
Abstract
Sharks and rays are key functional components of coral reef ecosystems, yet many populations of a few species exhibit signs of depletion and local extinctions. The question is whether these declines forewarn of a global extinction crisis. We use IUCN Red List to quantify the status, trajectory, and threats to all coral reef sharks and rays worldwide. Here, we show that nearly two-thirds (59%) of the 134 coral-reef associated shark and ray species are threatened with extinction. Alongside marine mammals, sharks and rays are among the most threatened groups found on coral reefs. Overfishing is the main cause of elevated extinction risk, compounded by climate change and habitat degradation. Risk is greatest for species that are larger-bodied (less resilient and higher trophic level), widely distributed across several national jurisdictions (subject to a patchwork of management), and in nations with greater fishing pressure and weaker governance. Population declines have occurred over more than half a century, with greatest declines prior to 2005. Immediate action through local protections, combined with broad-scale fisheries management and Marine Protected Areas, is required to avoid extinctions and the loss of critical ecosystem function condemning reefs to a loss of shark and ray biodiversity and ecosystem services, limiting livelihoods and food security.
Similar content being viewed by others

Introduction
Coral reefs are amongst the most diverse ecosystems on the planet, harbouring more than one-third of the ocean’s fish species1. Yet, they face some of the most intense and widespread threats of any ecosystem and are increasingly under threat globally due to climate change, poor water quality, and coastal development2,3. Overfishing, however, is the most immediate direct and indirect threat to most reefs and the cascading consequences for reef ecology and dependent coastal communities are only now being revealed. Coral reef fisheries directly support the livelihoods and food security of over half a billion people4,5. In turn, the footprint of coral reef fisheries is determined largely by proximity and size of human communities to reefs6 and their reliance on these ecosystems4. However, this human footprint far exceeds the local reef productivity across many of the world’s reefs7. These intense fisheries eliminate larger-bodied size classes and high trophic level fishes, particularly sharks and rays8. Such functional losses distort ecological pyramids through mesopredator release9, leading to ecosystem disruptions8 and cascading changes down coral reef food chains that lead to declines in functionally important herbivores10.
Chondrichthyans (hereafter, sharks and rays) are a phylogenetically diverse and ecologically important megafaunal lineage on coral reefs. There are 30 families, 59 genera, and 134 species of reef-associated sharks and rays11,12, each with varying degrees of coral-reef association; from residents that spend their entire lives at one or a few reefs (e.g. Halmahera epaulette shark, Hemiscyllium halmahera12), partial residents that spend most of their time on reefs (e.g. Australian weasel shark, Hemigaleus australiensis13), and those transient passing through reef habitats (e.g. Javanese cownose ray, Rhinoptera javanica11). These species fill a range of ecological niches, including: filter feeders, benthic invertivores, resident piscivorous mesopredators, transient apex predators, and more14,15. As highly mobile predators, some species are important nutrient vectors and controllers of primary production16, while others influence primary production through fear-induced trophic cascades17. Here are four examples of nutrient vectoring and cycling. First, grey reef shark (Carcharhinus amblyrhynchos) derives >80% of its diet from pelagic fishes then transfers these nutrients onto reefs16. Similarly, reef manta rays (Mobula alfredi) feed primarily on pelagic zooplankton and transfer these nutrients onto the reef during the day, acting as vectors for horizontal and vertical nutrient transport18. Stingrays are important ecosystem engineers, bioturbating large stretches of soft sediment while feeding or burying themselves in sand19,20. Finally, larger transient apex predators, like the tiger shark (Galeocerdo cuvier) mediate ecosystem structure and function of seagrass beds through fear-induced changes in grazing behaviour of turtles and dugongs17,21, which may be a stronger effect than direct predation22.
