings were cut near the lower end of the hypocotyl, and immediately submerged in water. New shoots were transplanted onto root stocks and arrested using silicone tubing (∅ 0.64 mm). Grafted plants were transferred to fresh medium and covered with filter paper soaked in ddH2O, then grown at long day conditions for two to three weeks. Prior to phenotyping or tissue harvest, tubing was removed from chimeric plants to determine grafting success. Lotus hetero-grafting involving two distinct ecotypes followed a different procedure.
Lotus hetero grafting
Plants were treated and germinated as described above. After germination plants were transferred to ¼-strength B&D / 1 mM KNO3 / 1% (w/v) phyto agar medium. Plates were kept three days in long day conditions. For graftings, seedlings were cut near the middle of the hypocotyl, and immediately submerged in water. New shoots were transplanted onto root stocks and arrested using silicone tubing (∅ 0.5 mm, ~3 mm long). Grafted plants were transferred to fresh medium, then kept at 26 °C 22 h light for five days to enable graft site regeneration. Afterwards grafted plants were incubated for two more weeks at long day conditions (16 h light, 21 °C / 8 h dark, 17 °C). Prior to phenotyping or tissue harvest, tubing was removed from chimeric plants to determine grafting success.
Arabidopsis grafting
Plants were treated and germinated as described above (section ‘Arabidopsis plant growth’), and grown for five days on ½-strength MS / 1% (w/v) phyto agar medium medium with full nitrogen content38. Grafting followed a published protocol, utilizing sterile precision forceps and a sapphire blade39. For grafting one cotyledon of Arabidopsis plants was removed and cut at the hypocotyl. Cut Arabidopsis plants were reassembled on a sterile nitrocellulose membrane on sterile water soaked Whatman paper. After grafting, plants left to recover at 26 °C for four days, then grown for seven more days on ½-strength MS medium containing 100 µM KNO3 at long day conditions before evaluation.
Split root assay
Plants were treated and germinated as described above (section ‘Lotus plant growth’). After germination, root tips were cut off and plants transferred to ¼-strength B&D/0.5 mM KNO3/1% (w/v) phyto agar medium. After 10 days at long day conditions, plants which had generated two secondary roots were selected for onward processing and transferred to plates containing slices of ¼-strength B&D/1.5% (w/v) phyto agar medium. Plants were positioned in a way that secondary roots were placed on separate agar patches not in physical contact with each other and containing KNO3 concentrations as indicated. Plants were grown for 13 more days under long day conditions until phenotypic evaluation or tissue collection.
Root phenotypic analysis
Plate grown plants were scanned using a conventional scanning system (CanoScan 8800 F, Canon). Image processing made use of OpenCV (https://opencv.org/) functions. Root architectural traits were measured using an in-house Python script. The script was optimized to recognize Lotus roots by color contrast and relied on manual confirmation.
RNA extraction and quantitative PCR (qRT-PCR) assays
For total RNA extractions, plant or aphid tissue was shock frozen in liquid N. Total RNA was extracted by a modified Lithium Chloride-TRIzol LS (ThermoFisher) protocol40. Plant RNA was extracted from tissue of at least ten independent plants per biological replicate. RNA was eluted in DEPC-treated water, RNA concentration was determined using a Nanodrop device (ThermoFisher). RNA was DNAse treated using DNAseI (ThermoFisher) according to manufacturer guidelines. cDNA was prepared using SuperScriptIV (ThermoFisher) or RevertAid (ThermoFisher) reverse transcriptase following a previously optimized pulsed protocol11,41. Briefly, RNA and primers (2 µM odT, 0,5 µM specific primers) were mixed and incubated for 5 min at 65 °C. The remaining reaction mix was assembled and incubated at 16 °C for 30 mins followed by 60 cycles (30 °C for 30 s, 42 °C for 30 s and 50 °C for 1 s) and 5 min at 85 °C for enzyme inactivation. Stemloop primers for reverse transcription of small RNAs were designed such that the six basepairs at the 5’ end of the stemloop primer were complementary to six nucleotides at the 3’ end of the small RNA, for reverse transcription of Lotus mRNAs oligo dT primer was used, for A. thaliana RNA only locus specific primers were used for reverse transcription (Supplementary Table 3). The RT-reaction was assembled according to manufacturer’s guidelines using 500 ng of total RNA. qRT-PCRs were assembled using SensiFAST™ SYBR® No-ROX mastermix (Bioline) at 10 µl reaction size and 500 nM primer concentration. Levels of target genes were normalized to levels of two independent reference genes, Lotus ATP SYNTHASE2 and Lotus PROTEIN PHOSPHATASE2a or Arabidopsis PROTEIN PHOSPHATASE2a and Arabidopsis UBIQUITIN EXTENSION PROTEIN 2 or U6 (Supplementary Fig. 11a). qRT-PCR reactions were executed in a BioRad CFX384 lightcycler (BioRad). Primers are listed in Supplementary Table 4. Data analysis made use of LinRegPCR42.
