
Abstract
The tumor microenvironment (TME) in pancreatic ductal adenocarcinoma (PDAC) is a complex ecosystem that drives tumor progression; however, in-depth single cell characterization of the PDAC TME and its role in response to therapy is lacking. Here, we perform single-cell RNA sequencing on freshly collected human PDAC samples either before or after chemotherapy. Overall, we find a heterogeneous mixture of basal and classical cancer cell subtypes, along with distinct cancer-associated fibroblast and macrophage subpopulations. Strikingly, classical and basal-like cancer cells exhibit similar transcriptional responses to chemotherapy and do not demonstrate a shift towards a basal-like transcriptional program among treated samples. We observe decreased ligand-receptor interactions in treated samples, particularly between TIGIT on CD8 + T cells and its receptor on cancer cells, and identify TIGIT as the major inhibitory checkpoint molecule of CD8 + T cells. Our results suggest that chemotherapy profoundly impacts the PDAC TME and may promote resistance to immunotherapy.
Similar content being viewed by others

Introduction
Pancreatic ductal adenocarcinoma (PDAC) is a highly lethal cancer with a five-year survival rate of approximately 11%1. With increasing incidence, it is projected to become the second leading cause of cancer-related deaths in the United States by 20302. Current treatment options are limited. Only 15% to 20% of patients qualify for upfront surgery with curative intent; the remaining patients present with unresectable locally advanced disease or distant metastases. Nearly all patients receive adjuvant, neoadjuvant, or palliative chemotherapy3. Recent advances in immunomodulating drugs, such as checkpoint inhibitors, have yet to show encouraging results in PDAC4,5. A deeper understanding of pancreatic cancer biology is needed to develop better therapeutics and improve outcomes.
Although recent genomic and transcriptomic studies on bulk tumor samples identified key molecular drivers (notably KRAS, TP53, CDKN2A, and SMAD4) and cancer cell subtypes (basal and classical), these findings have yet to translate into treatment advances6,7,8,9. Recent investigations into the PDAC tumor microenvironment (TME) revealed that this complex ecosystem is a critical mediator of therapeutic resistance and tumor progression10,11. Single-cell RNA sequencing (scRNA-seq) has emerged as a valuable tool in precision medicine by enabling in-depth characterization of the TME and serial assessment on scant amounts of tissue12,13,14,15,16,17. While recent single-cell studies have provided insights into PDAC, there is an unmet need to characterize the effects of chemotherapy on the TME18,19,20,21,22.
For a comprehensive view of the PDAC TME, and its response to chemotherapy in a real-time clinical context, here we perform scRNA-seq on freshly collected tumor specimens from 27 PDAC patients. We observe significant heterogeneity in the malignant epithelial compartment, with most tumors displaying a mixture of basal-like and classical-like cells. Furthermore, we examine the composition of CAFs and discrete groups of immune cells in the TME and execute comparative analyses on naive and chemotherapy-treated samples, which we confirm in an external dataset. Our data show that chemotherapy profoundly alters the PDAC TME, inducing features consistent with tumor progression and refractoriness to immunotherapy by downregulation of inhibitory checkpoint molecules in CD8 + T cells.
Results
Single-cell analysis reveals the transcriptomic landscape in PDAC
PDAC specimens were acquired via surgical resection of primary pancreatic lesions (n = 10) or by endoscopic/interventional radiology–guided biopsies of either primary pancreatic lesions (n = 7) or liver metastases (n = 10) (Fig. 1A, Supplementary Table 1). Of the 27 total patients, seven received chemotherapy (FOLFIRINOX-based n = 4, gemcitabine/abraxane-based n = 3) prior to tissue collection, while 20 patients had not been treated at the time of specimen acquisition (Fig. 1A, Supplementary Table 1). Evaluating the treated patients using the RECIST criteria, we found that two had stable disease, two partial response, three progressive disease, and one was nonevaluable (Supplementary Table 2). We prepared single-cell suspensions from tumor specimens and sequenced them using the 10x Genomics Chromium system (Fig. 1B, see Methods for details). Histopathological assessment was done on each tumor specimen to verify malignancy, and samples underwent mutational profiling (Supplementary Data 1, 2). After standard processing and quality filtering of the raw sequencing data (see Methods), a total of 139,446 cells were retained for analysis. Unsupervised, graph-based clustering and visualization via uniform manifold approximation and projection (UMAP) revealed ten distinct cell clusters that were annotated using canonical markers and gene expression profiles (Fig. 1C, D, Supplimentary Fig. 1A, Methods). Three major clusters were identified as epithelial, T/natural killer (NK), and myeloid cells. We further identified CAFs, endothelial cells, B/plasma cells, and a small mast cell population among our clusters. The proportional distribution of these major cell types was similar across all groups, with the exception of biopsies from liver metastases and primary PDAC which had quantitatively lower CAFs and endothelial cells than resection samples. We also identified an elevated amount of proliferating cancer cells in liver biopsies (Fig. 1E; Supplementary Fig. 1B). To ensure that tissue processing did not significantly change gene expression in our samples, we applied hypoxia and dissociation signatures23,24 to our dataset. This analysis showed that all scSeq samples contained very few hypoxic cells, with no significant differences between procedures (Supplimentary Fig. 1C), and demonstrated low dissociation signatures independent of procedure (Supplimentary Fig. 1D).

A Clinicopathologic characteristics for each patient sample including treatment status, stage, Moffitt subtype, and procedure. EUS endoscopic ultrasound. B Workflow depicting sample acquisition, processing, and analysis. Designed with BioRender©. C UMAP, embedding of the expression profile of 139,446 cells that passed quality control from n = 27 patient samples. Distinct clusters are annotated and color-coded. CAF cancer-associated fibroblast, NK natural killer, UMAP uniform manifold approximation and projection. D Heatmap of the five most differentially expressed genes within each cluster. Colors correspond to the cluster colors in Fig. 1C. E Proportional distribution of each cell cluster by individual sample.
The epithelial compartment reveals heterogeneous malignant subtype composition
The epithelial compartment evinced sample-specific differences while the mesenchymal and immune compartments clustered together across all samples (Supplimentary Fig. 2A). This suggests that non-epithelial cells of the PDAC TME share more common transcriptional signatures across all patients than their malignant counterparts within the same tumor. Since some epithelial cells also clustered across samples, we further assessed the proportion of malignant cells in the total epithelial compartment (Fig. 2A). To this end, we performed copy number variation (CNV) analysis. The vast majority of epithelial cells showed several CNV events, indicating malignancy and distinguishing them from normal epithelial cells (Fig. 2B, Supplimentary Fig. 2B). We further classified malignant cells as either basal or classical based on their Moffitt subtype signature expression7. (Any signature gene sets referenced in this paper can be found in Supplimentary Data 3). All samples but one were comprised of a mixture of basal and classical cancer cells in varying proportions (Fig. 2C, D). Tumor subtype composition did not correlate with the amount of any major non-malignant cell types (Supplimentary Fig. 2C).

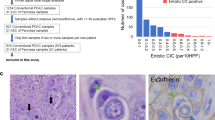
A Non-batch corrected UMAP of all epithelial cells, showing clustering by patient sample of most epithelial cells and clustering of some epithelial cells across samples. UMAP uniform manifold approximation and projection. B Non-batch corrected UMAP of malignant and normal epithelial cells as determined by InferCNV. C UMAP embedding of malignant epithelial cells labeled with Moffitt subtypes. Exemplary samples high in basal (P18) and classical (P08) cells are annotated. D Proportion of cells of each Moffitt subtype by sample. EUS endoscopic ultrasound. E Representative (n = 27) images of multiplex immunofluorescence across the subtype spectrum: higher in basal (upper panel, P03), mixed (middle panel, P24), and higher in classical (lower panel, P07) subtype tumors. Basal-to-classical ratio for each sample by scRNA-seq transcriptional analysis is shown on the right as a colored bar (dark = basal, light = classical). Channels (always including DAPI (blue)): GATA6 (green), CK19 (cytokeratin 19) (violet), CK17 (cytokeratin 17) (yellow), and merged. Scale bar = 100 μm.
We validated the Moffitt transcriptional subtypes on formalin-fixed, paraffin-embedded tissue (FFPE) sections using multiplex immunohistochemistry (IHC). We used the canonical markers cytokeratin 17 (CK17) for the basal subtype, GATA6 for the classical subtype, and cytokeratin 19 (CK19) as a pan-cancer cell marker. Representative samples with a higher percentage of basal or classical transcriptional subtypes were strongly positive for CK17 or GATA6, respectively (Fig. 2E, upper and lower panels), while a sample with a more even mixture of both subtypes showed an interspersed, more equal expression of GATA6 and CK17 (Fig. 2E, middle panels). Another patient sample with a mixture of both subtypes showed two distinct growth patterns with a CK17high region next to a GATA6high region (Supplimentary Fig. 2D). In general, GATA6 staining was more omnipresent, and CK17high cells often stained positive for GATA6. Of note, most samples included cells staining positive for both CK17 and GATA6 (Supplimentary Fig. 2E), consistent with a previous report that hypothesized an intermediate cell state25.