The warning signs of reef shark depletion first became apparent from space-for-time comparisons of recreational diver surveys of the Caribbean23, comparisons across various levels of protection on the Great Barrier Reef24, and in the remote atolls of the Indian and Pacific Oceans25,26. The most compelling evidence of widespread depletion comes from Baited Remote Underwater Video (BRUV) surveys of all shark species across 371 reefs, which found that a subset of sharks are functionally extinct at 20% of sites27. These conclusions relate to only a small fraction of reef shark and ray diversity; the survey recorded 59 species but 93% of the observations were of only the 10 most common sharks. The key question remains whether these local and regional surveys of the most common sharks forewarn of a global loss of coral reef shark and ray biodiversity and associated diminution of ecosystem services.
Here, we answer this question with the recently completed International Union for Conservation of Nature (IUCN) Red List of Threatened Species reassessment of sharks and rays28. Specifically, we provide a comprehensive assessment of the extinction risk of all 134 coral reef associated shark and ray species and compare their status with all other 4918 coral reef species assessed using IUCN Red List Categories and Criteria. Second, we identify the key threatening processes using (i) the IUCN threat classification scheme28, combined with (ii) species-level29 and (iii) national-level trait-based vulnerability analyses30. Third, we develop an IUCN Red List Index to track the progress toward international biodiversity targets over the past half century31.
Results
Shark and ray extinction risk
We estimate that two-thirds of coral reef sharks and rays are threatened with extinction, based on the observed number of threatened species combined with the estimated number of Data Deficient (DD) species that are likely to be threatened (Critically Endangered (CR), Endangered (EN), or Vulnerable (VU)). Of the 134 species assessed, 79 (59.0%) are in a threatened category with 14 (10.5%) CR, 24 (17.9%) EN, and 41 (30.6%) VU. A further 18 (13.4%) species are Near Threatened (NT), 28 (20.9%) Least Concern (LC), and nine are DD (6.7%). Assuming that DD species are threatened in the same proportion as non-DD species28, we estimate that 59.0%–65.7% of coral reef sharks and rays are threatened (mean 63.2%; n = 134; Fig. 1). This is the first assessment for 12.7% (n = 17 of 134) of sharks and rays, and these species arose from recent changes in taxonomic concept (Supplementary Data 1). Overall, most species (70.1%, n = 94), whether threatened or otherwise, exhibit a decreasing population trend; only 10.4% (n = 14) are stable and the population trend is unknown for 18.7% (n = 25). Only a single species - the bluespotted ribbontail ray (Taeniura lymma), is increasing globally, based on BRUV abundance estimates from across its range32. Overall, rays (range 64.3%–72.9%; n = 70) are more threatened than sharks (53.1%–57.8%; n = 64).

Red lines indicate the best estimate of threat assuming all Data Deficient (DD, grey shade) species faced a similar level of threat to the data-sufficient species in the taxon. Extinct (EX), Critically Endangered (CR), Endangered (EN), Vulnerable (VU), Near Threatened (NT), and Least Concern (LC). Source data are provided as a Source Data file.
Threat comparison to other taxa
Sharks and rays are the second most threatened group of the 4918 species found on coral reefs that have been assessed against Red List Categories and Criteria. Only marine mammals, comprising eight species of dolphins and sea-cows, are more threatened (Fig. 1). Eight taxa range from 4–31% of species in threatened categories, compared to 59% of sharks and rays, while the other three classes (Bivalvia, Cephalopoda, and Malacostraca) have no threatened species, based on those currently evaluated (Fig. 1). The high level of threat for coral reef sharks and rays is almost twice that of the whole Class Chondrichthyes, for which the estimated threat level is 38.5% for all 1199 species28. Coral reef sharks and rays are more at risk than all coastal sharks and rays, in which just over half (50.9%, n = 296 of 582)28 are threatened, but less threatened than oceanic pelagic sharks and rays (77.4%, n = 24 of 30)33.