Aphids
For aphid experiments, we used Planococcus citri43, which could be propagated on L. japonicus as sole host plant. Lotus plants were infected with aphids by placing an infested host stem with a small aphid population onto young, four week-old plants growing in a 3:1 clay granule (2–5 mm, Lamstedt): vermiculite (3–6 mm, Isola Vermiculite GmbH) mixture saturated with ¼-strength B&D medium. After two weeks, aphids were collected and snap frozen in liquid nitrogen. RNA extraction, as well as DNAse treatment, cDNA synthesis and qRT-PCRs, were performed as described. For qRT-PCR experiments on aphid RNA extracts we used aphid α-Tubulin as normalization reference44.
Staining and microscopic analysis
GUS staining and fixation was performed as described11. Plants were fixed by incubation in 1x phosphate buffer (50 mM NaH2PO4, 50 mM Na2HPO4, pH 7.0) supplemented with 4% Paraformaldehyde, followed by 3 washing steps using 1x phosphate buffer. For GUS staining, samples were incubated overnight at 37 °C in X-Gluc buffer (0.5 mg/ml X-Gluc, 1 mM K4(Fe(CN)6), 1 mM K3(Fe(CN)6), 0,05% Triton X-100 in 1x phosphate buffer). After 3 more washing steps in 1x phosphate buffer, plants were incubated in a buffer containing acetic acid, glycerol and ethanol (ratio 1:1:3) at 60 °C until tissue was cleared from chlorophyll.
Whole plant phenotypes were monitored, and photographs taken using a Leica MZ FLIII stereomicroscope. For analysis of semithin sections, fixed, GUS-stained roots or leaves were further dehydrated and embedded in resin (Kulzer Technovit 7100). Sections were prepared using a Leica RM2065 microtome, then analyzed and documented using a Zeiss Imager M2 microscope. For quantification of lateral root initiations, roots were separated from shoots and fixed using 4% Paraformaldehyde in 1x PBS buffer, cleared using ClearSee (10% Xylitol, 15% sodium deoxycholate and 25% Urea in ddH2O) and stained with Fluorol Yellow (0.01% in 96% Ethanol) as described45. Stained roots were scanned using a Leica SP8 confocal microscope, and images were used for phenotypic analysis. Fluorol yellow was imaged at λ = 488 nm excitation and λ = 520–588 nm emission, and additionally, transmission white light was observed. The resulting integrated images of whole roots allowed quantification of lateral root primordia at early, pre-emergence stages. Lateral root initiations include pre-emergence primordia as well as emerged lateral roots irrespective of the developmental stage (Supplementary Fig. 2).
Data analysis and graphical representation
Data analysis made use of Python 3.7.x using the libraries Statsmodels and Pandas. Plots were generated with the python libraries Matplotlib and Seaborn. Boxplot center lines show the medians, outer box limits indicate the 25th and 75th percentiles. Whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles, or the last data point. Data points are represented as dot. Dotplots center lines indicate the average value of all data points. All statistical tests used were two-sided. For pairwise comparison we used t-tests for multi-comparison we used ANOVA and post hoc Tukey-HSD testing. Results of ANOVA or t-test analyses, biological replicate numbers and individual datapoints are listed in the Source Data file.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All primary data and images analysed in the context of this study are available from the corresponding author upon request. Source data are provided with this paper.
References
Giehl, R. F. & von Wiren, N. Root nutrient foraging. Plant Physiol. 166, 509–517 (2014).
Maghiaoui, A. et al. The Arabidopsis NRT1.1 transceptor coordinately controls auxin biosynthesis and transport to regulate root branching in response to nitrate. J. Exp. Bot. 71, 4480–4494 (2020).