To test this intermediate cell state hypothesis in our dataset, we applied the basal, classical, and intermediate gene signatures from that report, which were derived from treatment-naive liver metastases, to malignant cells from our treatment-naive PDAC liver metastatic samples. Although their basal and classical signatures matched closely with ours, we did not see clear evidence of the intermediate signature (Supplimentary Fig. 2F). We also assessed the bulk RNA-derived subtype classifications of Collisson, Bailey and Puleo26,27,28. In our dataset, the Moffitt-basal signature showed overlap with the Collisson-quasi-mesenchymal and Puleo-pure-basal-like, while the Bailey-squamous signature did not show a strong correlation with the basal signatures. Similarly, the Moffitt-classical signature overlapped with the Collisson-classical and Puleo-pure-classical signatures, while the Bailey-progenitor signature did not show a strong correlation with the classical signatures. The less commonly found subtype signatures of Bailey (ADEX and immunogenic), Collisson (exocrine-like), and Puleo (desmoplastic) did not show strong expression in our dataset of CNV-confirmed cancer cells, suggesting that these might be derived from non-epithelial cells within tumor samples (Supplimentary Fig. 2G, H). These findings are supported by a TCGA publication from 2017 which examined sample purity and subtype classifications9. Further characterization of basal and classical cells by differential gene expression showed typical basal and classical genes upregulated in the respective subtypes (Supplimentary Fig. 2I). Pathway enrichment analysis revealed enrichment of EMT, angiogenesis, and extracellular matrix (ECM) interaction—features related to migration and invasion—in basal cells, while classical cells were enriched for metabolic pathways, especially lipid metabolism (Supplimentary Fig. 2J). These results emphasize the heterogeneity of malignant epithelial cells in PDAC and suggest that single-marker analysis may be insufficient to accurately identify cancer cell subtypes within a patient’s tumor.
Charting the CAF landscape in the PDAC tumor microenvironment
We next analyzed the mesenchymal compartment of the PDAC TME. Mesenchymal cell reclustering revealed seven distinct clusters (Fig. 3A). The two main clusters consisted of myofibroblastic CAFs (myCAFs) and inflammatory CAFs (iCAFs) (Fig. 3A, B)29. We did not find a strong antigen-presenting CAF (apCAF) signature (Fig. 3C). Notably, the apCAF signature was originally derived from mouse CAFs and may not translate to the human CAFs in our cohort. As apCAF features in human CAFs have been linked to co-expression of CD74 and human leukocyte antigen (HLA) molecules, we checked for expression of these genes in our CAF populations. A small number of cells in both the iCAF and myCAF subpopulations co-expressed CD74 and HLA genes (Fig. 3C, Supplementary Fig. 3A)19. Other clusters were identified as pericytes and Schwann cells, while a single sample provided a cluster of chondrocyte-like cells. Additionally, we detected a small proportion of cells expressing epithelial markers (Fig. 3A, B). We ruled out these cells being cancer cells undergoing EMT due to a lack of CNVs, low expression of EMT-related transcription factors (TFs) SNAIL1 and TWIST1, and low values of an EMT-TF independent partial EMT (pEMT) program signature30 (Supplimentary Fig. 3B). These cells were also not consistent with mesothelial cells, as mesothelin (MSLN) and other markers for mesothelial cells were minimally expressed; we therefore labeled this cluster “epithelial-like” (Supplimentary Fig. 3C).

A UMAP overview of the mesenchymal compartment. CAF cancer-associated fibroblast, iCAF inflammatory CAF, myCAF myofibroblastic CAF, UMAP uniform manifold approximation and projection. B Heatmap of the five most differentially expressed genes within each cluster. Colors correspond to the cluster colors in panel (A, C). iCAF, myCAF, and apCAF signatures from Elyada et al.19 overlaid on the UMAP embedding of our mesenchymal compartment. The iCAF and myCAF signatures are derived from human fibroblasts; the apCAF signature is derived from KPC mouse fibroblasts. apCAF antigen-presenting CAF. D Assessment of correlation (two-sided Pearson correlation) between myCAF/iCAF composition and basal/classical composition among our samples (n = 16). Samples are plotted as a function of the percentage of their CAF cells that are myCAFs and the percentage of their malignant epithelial cells that are basal. Samples with fewer than 40 total CAF cells were excluded; all but one of these samples were biopsies, which were not expected to pick up many mesenchymal cells. E Selected gene set enrichment analysis results from a comparison between the cells in the iCAF and myCAF compartments. ECM extracellular matrix.
The myCAF to iCAF ratio varied across samples, while the proportional distribution of both subpopulations in relation to Moffitt subtype distribution was not significantly different (Fig. 3D). In myCAFs, SDC131, a gene linked to CAF-mediated regulation of breast cancer cell migration, was upregulated, while in iCAFs, CCL1932, a gene linked to cytotoxic T cell recruitment by CAFs in lung cancer, was highly expressed (Supplimentary Fig. 3D, Supplimentary Data 4). Additionally, iCAFs showed enrichment for genes involved in the complement cascade and EGF signaling, whereas myCAFs demonstrated enrichment for ECM organization and smooth muscle contraction, supporting the notion of CAF heterogeneity and distinct functional characteristics of CAFs in the TME (Fig. 3E).
Charting the T/NK landscape in the PDAC tumor microenvironment
To better characterize tumor-infiltrating lymphocytes, we analyzed the T/NK population and identified 13 distinct clusters (Fig. 4A). CD4 + T cells clustered into four distinct subtypes: (i) CCR7+ CD4+ (high expression of naïve/central memory markers), (ii) IL7R + CD4 + (high expression of activation markers), (iii) FOXP3+ CD4+ (high expression of regulatory T cell genes), and (iv) CXCL13 + CD4 + (high expression of follicular helper genes) (Fig. 4B). CD8 + T cells clustered into three main subpopulations: (i) GZMH + CD8 + (high in cytotoxic gene expression), (ii) GZMK+ CD8+ (expressing effector memory genes), and (iii) ITGA1 + CD8 + (expression of tissue residency and exhaustion markers). We also identified a small ISG15 + cluster, shared between CD4 + and CD8 + T cells, with high interferon (IFN)-related gene expression. NK cells separated into a GNLY+ cluster highly expressing cytotoxic genes and an XCL1 + cluster with lower cytotoxic gene expression and higher expression of genes previously linked to tissue-resident NK cells33,34. Smaller clusters were identified as mast cells and plasma cells (Fig. 4A, B). T/NK subpopulations were heterogeneously distributed across samples and did not correlate with basal/classical subtyping (Supplimentary Fig. 4A, B).

A UMAP overview of the T/NK compartment. NK natural killer, UMAP uniform manifold approximation and projection. B The most differentially expressed gene within each T/NK cluster. C UMAP of the more granular CD8 + T cell reclustering reveals additional subpopulations. D The most differentially expressed gene within each CD8 + T cell cluster. E Hierarchical clustering of the CD8 + T cell subsets based on dysfunction and cytotoxicity scores. F Composition of the CD8 + T cell subset (excluding the two non-T cell clusters and the single-patient cluster).
Because our initial T/NK clustering did not identify distinct exhausted T cell subpopulations, we refined the resolution of the CD8 + T cell clustering (Fig. 4C). At this finer granularity, we discerned a small CTLA4 + CD8 + T cell cluster with high levels of exhaustion markers (CTLA4, HAVCR2, LAG3, TIGIT). Additional distinct CD8 + T cell clusters included (i) GZMK + CD8 + (high expression of effector memory genes), (ii) GZMH + CD8 + (high expression of cytotoxic genes), (iii) ITGA1 + CD8 + (high expression of tissue residency genes), (iv) SELL + CD8 + (expression of naive/central memory genes), and (v) ISG15 + CD8 + (expressing IFN activation genes) (Fig. 4C, D). We also found an epithelial-like cluster, an erythrocyte-like cluster, and an HSPA6 + cluster; the latter was mainly derived from one patient and did not show other distinct CD8 + subpopulation features (Fig. 4C, D).