Overfishing is the main threat
Three lines of evidence support widespread overfishing as the principal cause of the high threat level of coral reef sharks and rays: (1) IUCN threat classification, (2) species trait-based analysis, and (3) nation trait-based analysis. For all species with available threat information (100%, n = 132) fishing was identified as a threat mainly due to unintentional catch in both small and large-scale fisheries (Fig. 2a). Fishing was the sole threat for 75 species (57%). Targeted fishing was a threat for over two-thirds of species (68%) and unintentional catch was a threat for almost all species (98%) (Supplementary Table 1). Other threats that act in conjunction with fishing include: climate change (36% of species; n = 47 of 132), habitat loss and degradation [36% total; due to residential and commercial development (29%), agriculture and aquaculture (8%), and human disturbance (8%)], and pollution (9%). Uses of reef sharks and rays were available for 109 species. They are commonly used for human consumption (98%, n = 107 of 109) and body parts are fashioned into apparel/accessories (e.g. leather from ray skin; 23%); sharks are predominantly traded internationally for their fins and meat, and rays for their meat and skins, which are turned into leather34,35. Other common uses include aquarium display (27%), food for animals (19%), and medicines (11%).

a IUCN Threat classification for coral reef species. Percent of coral reef shark and ray species in each IUCN category by b residency patterns, and c trophic positions. The effects of d maximum body size, e geographic range size (indexed by number of nations a species occurs in), and (f) lower depth limit on the probability that a data-sufficient reef shark or ray species is listed as either Critically Endangered (CR), Endangered (EN), Vulnerable (VU), Near Threatened (NT), or Least Concern (LC) based on cumulative link mixed-effects models. Source data are provided as a Source Data file.
Species traits explain extinction risk
Migration behaviours and trophic level were important in explaining risk. Transient species had the highest overall threat level (76%), followed by partial residents (67%), while resident species had the lowest threat (44%; Fig. 2b). Resident species, including mainly benthic sharks and rays, often hide within the structures on a coral reef and are not easily caught by fishing gears; as opposed to partial residents or transient species, which transit through soft-bottom habitats which are comparatively easy locations to operate trawls and gillnets effectively36,37. Partial residents did have a higher proportion of Critically Endangered species than the other two groups, possibly due to high intrinsic sensitivity and small geographic range. These CR partial resident species tended to be larger bodied with long generation lengths or occurred in fewer than five nations. The filter feeders (low trophic position) were threatened, likely due to other attributes including their transient nature and large size. Overall, however, the higher trophic level species were significantly more threatened, based on an ordinal model (z-value(df=120)=2.75, p = 0.006) (Fig. 2c; Supplementary Data 2 and Supplementary Table 2).
Our results demonstrate that extinction risk is related to intrinsic life history sensitivity and indirect measures of the relative exposure to fishing mortality. The top-ranking ordinal logistic regression model revealed risk was greater in larger-bodied species (z-value(120)=4.35, p < 0.0001, coefficient estimate=2.16, 95% confidence interval (CI) = 1.17, 3.16), those occurring in a greater number of nations (z-value(120)=2.53, p = 0.012, coef.=1.24, 95% CI = 0.26, 2.23) and threat probability decreased slightly for species that occurred to depths greater than 100 m, suggesting depth refuge from threats (z-value(120)=−3.16, p = 0.002, coef.=−1.25, 95% CI = −2.04, −0.46; Fig. 2d–f and Supplementary Fig. 1). Relatively few non-threatened species had a maximum body size greater than 200 cm (24%, n = 13 of 55), while most (87%, n = 20/23) species over 300 cm maximum body size were Endangered or Critically Endangered. Geographic range varied from 1270 to 171,329,721 km2 and the number of nations a species occurred in varied from one to 148. The more nations a species occurs in, the more fisheries management regimes it may encounter. Species found in 10 or fewer nations had a relatively low risk (43%, n = 29/68) compared to the high risk faced by widespread species found in 50 or more nations (95%, n = 21/22). Residency type and trophic level traits significantly contributed to the status of sharks and rays on coral reefs, however, correlation between traits and maximum linear dimension (measured either as total length or disc width, see Methods) or number of nations excluded it from the final model (Supplementary Table 3 and Supplementary Fig. 2).
Species with larger maximum body sizes often have lower intrinsic rates of population growth, less capacity to withstand fishing mortality, and are typically the most commercially valuable38,39. However, the connections to extinction risk and attribution to overfishing are complicated because larger-bodied species also tend to have larger geographic ranges40. Yet, larger geographic ranges might not reduce extinction risk because mobile wide-ranging species transit through more jurisdictions and spend less time in protected areas, thus increasing their exposure to a wider range of fisheries and more fragmented management41. Threat level is determined by the interaction of intrinsic sensitivity, such as body size, with a threatening process, which we indexed with depth and geographic range as a measure of the accessibility of a species to fisheries and fragmented management29.