Marchive, C. et al. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 4, 1713 (2013).
Araya, T. et al. CLE-CLAVATA1 peptide-receptor signaling module regulates the expansion of plant root systems in a nitrogen-dependent manner. Proc. Natl Acad. Sci. USA 111, 2029–2034 (2014).
Hayashi-Tsugane, M. & Kawaguchi, M. Lotus japonicus HAR1 regulates root morphology locally and systemically under a moderate nitrate condition in the absence of rhizobia. Planta 255, 95 (2022).
Okamoto, S., Shinohara, H., Mori, T., Matsubayashi, Y. & Kawaguchi, M. Root-derived CLE glycopeptides control nodulation by direct binding to HAR1 receptor kinase. Nat. Commun. 4, 2191 (2013).
Ohkubo, Y., Kuwata, K. & Matsubayashi, Y. A type 2C protein phosphatase activates high-affinity nitrate uptake by dephosphorylating NRT2.1. Nat. Plants 7, 310–316 (2021).
Ota, R., Ohkubo, Y., Yamashita, Y., Ogawa-Ohnishi, M. & Matsubayashi, Y. Shoot-to-root mobile CEPD-like 2 integrates shoot nitrogen status to systemically regulate nitrate uptake in Arabidopsis. Nat. Commun. 11, 641 (2020).
Luo, Z. et al. NLP1 reciprocally regulates nitrate inhibition of nodulation through SUNN-CRA2 signaling in Medicago truncatula. Plant Commun. 2, 100183 (2021).
Luo, Z. et al. The small peptide CEP1 and the NIN-like protein NLP1 regulate NRT2.1 to mediate root nodule formation across nitrate concentrations. Plant Cell 35, 776–794 (2023).
Tsikou, D. et al. Systemic control of legume susceptibility to rhizobial infection by a mobile microRNA. Science 362, 233–236 (2018).
Gautrat, P., Laffont, C. & Frugier, F. Compact Root Architecture 2 promotes root competence for nodulation through the miR2111 systemic effector. Curr. Biol. 30, 1339–1345.e1333 (2020).
Pant, B. D. et al. Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiol. 150, 1541–1555 (2009).
Takahara, M. et al. Too much love, a novel Kelch repeat-containing F-box protein, functions in the long-distance regulation of the legume-Rhizobium symbiosis. Plant Cell Physiol. 54, 433–447 (2013).
Okuma, N., Soyano, T., Suzaki, T. & Kawaguchi, M. MIR2111-5 locus and shoot-accumulated mature miR2111 systemically enhance nodulation depending on HAR1 in Lotus japonicus. Nat. Commun. 11, 5192 (2020).
Devers, E. A. et al. Movement and differential consumption of short interfering RNA duplexes underlie mobile RNA interference. Nat. Plants 6, 789–799 (2020).
Oldroyd, G. E. D. & Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 368, eaba0196 (2020).
Skopelitis, D. S. et al. Gating of miRNA movement at defined cell-cell interfaces governs their impact as positional signals. Nat. Commun. 9, 3107 (2018).
Klepikova, A. V., Kasianov, A. S., Gerasimov, E. S., Logacheva, M. D. & Penin, A. A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 88, 1058–1070 (2016).
Krouk, G. et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 18, 927–937 (2010).
Shah, N. et al. Extreme genetic signatures of local adaptation during Lotus japonicus colonization of Japan. Nat. Commun. 11, 253 (2020).
Szczyglowski, K. & Stougaard, J. Lotus genome: pod of gold for legume research. Trends Plant Sci. 13, 515–517 (2008).
Pigliucci, M. Ecology and evolutionary biology of Arabidopsis. Arabidopsis Book 1, e0003 (2002).
Griesmann, M. et al. Phylogenomics reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 361, eaat1743 (2018).
Hsieh, L. C. et al. Uncovering small RNA-mediated responses to phosphate deficiency. Arabidopsis Deep Seq. Plant Physiol. 151, 2120–2132 (2009).
Williamson, L. C., Ribrioux, S. P., Fitter, A. H. & Leyser, H. M. Phosphate availability regulates root system architecture in Arabidopsis. Plant Physiol. 126, 875–882 (2001).
Dong, W. et al. An SHR-SCR module specifies legume cortical cell fate to enable nodulation. Nature, https://doi.org/10.1038/s41586-020-3016-z (2020).