To further assess exhaustion in our CD8 + T cell subpopulations, we scored each subset on a dysfunction and cytotoxicity scale35. As expected, GZMH + CD8 + T cells scored highest in cytotoxicity, and CTLA4 + CD8 + T cells highest in exhaustion (Fig. 4E). Except for CTLA4 + CD8 + T cells, the exhaustion state was similar between subsets (Fig. 4E). Exhausted CD8 + T cells represented only 2.8% of all CD8 + T cells (Fig. 4F). Moreover, exhausted T cells, along with most other CD8 + T cell subsets, were heterogeneously distributed across samples, independent of the Moffitt subtype composition of their epithelial compartments (Supplimentary Fig. 4C). Of note, ITGA1 + CD8 + T cells were the only subpopulation that was significantly negatively correlated with the basal subtype (Supplimentary Fig. 4D).
C1QC+ and SPP1+ TAMs comprise two distinct subpopulations within the myeloid cell compartment
Myeloid cells are an important component of the larger TME. We identified twelve distinct myeloid clusters (Fig. 5A). TAMs, representing the majority of myeloid cells, clustered into SPP1 + and C1QC + subpopulations (Fig. 5A, B). We also identified myeloid-derived suppressor cells (MDSCs), monocytes, and dendritic cells (DCs), which further clustered into conventional DCs 1-3 (cDC1-3) and plasmacytoid DCs (pDCs) (Fig. 5A, B). We did not detect any trends in myeloid composition across samples or in relation to Moffitt subtypes (Supplimentary Fig. 5A, B).

A UMAP overview of the myeloid compartment. DC dendritic cell, MDSC myeloid-derived suppressor cell, UMAP uniform manifold approximation and projection. B For each myeloid cluster, the two genes most differentially expressed between that cluster and the rest of the myeloid compartment. TAM, tumor-associated macrophages. C M1 (left) and M2 (right) polarization signatures in the macrophage populations. D Selected gene set enrichment analysis results from a comparison between SPP1 + and C1QC + TAMs. EMT, epithelial-mesenchymal transition. E Phagocytosis and angiogenesis scores for SPP1 + and C1QC + TAMs. One sample with fewer than 40 total TAM cells was excluded. (n = 26 samples; two-sided Wilcoxon rank-sum test; box plots centered around the median with hinges at 1st and 3rd quartiles and whiskers from hinge to max value or 1.5*IQR, whichever is smallest).
To better understand what distinguishes SPP1 + and C1QC + TAMs, we further characterized these two populations. Historically, TAMs have been classified as either M1 (pro-inflammatory) or M2 (pro-tumorigenic), so we assessed M1 and M2 signature expression in our TAMs13. While C1QC + TAMs showed higher M1 signature expression than SPP1 + TAMs, both SPP1 + and C1QC + subsets displayed strong M2 signature expression and therefore could not be clearly distinguished based on the M1/M2 classification (Fig. 5C). SPP1 + TAMs showed increased expression of S100A8 and S100A9, genes previously linked to immunosuppressive macrophages36, while C1QC + TAMs had increased expression of HLA-DRB5 and CXCL9, which is involved in the anticancer immune response37 (Supplimentary Fig. 5C, Supplimentary Data 4). In general, SPP1 + TAMs were enriched for genes involved in EMT, metabolism of carbohydrates, and hypoxia, while C1QC + TAMs showed enrichment for IFN signaling and antigen presentation (Fig. 5D). Consistent with the gene set enrichment analysis (GSEA) results and a recent colon cancer study38, SPP1 + TAMs were enriched for angiogenesis-related genes and C1QC + TAMs for phagocytosis (Fig. 5E).
To estimate whether the TAM subtype may affect overall survival in PDAC, we evaluated SPP1 + and C1QC + TAM signature expression in the Cancer Genome Atlas (TCGA) PDAC bulk tumor dataset9. While a similar analysis of the colorectal cancer (CRC) TCGA dataset implied worse overall survival for patients with a SPP1 + high /C1QC + low TAM combination38, we found that estimated TAM composition did not correlate with overall survival in the TCGA PDAC data (Supplimentary Fig. 5D).
Chemotherapy induces transcriptional changes in cancer cells independent of subtype
To analyze the effect of chemotherapy on the PDAC TME, we focused on primary pancreatic tumor specimens collected before treatment (treatment-naive, n = 11) and after treatment (n = 6). Specimens collected from liver biopsies were excluded from this part of the analysis in order to minimize potential confounding effects of mixing tumors from multiple sites. We also obtained an external dataset from Cui Zhou et al.39 to use as a confirmatory set to validate our findings; this dataset contains primary pancreatic tumor specimens collected before treatment (n = 7) and after treatment (n = 14).
As our results have shown that most samples are a mixture of basal and classical cancer cells, which could imply subtype plasticity, we investigated the effect of chemotherapy on subtype distribution. Among the top differentially expressed genes was TFF3, part of the classical subtype signature7,27, which was significantly higher in naive samples than treated samples (Supplimentary Fig. 6A, Supplimentary Data 4). To validate the differences in RNA expression we performed IHC analyses of TFF3 in select corresponding tissue slides (Supplimentary Fig. 6B, C). Since a single marker might not be sufficient to characterize samples as basal or classical, we used representative genes for both Moffitt subtypes7 to create a basal and classical signature score which allowed us to place each cell on a basal-classical spectrum rather than assigning discrete phenotypes. Overall, treated samples did not show a shift towards a more basal-like expression pattern (Fig. 6A), although the treated sample P03 was distinctly more basal than any other sample (Fig. 6B). P03 was, notably, the only treated sample with poorly differentiated histology. Generally, cancer cells clustered by sample on this subtype spectrum, highlighting intertumoral heterogeneity (Fig. 6B). We saw this behavior in our confirmatory dataset (Supplimentary Fig. 6D, E), which also contained one distinctly basal treated patient; when this patient was removed, the median classical score in the treated samples was actually higher than in the untreated samples. Looking beyond Moffitt signature genes, we also measured the heterogeneity of a sample’s expression profile by calculating the correlations between every pair of its cancer cells. By this metric, treated and naive samples displayed no difference in intratumor heterogeneity, either per sample (Fig. 6C) or per cell (Fig. 6D).

A Basal and classical signature expression scatterplot of cancer cells (treated, naive and combined). B Basal and classical signature expression scatterplot of cancer cells (treated and naïve by patient, combined by treatment); sample P03 has been removed from the combined plot. C Box plot showing the median intracluster correlation of cancer cells by sample for treated (n = 6) and naive (n = 11) groups (two-sided Wilcoxon rank-sum test, p = 0.26; box plots centered around the median with hinges at 1st and 3rd quartiles and whiskers from hinge to max value or 1.5*IQR, whichever is smallest). D Density plots of cancer cell correlation within samples for treated and untreated groups. E Gene set enrichment analysis of overall cancer cells, treated and untreated.
To identify cell states, we used GSEA to check for common gene expression patterns of cancer cells in each sample. Here, cancer cells from treated samples were enriched in pathways related to angiogenesis and EMT, processes that can contribute to tumor recurrence and progression, as compared to naive samples (Fig. 6E). In the confirmatory dataset, we continued to see pathways related to angiogenesis and EMT enriched in the treated samples (Supplimentary Fig. 6F). Analysis of CAF and TAM subpopulation distributions between naive and treated groups yielded no significant changes (Supplimentary Fig. 6G).
Chemotherapy reduces expression of inhibitory checkpoint molecules and ligand-receptor interactions in PDAC
We applied CellPhoneDB40 to infer potential ligand-receptor interactions (LRIs) between malignant epithelial cells, TAM and CAF subpopulations, CD8 + and CD4 + T cells, MDSCs, and DCs in naïve and treated samples (Fig. 7A; more detailed results in Supplimentary Data 5), and found a general decrease of cell-cell interactions in the chemotherapy-treated group in our dataset (Fig. 7B). We noted an unexpectedly high volume of putative interactions between CAFs and TAMs, which is highlighted in Supplimentary Fig. 7A. Before replicating this analysis in our confirmatory dataset, we removed one sample, which had an extreme number of SPP1 + cells (Supplimentary Fig. 7B) and was producing highly unusual results. The general overview of LRIs in the confirmatory dataset matched that of our original dataset, with the decrease in LRIs between naïve and treated samples being even more prevalent across compartments (Supplimentary Fig. 7C, D).

A Heatmap displaying an overview of all inferred LRIs between displayed cell types (cancer cells, MDSCs, DCs, C1QC + TAMs, SPP1 + TAMs, iCAFs, myCAFs, CD8 + T cells, CD4 + T cells). Left: untreated samples, right: treated samples. CAF cancer-associated fibroblast, DC dendritic cell, iCAF inflammatory CAF, LRI ligand-receptor interaction, MDSC myeloid-derived suppressor cell, myCAF myofibroblastic CAF, TAM tumor-associated macrophage. B Heatmap displaying the fold change between the number of inferred LRIs in naïve and treated samples for each pair of cell types. Red indicates more inferred LRIs in naïve samples and blue indicates more inferred LRIs in treated samples. C Dot plot of CellphoneDB output (see Methods) for recruitment-associated LRIs between iCAFs/myCAFs and SPP1 + /C1QC + TAMs comparing untreated and treated samples. D Differential gene expression of checkpoint molecules in CD8 + T cells between treated and untreated samples. E Dot plot of CellphoneDB output (see Methods) for checkpoint molecule LRIs between CD8 + T cells, SPP1 + or C1QC + TAMs, and cancer cells comparing untreated and treated samples.