National attributes also explain extinction risk
We found that the national attributes associated with variations in extinction risk differed between coral reef sharks and rays. For both sharks and rays, the national percentage of globally threatened species was associated mainly with abiotic factors related to population size [sharks: 50.2%, rays 46.6% of the summed average variable importance (AVI)], followed by fishing pressure (27.1% and 43.5%, respectively), and management capacity (22.6% and 9.9%, respectively; Fig. 3a, b; Supplementary Notes; Supplementary Fig. 3).

Variable importance for explaining proportion of threatened species of all covariates across 1,000 bootstrapped boosted regression tree models for a sharks and b rays. The marginal effect c–n of each variable on the proportion of threatened species for rays and sharks ordered by variable importance. Explanatory variables were all scaled and centred around zero for presentation purposes; raw values were used in the analysis. Explanatory variables include: c sea surface temperature, d catch-per-unit-effort (CPUE), e shelf area, f primary production, g coastal population, h Gross Domestic Product (GDP), i marine protein consumption, j World Governance Index (WGI), k fishing effort, l catch, m sightings-per-unit-effort (SPUE), and n Human Development Index (HDI).
Sea surface temperature had the highest overall association with shark and ray extinction risk, with a greater importance for rays (32.4% AVI) than sharks (19.8% AVI; Fig. 3c), possibly indicating a latitudinal effect, where there are fewer threatened species in more tropical waters. Primary production was the second most important variable for sharks with a decreased extinction risk with increasing primary production (15.3% AVI), possibly due to increased population sizes resulting from carrying capacity and prey availability (Fig. 3f)26. National catch-per-unit-effort (CPUE) of reef-associated species was the second most important variable for rays (19.2% AVI) such that at very low CPUE, rays had a lower extinction risk and a slight increase in CPUE was associated with a rapid increase in extinction risk (Fig. 3d). The extinction risk of sharks was greatest in nations with larger coastal human populations, and this was the most important predictor for sharks from the fishing pressure variables (12.3% AVI), although it had little importance for rays (2.8% AVI; Fig. 3g). The level of marine protein consumption was moderately important for sharks and rays (9.1 and 6.4%, respectively), but had a counterintuitive relationship with their extinction risk; shark extinction risk decreased with an increase in marine protein consumption, whereas ray risk increased (Fig. 3i). World Governance Index (WGI) and Human Development Index (HDI) did not have strong influence on reef shark or ray extinction risk (Fig. 3j, n), indicating that even nations with higher governance capacity are not always translating this into conservation success for sharks and rays.
Where are reef species threatened?
Separately, both shark and ray richness on coral reefs are highest in northern Australia and Southeast Asia, with up to 26 and 30 species per 23,322 km² grid cell, respectively (Fig. 4a, e; Supplementary Fig. 4). The highest proportion of threatened species, however, differs such that sharks have a higher threat proportion in the western Atlantic (Fig. 4c) and rays had the highest proportion threatened throughout Asia and southeast Africa, with over 75% of ray species threatened in these areas (Fig. 4g). Species’ status are applied homogeneously throughout their entire range, despite known differences in status among nations of occurrence. Therefore, the percent of species threatened appears high in the Pacific Islands and other remote locations despite studies showing that these locations often have healthy populations of sharks26,27. Indeed, a beta regression model proved the proportion of reef sharks and rays threatened did not have a significant relationship to local relative abundance based on the results in MacNeil et al. (2020), which included shark and ray surveys on coral reefs in 52 nations (z-value(51)=−0.914, p = 0.361; Supplementary Fig. 5). Locations like the Pacific Islands, with higher abundances of reef sharks and rays act as refuges for threatened, widespread species and are important for regional and global conservation efforts (purple quadrant, Supplementary Fig. 5). Other nations with low abundances and high threat should be the focus for conservation capacity building27,42 (yellow and red quadrants, Supplementary Fig. 5). Thus, the maps should not be used in isolation to identify locations for conservation action, but rather used with information on local abundance (e.g., MacNeil et al. 2020).