Schiessl, K. et al. NODULE INCEPTION recruits the lateral root developmental program for symbiotic nodule organogenesis in Medicago truncatula. Curr. Biol. 29, 3657–3668 e3655 (2019).
Soyano, T., Shimoda, Y., Kawaguchi, M. & Hayashi, M. A shared gene drives lateral root development and root nodule symbiosis pathways in Lotus. Science 366, 1021–1023 (2019).
Shrestha, A. et al. Lotus japonicus Nuclear Factor YA1, a nodule emergence stage-specific regulator of auxin signalling. N. Phytol. 229, 1535–1552 (2021).
Schumann, N., Navarro-Quezada, A., Ullrich, K., Kuhl, C. & Quint, M. Molecular evolution and selection patterns of plant F-box proteins with C-terminal kelch repeats. Plant Physiol. 155, 835–850 (2011).
Malolepszy, A. et al. The LORE1 insertion mutant resource. Plant J. 88, 306–317 (2016).
Magori, S. et al. Too much love, a root regulator associated with the long-distance control of nodulation in Lotus japonicus. Mol. Plant Microbe Interact. 22, 259–268 (2009).
Lazo, G. R., Stein, P. A. & Ludwig, R. A. A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. Biotechnol. (N. Y) 9, 963–967 (1991).
Thykaer, T., Schauser, L., Danielsen, D., Finneman, J. & Stougaard, J. in Cell Biology: A laboratory handbook Vol. 3 518–525 (Academic Press, 1998).
Maekawa, T. et al. Gibberellin controls the nodulation signaling pathway in Lotus japonicus. Plant J. 58, 183–194 (2009).
Broughton, W. J. & Dilworth, M. J. Control of leghaemoglobin synthesis in snake beans. Biochem J. 125, 1075–1080 (1971).
Murashige, T. & Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologica Plant. 15, 473–497 (1962).
Melnyk, C. W. In Plant Hormones. Methods in Molecular Biology Vol. 1497 (eds J. Kleine-Vehn & M. Sauer) 9-18 (Humana Press, 2017).
Holt, D. B. et al. micro RNA 172 (miR172) signals epidermal infection and is expressed in cells primed for bacterial invasion in Lotus japonicus roots and nodules. N. Phytol. 208, 241–256 (2015).
Varkonyi-Gasic, E., Wu, R., Wood, M., Walton, E. F. & Hellens, R. P. Protocol: a highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods 3, 12 (2007).
Ruijter, J. M. et al. Amplification efficiency: linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 37, e45 (2009).
Danzig, E. M. & Gavrilov, I. A. Mealybugs of the genera Planococcus and Crisicoccus (Sternorrhyncha: Pseudococcidae) of Russia and adjacent countries. Zoosystematica Rossica 19, 39–49 (2010).
Zheng, L. et al. Selection of reference genes for RT-qPCR analysis of Phenacoccus solenopsis (Hemiptera: Pseudococcidae) sex-dimorphic development. J. Integr. Agric. 18, 854–864 (2019).
Sexauer, M., Shen, D., Schön, M., Andersen, T. G. & Markmann, K. Visualizing polymeric components that define distinct root barriers across plant lineages. Development 148, dev199820 (2021).
Acknowledgements
We thank Johanna Schröter and the ZMBP gardening team for dedicated plant care, Caterina Brancato for help with plant transformations, Dugald Reid for help with construct generation, Angela Fischer for designing Supplementary Figure 1b, and Laura Ragni for discussions. Our thanks further go to Christoph Weiste, Wolfgang Dröge-Laser and Arthur Korte for critical reading of the manuscript. We apologize to authors whose work could not be cited due to space limitations. This research was supported by the German Research Foundation (grant CRC1101, project C07), Ministry of Science, Research and Art of Baden-Wuerttemberg (Az:7533-30-20/1).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
M.Se., H.B., M.Sch., E.R., C.W. and U.H. performed experiments and analyzed data; K.M., M.Se. and H.B. conceived and designed research; K.M. and M.Se. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Takuya Suzaki and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sexauer, M., Bhasin, H., Schön, M. et al. A micro RNA mediates shoot control of root branching. Nat Commun 14, 8083 (2023). https://doi.org/10.1038/s41467-023-43738-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-023-43738-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