As CAFs are known to recruit and polarize macrophages, we checked for recruitment-associated ligand-receptor interactions between CAFs and TAMs41. There was evidence for only two ligand-receptor pair interactions in this analysis: low levels of the CSF1-CSF1R interaction that did not change with treatment, and an interaction between CXCL12 (on iCAFs but not myCAFs) and CXCR4 (on both TAM subpopulations). CXCL12-CXCR4, the strongest potential ligand-receptor interaction pair in naive samples, significantly weakened with treatment in both our dataset and the confirmatory dataset (Fig. 7C, Supplimentary Fig. 7E–H). Notably, it appears that the change in the volume of the CXCL12-CXCR4 interaction is driven by a decrease in CXCR4 in TAMs with treatment (Supplimentary Fig. 7F–H). The CXCL12-CXCR4 interaction between perivascular CAFs and TAMs has recently been linked to metastasis42 and may be one way in which conventional chemotherapy affects crosstalk in the TME.
Since checkpoint inhibitory drugs have yet to show benefit in PDAC clinical trials (except in the 1% of PDAC cases with mutations in mismatch repair genes43), we further evaluated treatment-related checkpoint molecule expression in CD8 + T cells. We found TIGIT to be the highest and most broadly expressed inhibitory checkpoint molecule, while PDCD1 was expressed at low levels and in a minority of CD8 + T cells (Fig. 7D). In addition, expression of TIGIT, PDCD1, and other checkpoint molecules was lower in CD8 + T cells of chemotherapy-treated samples than naive samples (Fig. 7D). The findings in Fig. 7D are replicated very closely in the confirmatory set (Supplimentary Fig. 7I). To further validate our findings, we used multiplex IHC to stain for CD8 + T cells, TIGIT, and PD-1 in corresponding tissue slides of treatment-naïve and treated and pancreatic samples (Supplimentary Fig. 7J). Using representative samples, we found that TIGIT was expressed in most CD8 + T cells in naive samples and to a significantly lesser extent in CD8 + T cells in treated samples (Supplementary Figure 7K). In addition, most CD8 + T cells did not show expression of PD-1 in either naive or treated samples (Supplimentary Fig. 7L). Of note, we found cells in the TME that are TIGIT positive but CD8 negative, representing expression in other immune cell types.
We also assessed whether treatment alters checkpoint molecule interactions between CD8 + T cells, TAMs, and malignant cells. We evaluated all CD8 + T cells due to the scarcity of exhausted T cells in our dataset. Moreover, CD8 + T cells can exhibit different states of exhaustion, and the larger CD8 + population is more representative of response regarding checkpoint molecule interactions with chemotherapy44. We observed more potential ligand-receptor interactions between TAMs and CD8 + T cells than between cancer cells and CD8 + T cells in both our samples (Fig. 7E; Supplimentary Fig. 7M, N) and the confirmatory dataset (Supplimentary Fig. 7O–P). Comparing naïve and treated samples, we noticed that all of the putative inhibitory ligand-receptor interactions were significant only in naive samples, while immune stimulatory interactions between TAMs and CD8 + cells were significant in both treated and naive samples (Fig. 7E). The only significant PD-1 (PDCD1) interaction occurred in naive samples, between PD-1 on C1QC + TAMs and PD-L2 (PDCD1LG2) on CD8 + T cells (Fig. 7E); this interaction was not present in the confirmatory dataset (Supplimentary Fig. 7O). We only detected three significant interactions between CD8 + T cells and cancer cells, with TIGIT-PVR being the strongest. TIGIT-PVR was also the only interaction that was still significant in treated samples, though the effect was weaker than in naive samples (Fig. 7E). Of those three, only PVR-TIGIT was found in the confirmatory dataset (Supplimentary Fig. 7O). Of note, we did not detect any PD-1 related interactions between CD8 + T cells and cancer cells. We next performed multiplex IHC to assess coexpression of TIGIT + on CD8 + T cells and PVR + on CK19 + cancer cells at the protein level. We detected both cell types expressing the ligand-receptor pair in close proximity to each other, supporting the potential for ligand-receptor interactions between those cell types (Supplimentary Fig. 7Q).
Discussion
In this study we systematically characterized the composition of human PDAC at the single-cell transcriptomic level and assessed the effects of chemotherapy on the TME. Individual samples demonstrated significant heterogeneity in their Moffitt subtypes and TME composition. Analysis of the TME revealed discrete cell subpopulations with distinct features, further highlighting the complexity of the PDAC TME. We found, and confirmed in an external dataset, that chemotherapy profoundly alters the PDAC TME and might lead to further resistance to immunotherapy by reduced inhibitory checkpoint molecule expression and interactions involving CD8 + T cells.
Consistent with previous reports on subtype heterogeneity, our initial per-sample analysis of cancer cell composition revealed a heterogeneous mixture of cells with classical or basal transcriptional signatures in most samples and a variety of subclonal growth patterns22,25,45,46. A recent study demonstrating plasticity of pancreatic cancer cells and subtype switching dependent on culture conditions in vitro also identified a new intermediate subtype state25. Although we did not observe this specific intermediate state gene signature (the number of samples in each dataset may limit the generalizability of such a signature), we did discover cancer cells in numerous patient samples that expressed both basal and classical histological markers to varying degrees. Since both subtypes exhibit different biology and metabolic features47,48, using scRNA-seq to evaluate individual subtype composition prior to therapy might help understand how these elements dictate responses to different therapeutic approaches.
In recent years, investigation of CAFs has led to the identification of distinct CAF subpopulations in human and mouse PDAC19. While iCAFs and myCAFs were present in our dataset, neither unsupervised clustering nor signature analysis revealed a defined apCAF population. Our finding is also consistent with a recent publication on single nucleus sequencing in human PDAC49. Nevertheless, a selection of iCAFs and myCAFs did co-express CD74 and HLA genes, consistent with another recent report22. Plasticity between CAF subtypes, as hypothesized by multiple groups, might explain the differences in CAF subpopulation findings19,29.
Myeloid cell analysis led to the identification of SPP1 + and C1QC + TAMs as the major TAM subpopulations in human PDAC. In our data, M1/M2 polarization did not fully match these recently described subpopulations, suggesting that M1/M2 classification does not capture the complexity of TAM subpopulations in the human TME50,51,52. Further characterization showed enrichment for EMT and a high angiogenesis score in SPP1 + TAMs, while C1QC + TAMs were enriched for antigen presentation and phagocytosis. These findings are consistent with recent scRNA-seq studies in CRC38,53. A SPP1 + high/C1QC + low TAM signature has been associated with worse overall survival in the TCGA CRC dataset, and SPP1 + TAMs have been linked to CRC liver metastases38,54. In our analysis of both TAM signatures in the TCGA PDAC dataset, TAM subtype combination did not correlate with worse overall survival. Since the TCGA PDAC dataset has only bulk transcriptomics and a limited variety of clinical stages, the role of SPP1 + and C1QC + TAMs in PDAC will require further evaluation in a larger, non-bulk RNA clinical cohort.
For a more unambiguous analysis of chemotherapy effects on the TME we compared naive and treated samples from primary tumors only. Recent studies have correlated the classical subtype with better therapy response than the more aggressive basal subtype and proposed that chemotherapy might shift the Moffitt subtype from classical to basal55,56.However, when assessing for a basal and classical multi-gene signature we found heterogeneity between tumors in regard to Moffitt subtype composition but no shift toward a basal subtype in chemotherapy-treated samples, nor a significant difference in intratumor heterogeneity. Taken together, the heterogeneity within each tumor and differences in individual gene expression imply that PDAC subtype categorization should not only rely on single marker analysis. Since individual genes might not reveal biological changes in cancer cells with treatment, we also checked for gene expression programs with GSEA and noticed a Moffitt-independent shift in enrichment of angiogenesis and EMT genes in treated samples. EMT is a hallmark feature of tumor invasion, and different EMT programs have been linked to PDAC progression, dissemination, and resistance to chemotherapy57,58,59. While the complexity of EMT makes therapeutic approaches targeting it challenging, novel approaches would be highly impactful60. Tumor angiogenesis and microvessel density have been linked to metastasis in PDAC61,62,63; while anti-angiogenic drugs have thus far not yielded clinical results, the question of how chemotherapy affects the microvasculature in PDAC warrants further investigation64.