a, e species richness by group. b, f number of threatened species by group. c, g threatened species as a proportion of total richness (for cells with >5 species) by species group. a–d sharks and e–h rays. Scale bar: 5000 km. d, h histograms represent the number of grid cells containing different percentages of threatened species.
Red list index
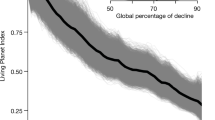
The Red List Index (RLI) is a measure of extinction status across a group of species, where a value of 1 indicates all species are Least Concern, and a value of 0 indicating all species are extinct. Risk of extinction has substantially increased since 1970 for the 125 coral reef shark and ray species included in this study that were not assessed as DD. The RLI of all coral reef sharks and rays declined from a retrospective estimate of 0.95 in 1970 to 0.64 in 2021. Coral reef sharks and rays currently have a lower RLI than the whole Class Chondrichthyes (RLI = 0.77, n = 1199) (Fig. 5). Reef rays (RLI2021 = 0.60) are more at risk of extinction than reef sharks (RLI2021 = 0.67), but their RLI trends are similar, with sharks and rays declining faster between 1980 and 2005 and slightly less so between 2005 and 2021 (Fig. 5). Most of the RLI decline for reef sharks and rays occurred prior to the 2005 assessment, likely due to a boom in coral reef fisheries in the 1960s and 70 s and collapse by the early 2000s7,43. As a result, regions like Southeast Asia, where the highest species richness occurs, were already experiencing overfishing by the 1980s43. The 2021 reef shark and ray RLI value is lower than any other coral reef taxa, including mammals, which have a higher percent of threatened species (Fig. 1).

Global Red List index (RLI) for the 125 data-sufficient reef sharks and rays combined, and each separately, estimated in 1970, 1980, 2005, and 2021, and compared to the RLI for all shark and ray species. An RLI value of 1 indicates that all species in the taxa are Least Concern and an RLI value of 0 indicates that all species are Extinct. Source data are provided as a Source Data file.
Discussion
We show that the global extinction risk of sharks and rays, as a percentage of threatened species, on coral reefs is almost double that of all 1199 sharks and rays28. Overfishing is the major threat reported and the main cause for population declines, causing dramatic declines in a very short space of time (i.e., the past 50 years). This work builds on previous work by MacNeil et al. (2020)27 showing that a smaller number of common shark species were functionally extinct on approximately 20% of surveyed coral reefs. Our analysis extends this study by including the complete list of coral reef sharks and rays, and by demonstrating that rays, as well as sharks, have high levels of extinction risk. With many threatened species, including 14 that are Critically Endangered, it is likely that the number of reefs where sharks and rays are functionally extinct will increase unless urgent action is taken to address threats. With sharks and rays playing important functional roles in coral reef ecosystems14, and their presence or absence affecting both the abundance and behaviour of other species9, there will be growing ecological consequences for coral reefs, many of which will be hard or impossible to reverse7,15.