The complexity of PDAC is not only determined by the cancer cell heterogeneity but also the intricate composition of and communication between cells in the TME. In our analysis, we found a high volume of putative ligand receptor interactions between TAMs and CAFs. There is limited data on crosstalk between CAFs and TAMs in the TME, even though these interactions likely play an important role in PDAC biology and may reveal potential targets for therapy41. We found CXCL12-CXCR4, specific to iCAFs and TAMs respectively, to be the most significant potential interaction. Notably, it has been demonstrated that CXCL12 secretion by CAFs in PDAC promotes immune evasion by T cell exclusion65, and CXCL12-CXCR4 interaction between perivascular CAFs and TAMs promotes metastasis in a breast cancer model42. In our dataset, CXCL12-CXCR4 interaction between iCAFs and TAMs was significantly lower in treated samples, exemplifying how conventional chemotherapy may alter the PDAC TME in ways we could not previously observe.
Although effective in a number of other malignancies, checkpoint inhibition therapy has performed poorly in PDAC. While the underlying biology is still being uncovered, few studies have explored checkpoint expression in human PDAC samples at the single cell level. In our analysis, exhausted CD8 + T cells constituted only 2.8% of all CD8 + T cells. This corresponds with previous studies showing PDAC to be a relatively immune-cold tumor with low PD-1 expression and might be a reason why checkpoint inhibition has failed in clinical trials for PDAC4,5,66. The strongest interaction we observed between CD8 + T cells and cancer cells was TIGIT-PVR, which significantly decreased with treatment. Recently, Steele et al. found elevated TIGIT expression levels in tumor-infiltrating CD8 + T cells compared to CD8 + T cells from normal pancreatic tissue, but no changes in PD1 between the two groups21. Functionally, Freed-Pastor et al. used a PDAC mouse model to show that the TIGIT-PVR axis is critical for maintaining immune evasion and that TIGIT blockage elicited an immune response to PDAC67. TIGIT was the most expressed inhibitory checkpoint molecule on CD8 + T cells and its expression, as with other checkpoint molecules, decreased with treatment. Lately, another study using single-nucleus sequencing in PDAC also found TIGIT overexpressed in CD8 + T cells in naive samples49. These findings and ours imply that TIGIT may be a stronger immunotherapy candidate than PD-1 in PDAC. The higher TIGIT expression in treatment-naive samples further suggests that blocking TIGIT may be more effective given as first-line monotherapy or administered concurrently with chemotherapy, and that immune blockade as second-line treatment following chemotherapy may lessen its antitumor effects. While a recent phase III trial of tiragolumab as first line therapy in small cell lung cancer was unsuccessful68, the Morpheus-Pancreatic Cancer trial (NCT03193190) is investigating the effect of TIGIT blockage in metastatic PDAC and will hopefully provide more insight into the clinical relevance of TIGIT inhibition in PDAC. Overall, our results provide an in-depth view into human PDAC composition, its TME, and the effects of chemotherapy, and may provide insight into more effective strategies to treat this recalcitrant disease.
Methods
Patient cohort and data collection
A total of 27 PDAC patients were recruited for study participation at the Perlmutter Cancer Center at New York University (NYU) Langone Health between May 1, 2020, and June 30, 2021. Informed written consent for the collection of blood, tissue, and clinical information was obtained from all patients using a study protocol approved by the NYU Langone Health Institutional Review Board. Standard clinicopathological variables including sex, age, tumor site, disease stage, type of procedure, tumor stage (TNM classification), and treatment type were collected for each patient as part of a prospective PDAC database. Clinical germline and somatic tumor analyses (Tempus xT 596 gene panel, Tempus Labs, Chicago, IL; Invitae 91 gene panel, Invitae Corporation, San Francisco, CA; FoundationOne CDx 324 gene panel, Foundation Medicine, Cambridge, MA) were performed. A supplement providing individual UMAP, H&E as well as clinical and mutational information is provided as Supplementary Data 2.
Sample acquisition and processing
Of the 27 samples, 10 were obtained following surgical resection, 7 were obtained from endoscopic ultrasound (EUS)-guided biopsies, and 10 were obtained by interventional radiology (IR)-guided biopsies from metastatic liver lesions. For EUS- and IR-guided biopsies, 2-3 cores were used for scRNA-seq analysis. The pathologic diagnosis of PDAC was confirmed in all samples by a board-certified pancreatic pathologist. Within two hours of tissue acquisition, the specimen was placed in refrigerated RPMI 1640 medium (Gibco, Cat. No. 11875101) and transported to the Center for Biospecimen Research & Development at NYU, where the sample was provided for tissue processing.
Tissue processing and single cell isolation
Tissue samples were washed in ice-cold phosphate-buffered saline (PBS; Corning, Cat. No. 21-040-CM) and grossly assessed for adequacy (solid tissue component, free of large hemorrhagic or necrotic areas). Next, the tissue was manually minced with a scalpel blade and filtered through a 40 µm mesh filter (Falcon; Cat. No. 352340) to remove any excess debris or blood. The tissue was then transferred to a sterile gentleMACS C tube (Miltenyi Biotec, Cat. No. 130096334), resuspended in 3–5 mL of Miltenyi Tumor Dissociation Kit buffer (Cat. No. 130-095-929), and mechanically minced to submillimeter particles. The suspension was then digested utilizing the gentleMACS Octo Dissociator (Miltenyi Biotec, Cat. No. 130-095-937) for 25–30 min using the “37C_h_TDK_3” program for tumor tissue. Dissociated cells were then quenched with ice-cold RPMI 1640 containing 10% fetal bovine serum (FBS; Gibco, Cat. No. 10082147) and filtered through a 40 µm mesh filter. Red blood cells were removed by treatment with 3–5 mL of ACK lysis buffer (Gibco, Cat. No. A1049201) and a subsequent washing step in RPMI 1640 with 10% FBS. Final cell counts were established with 0.4% trypan blue solution and a Countess II Automated Cell Counter (Applied Biosystems, Cat. No. A27977). The final concentration was titrated to 300–500 viable cells/µL for further analysis.
Single-cell library preparation
Single-cell suspensions were processed for 10x Genomics by the Genome Technology Center at the NYU School of Medicine per the manufacturer’s guidelines. The libraries were prepared using the following reagents: Chromium Single Cell 3′ Reagent Kits (v3)—the Single Cell 3′ Library & Gel Bead Kit v3 (PN-1000075), the Chromium Next GEM Chip G Single Cell Kit (PN-100120), and the i7 Multiplex Kit (PN-120262) (10x Genomics) —by following the Single-Cell 3′ Reagent Kits (v2) User Guide (manual part no. CG00052 Rev A) and the Single-Cell 3′ Reagent Kits (v3) User Guide (manual part no. CG000183 Rev C). DNA libraries were run using the NovaSeq 6000 system (Illumina, San Diego, CA) to achieve roughly 500 million paired-end reads per sample.
Single-cell RNA sequencing and data processing
Sequencing results were de-multiplexed and converted to FASTQ format using Illumina bcl2fastq software. The 10x Genomics Cell Ranger 5.0.1 software suite69 was used to perform sample de-multiplexing, barcode processing, and single-cell 3’ gene counting. The cDNA insert was aligned to the hg38/GRCh38 reference genome. Only confidently mapped, non-PCR duplicates with valid barcodes and unique molecular identifiers were used to generate the gene-barcode matrix. Further analysis including the identification of highly variable genes, dimensionality reduction, standard unsupervised clustering algorithms, and the discovery of differentially expressed genes was performed using Seurat70 and scooter71.
Raw data were initially filtered to remove ambient RNA using SoupX version 1.5.272; and cells were subsequently filtered to include only high-quality cells (as defined by >500 detectable genes, >1500 unique molecular identifiers, and <15% of transcripts coming from mitochondrial genes). Cells with >1% of transcripts representing erythroid genes (HBA1, HBA2, HBB, HBM, and ALAS2) were also excluded from the analysis. The data were normalized to the total expression, multiplied by a scaling factor of 10,000, and log-transformed. Likely doublets/multiplets were identified and removed using scDblFinder 1.6.073.
To account for biological and technical batch differences between individual patients and scRNA-seq libraries, the Seurat anchor-based integration method for merging datasets that identify pairwise correspondences between cell pairs across datasets74 was utilized to transform them into a shared space. The 2000 most variable genes based on standardized variance were selected for canonical correlation analysis as an initial dimensional reduction. The integration anchors were then identified based on the first 30 dimensions and used to generate a new dimensional reduction for further analysis.
To visualize the data, the dimensionality of the scaled integrated data matrix was further reduced to project the cells in two-dimensional space using principal component analysis followed by UMAP75. The 30 nearest neighbors were used to define the local neighborhood size with a minimum distance of 0.3. The resulting principal components were also used as a basis for partitioning the dataset into clusters using a smart local moving community detection algorithm76. A range of resolutions (0.1–10) was utilized to establish a sufficient number of clusters to separate known populations based on the expression of established markers.
Cell identification and clustering
Single-cell clusters were identified based on common marker genes for various cell types, the most prominent of which are listed here. Full dataset (Fig. 1C): T/NK (CD3E), epithelial (KRT19), endothelial (VWF, PECAM1), myeloid (CD68), proliferating epithelial (KRT19, MKI67), proliferating lymphoid (CD3E, MKI67), B/plasma (CD79A), proliferating myeloid (CD68, MKI67), mesenchyme (DCN), mast (KIT). Mesenchyme (Fig. 3A): myCAF (ACTA2, MMP11, COL10A1−), iCAF (C3, C7, CFD, PTGDS), pericyte (ACTA2+, RGS5+, CSPG4, NOTCH3), proliferating (MKI67), peri-islet Schwann cells (SOX10, S100B, PLP1), epithelial-like (KRT19, EPCAM), chondrocyte-like (SPP1, IBSP, MMP13). T/NK (Fig. 4A): CCR7 + CD4 + T cells (CCR7), IL7R + CD4 + T cells (IL7R), FOXP3 + CD4 + T cells (FOXP3), CXCL13 + CD4 + T cells (CXCL13), GZMH + CD8 + T cells (GZMH), GZMK + CD8 + T cells (GZMK), ITGA1 + CD8 + T cells (ITGA1), ISG15 + T cells (ISG15), GNLY + NK cells (GNLY, NCAM1), XCL1 + NK cells (XCL1, NCAM1), KIT + mast (KIT), CD79 + B/Plasma (CD79A), MKI67 + proliferating (MKI67). CD8 + T cells (Fig. 4C): GZMK + CD8 + T cells (GZMK), GZMH + CD8 + T cell (GZMH), ITGA1 + CD8 + T cells (ITGA1), SELL + CD8 + T cells (SELL), ISG15 + CD8 + T cells (ISG15), CTLA4 + CD8 + T cells (CTLA4), HSPA6 + CD8 + T cells (HSPA6), epithelial-like (KRT19), erythrocytes (HBB). Myeloid (Fig. 5): SPP1 + macrophage (SPP1, MARCO), C1QC + macrophage (C1QA, C1QB, C1QC), MDSC (S100A8, S100A9, S100A12), monocyte (FCGR3A, CDKN1C), cDC1 (CLEC9A, XCR1), cDC2 (CD1C, FCER1A), cDC3 (CCL19, CCL22, CCR7), pDC (LILRA4, PLD4), proliferating (MKI67), mast (KIT), T (CD3E), epithelial-like (KRT18, KRT19).
Identification of malignant and pancreatic epithelial cells
To detect large-scale chromosomal CNVs, InferCNV version 1.8.177 was run at a sample level with the parameters recommended for 10x Genomics data. For each sample, all epithelial cells were tested for copy number aberrations. Non-epithelial (CAF, endothelial, immune) cells were used as the reference normal set. For samples with over 1,000 non-epithelial cells, they were randomly subsampled to 1000. Additionally, 10% of non-epithelial cells were used in the observation set. We used a 101-gene window in subclustering mode with HMM enabled and a six-group clustering of observations.
Ligand-receptor interaction
CellPhoneDB v.2.1.740 was used to estimate ligand-receptor interaction. We used the statistical method with 10 iterations and no subsampling; other parameters were left as default. CPDB calculates statistical significance using permutation testing; see original paper for details.
Gene signature scores
Gene signature scores were created using Seurat’s AddModuleScore function. Lists of genes used for each score can be found in Supplementary Data 3.
Differential gene expression
Differential gene expression was analyzed using the Wilcoxon rank-sum test with Bonferroni correction that is included in Seurat’s FindMarkers function.
Gene set enrichment analysis
Gene set enrichment analysis was performed using the GSEAPreranked function of GSEA version 4.2.078. Genes were pre-ranked for each comparison using the output of our differential gene expression; first by taking the -log10(adjusted p-value) and then multiplying it by the sign of the fold change (for example, a gene with a fold change of −2.3 and an adjusted p-value of 1e-160 would have a rank metric of −160). GSEAPreranked parameters were set to the defaults. The gene chip annotation file used was Human_Gene_Symbol_with_Remapping_MSigDB.v7.4.chip, and genes were collapsed and remapped to gene symbols. Gene sets used included Hallmarks (h.all.v7.5.1), KEGG (c2.cp.kegg.v7.5.1), Reactome (c2.cp.reactome.v7.5.1), Canonical Pathways (c2.cp.v7.5.1), and GO (c5.go.v7.5.1).
Survival analysis of TCGA data
The TCGA PDAC (PAAD) clinical and gene expression data9 were downloaded, using TCGABiolinks version 3.1579, to examine the prognostic utility of specific cell subpopulation signatures. Gene expression was z-score transformed. For each signature, two distinct groups (low and high expression) were formed, corresponding to mean expression values below the 45th and above the 55th quantile values, respectively. The survival analysis was performed using the R package survminer80. Cox proportional hazards models from the R package survival81 were used to adjust for tumor stage, age, and sex.
Histology and multiplex IHC
For histopathological assessment, tumor sections were cut and slides prepared by routine processes, followed by staining with hematoxylin & eosin. For multiplex immunofluorescence and imaging, five-micron formalin fixed paraffin-embedded sections were stained with Akoya Biosciences® Opal™ multiplex automation kit reagents (Leica Cat #ARD1001EA) on a Leica BondRX® autostainer, according to the manufacturers’ instructions. In brief, slides underwent sequential epitope retrieval with Leica Biosystems Epitope Retrieval 2 solution (ER2, EDTA based, pH 9, Cat. AR9640), primary and secondary antibody incubation, and tyramide signal amplification with Opal® fluorophores as shown in Supplementary Table 3. Primary and secondary antibodies/polymers were removed during sequential epitope retrieval steps while the relevant fluorophores remained covalently attached to the antigen. Slides were counterstained with spectral DAPI (Akoya Biosciences, FP1490) and mounted with ProLong Gold Antifade (ThermoFisher Scientific, P36935). Semi-automated image acquisition was performed on a Vectra® Polaris multispectral imaging system. After whole slide scanning at 20x in the motif mode, regions of interest were selected for spectral unmixing and image processing using InForm® version 2.4.10 software from Akoya Biosciences.
IHC Quantification
Semi-automated image acquisition was performed on a Vectra® Polaris multispectral imaging system. Eleven cases were used for this quantification. After whole slide scanning at 20X the tissue was manually outlined to select fields for spectral unmixing and image analysis using InForm® version 2.6 software from Akoya Biosciences. Selection of areas was performed by a trained pathologist on H&E-stained sequential sections and copied onto the fluorescent image on Akoya Biosciences Phenochart software. A training algorithm was first used to segment images into tissue categories: “CK19 + ” or “CK19-” (to exclude areas of the image with no tissue). Tissue segmentations were reviewed for different samples during training iterations. Following tissue classification cells were segmented based on nuclear signal (DAPI). Cells were phenotyped after segmentation using inForm’s trainable algorithm based on the R glmnet package82.
Four algorithms were created to classify cells as CD8 + or ‘other’, CK19 + or ‘other’, TIGIT + or ‘other’, and PD1 + or ‘other’. Phenotypes were reviewed for different samples during training iterations. Data was exported as text containing sample names, field of acquisition coordinates, individual cell information including coordinates and identified phenotype. Each image was analyzed with all four algorithms so that every cell was classified four times. Concatenation and aggregation of all phenotyping information was performed in R using the phenoptrReports package83.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The raw scRNA-seq data generated for this project is available on GEO with accession number GSE205013. TCGA data9 is publicly available on the GDC Data Portal [https://portal.gdc.cancer.gov/]. The confirmatory dataset from Cui Zhou et al.39 is publicly available at the Human Tumor Atlas Network (HTAN) under the HTAN WUSTL Atlas [https://humantumoratlas.org/explore?selectedFilters=%5B%7B%22group%22%3A%22AtlasName%22%2C%22value%22%3A%22HTAN+WUSTL%22%7D%5D&tab=file] and on dbGaP with accession number phs002371.v2.p1. The remaining data are available within the article and its supplementary files.
Change history
03 July 2023
A Correction to this paper has been published: https://doi.org/10.1038/s41467-023-39680-2
References
Siegel, R. L., Miller, K. D., Fuchs, H. E. & Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 72, 7–33 (2022).