To achieve recovery, fishing mortality from subsistence, artisanal, and industrial fisheries needs to be controlled, with the best way forward being science-based fisheries management (both traditional and ‘western’) with strong enforcement44,45 and increased use of, well-implemented and enforced, marine protected areas (MPAs)46. Additionally, due to the large spatial distribution of many coral reef species, regional level management should be implemented to address species’ status differences between neighbouring countries. For example, in Central and South America, four nations have created the Eastern Tropical Pacific Marine Corridor (CMAR) to preserve biodiversity through protecting marine corridors, which will benefit migratory and transient species47. Despite climate change being a major global threat to coral reefs2, and a threat to several coral reef shark and ray species48, it is not yet the most immediate threat that needs to be addressed to reverse the declines in shark and ray populations. In particular, the use of MPAs to successfully recover coral reef shark populations has been demonstrated both in theory49 and in practice24,50. The specific implementation of management, either through fisheries regulations or MPAs, depends on the context in which it occurs (e.g., Mizrahi et al. 201951). In many developing nations, where coral reef fisheries play important food security and livelihood roles, management responses will be different to those in developed nations where alternative livelihoods and protein sources are available45. In addition to fisheries management and MPA implementation, the following recommendations are required to ensure long-term survival of coral reef sharks and rays: (1) capacity development to enable implementation of conservation actions, including fisheries monitoring and enforcement42 and through development of Sustainable Development Goals and Convention on Biological Diversity targets (IPBES 2019)52,53; (2) improved education and diversification of rural livelihoods in nations with overexploited reefs to reduce fishing pressure on threatened species54,55; and (3) better use of trade controls to address international demand (e.g., Convention on International Trade in Endangered Species of Wild Fauna and Flora, CITES), particularly with recent increases in the shark meat and ray skin trade, which includes a large suite of reef species56. These recommendations are currently being applied to varying degrees and with varying success. The first two also address climate change threats to coral reefs by increasing resilience by maintaining abundance and conserving biodiversity57,58, and reducing human reliance on reefs through alternative livelihoods59. Without broad-scale action to improve the status of sharks and rays on coral reefs, the declines in global populations reported here will continue, with increasingly dire consequences for the ecosystem health of coral reefs and coastal communities that rely on them.
Methods
IUCN red list assessments
The IUCN Red List of Threatened Species, hereafter referred to as the IUCN Red List, provides the most widely-used approach for assessing extinction risk60. Species evaluated are classified according to the IUCN Categories and Criteria (Version 3.1) following the Guidelines for Using the IUCN Red List Categories and Criteria and these categories range from Least Concern to Extinct, with those that have an elevated risk of extinction considered threatened (Vulnerable, Endangered, or Critically Endangered)61. Extant species assessed as not having an elevated risk of extinction are placed in the categories of Least Concern or Near Threatened (i.e., those that may soon be eligible for a threatened category). The coral reef species were assessed as part of a nine-year effort to reassess all 1199 species, through 17 taxonomic and thematic workshops, with 353 participants from 244 unique experts and/or members of the IUCN Species Survival Commission (SSC) Shark Specialist Group (SSG) from 71 countries and territories (Supplementary Data 3). The collation of information, assessment, peer-review, and consultation process are detailed in Dulvy et al. (2021). Some of the coral reef shark assessments required estimation of population reduction using hierarchical Bayesian state-space methods and space-for-time substitution methods and these are detailed in the supplementary files associated with the Red List Assessments28,62.
Species list and attributes
We compiled a list of 134 coral reef associated chondrichthyans (70 rays, 64 sharks). We included all species listed on the IUCN Red List that included ‘coral reef’ as a habitat (n = 126) as of April 17, 2021. We removed 12 of those species for various reasons (Supplementary Table 4), leaving 114. We then added an additional 20 species that also occur at coral reefs at some point during their lives, based on expert knowledge (Supplementary Table 5). Species were split based on their residency pattern on coral reefs, ranging from ‘resident,’ ‘partial resident,’ and ‘transient’. Residents are those species that spend their entire lives on coral reefs (e.g., bluespotted lagoon ray, Taeniura lymma). Partial residents include species that use coral reefs for most of their lives, but use other habitats as well (e.g., bowmouth guitarfish, Rhina ancylostoma). Finally, transient species are those that use reefs occasionally but spend most of their lives in other habitats (e.g., spotted eagle ray, Aetobatus narinari) (Supplementary Data 2). We additionally categorized the species based on their trophic level into filter feeders, small invertivores, large invertivores, mesopredators, and apex predators (Supplementary Data 2). Large invertivores were classified as rays with a maximum size greater than 100 cm disc width. For statistical analyses, these are coded in an ordinal manner from 1 to 3 for residency type (1 for residents to 3 for transients) and from 1 to 5 for trophic level (1 for filter feeders up to 5 for apex predators). Maximum size values were obtained from the published species’ Red List Assessments.