Rahib, L., Wehner, M. R., Matrisian, L. M. & Nead, K. T. Estimated projection of US cancer incidence and death to 2040. JAMA Netw. Open 4, e214708 (2021).
Vincent, A., Herman, J., Schulick, R., Hruban, R. H. & Goggins, M. Pancreatic cancer. Lancet 378, 607–620 (2011).
O’Reilly, E. M. et al. Durvalumab with or without tremelimumab for patients with metastatic pancreatic ductal adenocarcinoma: a phase 2 randomized clinical trial. JAMA Oncol. 5, 1431–1438 (2019).
Bear, A. S., Vonderheide, R. H. & O’Hara, M. H. Challenges and opportunities for pancreatic cancer immunotherapy. Cancer Cell 38, 788–802 (2020).
Collisson, E. A., Bailey, P., Chang, D. K. & Biankin, A. V. Molecular subtypes of pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 16, 207–220 (2019).
Moffitt, R. A. et al. Virtual microdissection identifies distinct tumor- and stroma-specific subtypes of pancreatic ductal adenocarcinoma. Nat. Genet. 47, 1168–1178 (2015).
Wood, L. D., Canto, M. I., Jaffee, E. M. & Simeone, D. M. Pancreatic cancer: pathogenesis, screening, diagnosis and treatment. Gastroenterology (2022) https://doi.org/10.1053/j.gastro.2022.03.056
Cancer Genome Atlas Research Network. Integrated genomic characterization of pancreatic ductal adenocarcinoma. Cancer Cell 32, 185–203.e13 (2017).
Ho, W. J., Jaffee, E. M. & Zheng, L. The tumour microenvironment in pancreatic cancer - clinical challenges and opportunities. Nat. Rev. Clin. Oncol. 17, 527–540 (2020).
Beatty, G. L., Werba, G., Lyssiotis, C. A. & Simeone, D. M. The biological underpinnings of therapeutic resistance in pancreatic cancer. Genes Dev. 35, 940–962, https://doi.org/10.1101/gad.348523.121 (2021).
Moncada, R. et al. Integrating microarray-based spatial transcriptomics and single-cell RNA-seq reveals tissue architecture in pancreatic ductal adenocarcinomas. Nat. Biotechnol. 38, 333–342 (2020).
Azizi, E. et al. Single-cell map of diverse immune phenotypes in the breast tumor microenvironment. Cell 174, 1293–1308.e36 (2018).
Zhang, Y. et al. Single-cell analyses reveal key immune cell subsets associated with response to PD-L1 blockade in triple-negative breast cancer. Cancer Cell 39, 1578–1593.e8 (2021).
Chan, J. M. et al. Signatures of plasticity, metastasis, and immunosuppression in an atlas of human small cell lung cancer. Cancer Cell 39, 1479–1496.e18, https://doi.org/10.1016/j.ccell.2021.09.008 (2021).
Wu, H. et al. Single-cell transcriptomics of a human kidney allograft biopsy specimen defines a diverse inflammatory response. J. Am. Soc. Nephrol. 29, 2069–2080 (2018).
Kim, N. et al. Single-cell RNA sequencing demonstrates the molecular and cellular reprogramming of metastatic lung adenocarcinoma. Nat. Commun. 11, 2285 (2020).
Peng, J. et al. Single-cell RNA-seq highlights intra-tumoral heterogeneity and malignant progression in pancreatic ductal adenocarcinoma. Cell Res. 29, 725–738, https://doi.org/10.1038/s41422-019-0195-y (2019).
Elyada, E. et al. Cross-species single-cell analysis of pancreatic ductal adenocarcinoma reveals antigen-presenting cancer-associated fibroblasts. Cancer Disco. 9, 1102–1123 (2019).
Wang, Y. et al. Single-cell analysis of pancreatic ductal adenocarcinoma identifies a novel fibroblast subtype associated with poor prognosis but better immunotherapy response. Cell Disco. 7, 36 (2021).
Steele, N. G. et al. Multimodal mapping of the tumor and peripheral blood immune landscape in human pancreatic cancer. Nat. Cancer 1, 1097–1112 (2020).
Lee, J. J. et al. Elucidation of tumor-stromal heterogeneity and the ligand-receptor interactome by single-cell transcriptomics in real-world pancreatic cancer biopsies. Clin. Cancer Res 27, 5912–5921 (2021).
Abou Khouzam, R. et al. An eight-gene hypoxia signature predicts survival in pancreatic cancer and is associated with an immunosuppressed tumor microenvironment. Front. Immunol. 12, 680435 (2021).
O’Flanagan, C. H. et al. Dissociation of solid tumor tissues with cold active protease for single-cell RNA-seq minimizes conserved collagenase-associated stress responses. Genome Biol. 20, 1–13 (2019).
Raghavan, S. et al. Microenvironment drives cell state, plasticity, and drug response in pancreatic cancer. Cell 184, 6119–6137.e26 (2021).
Puleo, F. et al. Stratification of pancreatic ductal adenocarcinomas based on tumor and microenvironment features. Gastroenterology 155, 1999–2013.e3 (2018).
Collisson, E. A. et al. Subtypes of pancreatic ductal adenocarcinoma and their differing responses to therapy. Nat. Med. 17, 500–503 (2011).
Bailey, P. et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 531, 47–52 (2016).
Öhlund, D. et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J. Exp. Med 214, 579–596 (2017).
Puram, S. V. et al. Single-cell transcriptomic analysis of primary and metastatic tumor ecosystems in head and neck cancer. Cell 171, 1611–1624.e24 (2017).
Yang, N., Mosher, R., Seo, S., Beebe, D. & Friedl, A. Syndecan-1 in breast cancer stroma fibroblasts regulates extracellular matrix fiber organization and carcinoma cell motility. Am. J. Pathol. 178, 325–335 (2011).
Cheng, H.-W. et al. CCL19-producing fibroblastic stromal cells restrain lung carcinoma growth by promoting local antitumor T-cell responses. J. Allergy Clin. Immunol. 142, 1257–1271.e4 (2018).
Crinier, A. et al. High-dimensional single-cell analysis identifies organ-specific signatures and conserved nk cell subsets in humans and mice. Immunity 49, 971–986.e5 (2018).
de Andrade, L. F. et al. Discovery of specialized NK cell populations infiltrating human melanoma metastases. JCI Insight 4, e133103 (2019).
Li, H. et al. Dysfunctional CD8 T cells form a proliferative, dynamically regulated compartment within human melanoma. Cell 176, 775–789.e18 (2019).
Kwak, T. et al. Distinct populations of immune-suppressive macrophages differentiate from monocytic myeloid-derived suppressor cells in cancer. Cell Rep. 33, 108571 (2020).
House, I. G. et al. Macrophage-derived CXCL9 and CXCL10 are required for antitumor immune responses following immune checkpoint blockade. Clin. Cancer Res. 26, 487–504,https://doi.org/10.1158/1078-0432.ccr-19-1868 (2019).
Zhang, L. et al. Single-cell analyses inform mechanisms of myeloid-targeted therapies in colon cancer. Cell 181, 442–459.e29 (2020).
Cui Zhou, D. et al. Spatially restricted drivers and transitional cell populations cooperate with the microenvironment in untreated and chemo-resistant pancreatic cancer. Nat. Genet. 54, 1390–1405 (2022).
Efremova, M., Vento-Tormo, M., Teichmann, S. A. & Vento-Tormo, R. CellPhoneDB: inferring cell-cell communication from combined expression of multi-subunit ligand-receptor complexes. Nat. Protoc. 15, 1484–1506 (2020).
Gunaydin, G. CAFs interacting with TAMs in tumor microenvironment to enhance tumorigenesis and immune evasion. Front. Oncol. 11, 668349 (2021).
Arwert, E. N. et al. A unidirectional transition from migratory to perivascular macrophage is required for tumor cell intravasation. Cell Rep. 23, 1239–1248 (2018).
Hu, Z. I. et al. Evaluating mismatch repair deficiency in pancreatic adenocarcinoma: challenges and recommendations. Clin. Cancer Res 24, 1326–1336 (2018).
Beltra, J.-C. et al. Developmental relationships of four exhausted CD8 T cell subsets reveals underlying transcriptional and epigenetic landscape control mechanisms. Immunity 52, 825–841.e8 (2020).
Chan-Seng-Yue, M. et al. Transcription phenotypes of pancreatic cancer are driven by genomic events during tumor evolution. Nat. Genet. 52, 231–240 (2020).
Grünwald, B. T. et al. Spatially confined sub-tumor microenvironments in pancreatic cancer. Cell 184, 5577–5592.e18 (2021).
Espiau, R. P., Courtois, S., Parejo, A. B. & Sancho, P. Molecular and metabolic subtypes correspondence for pancreatic ductal adenocarcinoma classification. J. Clin. Med. Res. 9, 4128 (2020).
O’Kane, G. M. et al. GATA6 expression distinguishes classical and basal-like subtypes in advanced pancreatic cancer. Clin. Cancer Res 26, 4901–4910 (2020).
Hwang, W. L. et al. Single-nucleus and spatial transcriptome profiling of pancreatic cancer identifies multicellular dynamics associated with neoadjuvant treatment. Nat. Genet. 54, 1178–1191 (2022).
Mills, C. D., Kincaid, K., Alt, J. M., Heilman, M. J. & Hill, A. M. M-1/M-2 macrophages and the Th1/Th2 paradigm. J. Immunol. 164, 6166–6173 (2000).
Anderson, C. F. & Mosser, D. M. A novel phenotype for an activated macrophage: the type 2 activated macrophage. J. Leukoc. Biol. 72, 101–106 (2002).
Italiani, P. & Boraschi, D. From monocytes to M1/M2 macrophages: phenotypical vs. functional differentiation. Front. Immunol. 5, 514 (2014).
Cheng, S. et al. A pan-cancer single-cell transcriptional atlas of tumor infiltrating myeloid cells. Cell 184, 792–809.e23 (2021).
Liu, Y. et al. Immune phenotypic linkage between colorectal cancer and liver metastasis. Cancer Cell 40, 424–437.e5 (2022).
Aung, K. L. et al. Genomics-driven precision medicine for advanced pancreatic cancer: early results from the COMPASS trial. Clin. Cancer Res 24, 1344–1354 (2018).
Porter, R. L. et al. Epithelial to mesenchymal plasticity and differential response to therapies in pancreatic ductal adenocarcinoma. Proc. Natl. Acad. Sci. USA (2019) https://doi.org/10.1073/pnas.1914915116
Aiello, N. M. et al. EMT subtype influences epithelial plasticity and mode of cell migration. Dev. Cell 45, 681–695.e4 (2018).
Arumugam, T. et al. Epithelial to mesenchymal transition contributes to drug resistance in pancreatic cancer. Cancer Res 69, 5820–5828 (2009).
Rhim, A. D. et al. EMT and dissemination precede pancreatic tumor formation. Cell 148, 349–361 (2012).
Bulle, A. & Lim, K.-H. Beyond just a tight fortress: contribution of stroma to epithelial-mesenchymal transition in pancreatic cancer. Signal Transduct. Target Ther. 5, 249 (2020).
Bielenberg, D. R. & Zetter, B. R. The contribution of angiogenesis to the process of metastasis. Cancer J. 21, 267–273 (2015).
Hanahan, D. & Weinberg, R. A. Hallmarks of cancer: the next generation. Cell 144, 646–674 (2011).
Takagi, K., Takada, T. & Amano, H. A high peripheral microvessel density count correlates with a poor prognosis in pancreatic cancer. J. Gastroenterol. 40, 402–408 (2005).
Annese, T., Tamma, R., Ruggieri, S. & Ribatti, D. Angiogenesis in pancreatic cancer: pre-clinical and clinical studies. Cancers 11, 381 (2019).
Feig, C. et al. Targeting CXCL12 from FAP-expressing carcinoma-associated fibroblasts synergizes with anti-PD-L1 immunotherapy in pancreatic cancer. Proc. Natl Acad. Sci. USA 110, 20212–20217 (2013).
Vonderheide, R. H. The immune revolution: a case for priming, not checkpoint. Cancer Cell 33, 563–569 (2018).
Freed-Pastor, W. A. et al. The CD155/TIGIT axis promotes and maintains immune evasion in neoantigen-expressing pancreatic cancer. Cancer Cell 39, 1342–1360.e14 (2021).
Mullard, A. Roche’s anti-TIGIT drug suffers a phase III cancer setback. Nat. Rev. Drug Discov. 21, 327 (2022).
Zheng, G. X. Y. et al. Massively parallel digital transcriptional profiling of single cells. Nat. Commun. 8, 14049 (2017).
Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420 (2018).
Dolgalev, I. & Yeaton, A. Scooter: streamlined scRNA-seq analysis pipeline. (2021).
Young, M. D. & Behjati, S. SoupX removes ambient RNA contamination from droplet-based single-cell RNA sequencing data. Gigascience 9, 151 (2020).
Germain, P.-L., Lun, A., Macnair, W. & Robinson, M. D. Doublet identification in single-cell sequencing data using scDblFinder. F1000Research 10, 979, https://doi.org/10.12688/f1000research.73600.1 (2021).
Stuart, T. et al. Comprehensive Integration of Single-Cell Data. Cell 177, 1888–1902.e21, https://doi.org/10.1016/j.cell.2019.05.031 (2019).
Becht, E. et al. Dimensionality reduction for visualizing single-cell data using UMAP. Nat. Biotechnol. (2018) https://doi.org/10.1038/nbt.4314.
Waltman, L. & van Eck, N. J. A smart local moving algorithm for large-scale modularity-based community detection. Eur. Phys. J. B 86, 471 (2013).
Tickle, T., Tirosh, I., Georgescu, C., Brown, M. & Haas, B. inferCNV of the Trinity CTAT Project. (Bioconductor, 2019). https://doi.org/10.18129/B9.BIOC.INFERCNV
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Colaprico, A. et al. TCGAbiolinks: an R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. vol. 44, e71–e71, https://doi.org/10.1093/nar/gkv1507 (2016).
Kassambara, A., Kosinski, M. & Biecek, P. survminer. (2021).
Therneau, T. M. A Package for Survival Analysis in R. (2022). https://CRAN.R-project.org/package=survival
Friedman, J., Hastie, T. & Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 33, 1–22 (2010).
Johnson, K. S. PhenoptrReports: Create reports using Phenoptics data. (2022). https://akoyabio.github.io/phenoptrReports/
Acknowledgements
We thank Valeria Mezzano Robinson, Shanmugapriya Selvaraj and Branka Brukner Dabovic from the Experimental Pathology Core at NYU Langone Health for support with multiplex IHC staining and imaging. We thank Peter Meyn and Yutong Zhang from the Genome Technology Center Core at NYU Langone Health. We thank the Center for Biospecimen Research and Development at NYU Langone Health for support with tissue acquisition. These shared resources are partially supported by the Cancer Center Support Grant P30CA016087 at the Laura and Isaac Perlmutter Cancer Center. We thank the Ding Lab at WUSTL for providing us with their data. This project was supported by NIH R01CA245005 (DMS) and a research scholar grant from the German Research Foundation (WE7047/1-1; DW). This investigation was also supported by the National Institutes of Health under Ruth L. Kirschstein National Research Service Award T32CA193111. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIH.
Author information
Authors and Affiliations
Contributions
G.W., D.W., E.A.K., I.D., A.T. and D.M.S. developed the study concept and were responsible for the study design. D.W., E.A.K. and G.W. wrote the manuscript, which was then edited by all co-authors. D.M.S., T.G., X.J. and C.H. helped acquire human biospecimens. G.W., D.W., E.Z., L.W., G.O. and A.H. processed samples for scSeq. D.W., E.A.K., G.W., I.D. and D.K. analyzed the scRNA-Seq data. E.A.K., I.D. and D.K. were responsible for the bioinformatics analysis pipelines. X.J., G.W., E.Z., H.B.L., and D.W. collected patient information and maintained the databases. C.H., C.L., D.W., L.W., G.W. and D.M.S. analyzed the H&E and multiplex IHC. S.D., T.G., N.B., L.K. and P.O. provided study guidance and feedback on the manuscript. M.H.S., A.W.L. and T.H.W. assisted with study design and analysis. D.M.S. and A.T. directed the study. All co-authors approved the final version of the manuscript before submission.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Werba, G., Weissinger, D., Kawaler, E.A. et al. Single-cell RNA sequencing reveals the effects of chemotherapy on human pancreatic adenocarcinoma and its tumor microenvironment. Nat Commun 14, 797 (2023). https://doi.org/10.1038/s41467-023-36296-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-023-36296-4
This article is cited by
-
Comparative immune profiling of pancreatic ductal adenocarcinoma progression among South African patients
BMC Cancer (2024)
-
CD105+CAF-derived exosomes CircAMPK1 promotes pancreatic cancer progression by activating autophagy
Experimental Hematology & Oncology (2024)
-
Tumor-associated neutrophils upregulate Nectin2 expression, creating the immunosuppressive microenvironment in pancreatic ductal adenocarcinoma
Journal of Experimental & Clinical Cancer Research (2024)
-
Tumor immune microenvironment-based therapies in pancreatic ductal adenocarcinoma: time to update the concept
Journal of Experimental & Clinical Cancer Research (2024)
-
The nuclear factor ID3 endows macrophages with a potent anti-tumour activity
Nature (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