Threats to sharks and rays were analysed at three levels: 5 Biological Resource Use, 5.4 Fishing and harvesting aquatic resources), and 5.4.1 Intentional mortality (human use - subsistence/small scale))63. They were determined based on coded threats in the IUCN Red List assessment for each species with the exception of climate change64. Climate change was also included as a threat if it was mentioned in the text of the ‘threats’ section of an assessment regardless of being included in the coded threats. For example, the Arabian carpetshark (Chiloscyllium arabicum) did not have ‘climate change’ included as a coded threat, however, it was discussed in the threat section text and therefore included65.
Generation length
Generation lengths were taken from IUCN Red List assessments when available, where it is calculated as the mid-point between the age-at-maturity and longevity28. Generation lengths were either taken from published age-at-maturity and longevity for the species, directly from the species’ IUCN Red List assessment, or estimated using proxy data from related species that have known age-at-maturity and longevity (Supplementary Data 2).
Extinction risk and other reef taxa
To compare the status of sharks and rays to other coral reef taxa, the IUCN Red List website (www.iucnredlist.org) was searched on February 4, 2022 for all animal Classes that included at least a single species with ‘coral reef’ listed as a habitat as per their respective IUCN Red List assessment. The numbers of species within each Class and each IUCN Red List status were recorded. Additionally, we compare reef elasmobranchs to the family Epinephelidae (Coral Groupers), which were recently reassessed and consist mostly of coral reef species66. We included a total of 4918 species (4747 through the IUCN website, additional 167 by including all Epinephelidae66, and 134 sharks and rays [this study]). To best estimate the true percent threatened for each species group considering DD species, we considered that DD species were equally as threatened as data-sufficient species as per IUCN guidelines (Eq. 1; www.iucnredlist.org/resources/summary-statistics).
Species traits and threat level
Each species’ geographic range was calculated as the total two-dimensional area of the species’ distribution as mapped in each species’ respective IUCN Red List assessment. To determine the number of management schemes a species fell under, the number of nations was included as a variable, calculated by determining the number of different Exclusive Economic Zones (EEZs) a species’ distribution overlapped with.
Statistical models were conducted with R v.4.1.0 using relevant software packages67. The variables that could best predict extinction risk of a species were estimated using ordinal logistic regression models (ordinal package68). All numeric variables (species range, generation length, maximum linear dimension, number of nations, lower depth) and the ordinal values of residency type and trophic level were tested for correlation (Pearson’s r; PerformanceAnalytics package69). Variables with correlation >0.7 were not included in the same models (Supplementary Fig. 2)70. As shark sizes are measured as total length and rays are generally measured as the disc width, models were tested both with and without maximum sizes nested in measurement type (i.e., total length and disc width). Models performed better when measurement type was not considered, therefore, models were run with the variable ‘maximum linear dimension’, which included both measurement types. For residency type, sharks and rays were split into 55 resident species, 54 partial residents, and 25 transient species (Supplementary Data 2). Due to the high degree of pairwise correlation between each of residency and trophic level traits with maximum body size, these could not occur in the same models with maximum body size and measures of geographic range, which were preferentially included in the top model based on AIC (Supplementary Table 3 and Supplementary Fig. 2). In addition to a null model, we ran 35 proposed models with different combinations of: generation length, residency type (both ordinal and categorical), trophic level (both ordinal and categorical), lower depth limit, maximum linear dimension, number of nations, and species range (Supplementary Table 3). All models were tested for variable collinearity using variance inflation factors (VIF), and only models with all variables returning values of <2 were further considered71. The number of nations a species occurs in was more important at predicting risk than the geographic range and was included in the final model. The most parsimonious model with an AIC value within 2 units of the lowest AIC was selected as the best performing model72. Coefficients were estimated and standardised for effect magnitude of variables in the top ordinal model (coefplot package73).
National trait boosted regression tree analysis
Management typically occurs at the national level, therefore, we used a machine learning algorithm to model the percent of threatened species per nation against 11 national-level covariates as per Yan et al30. (Supplementary Data 4). We used gross domestic product (GDP (mean from 1960–2019) in USD; https://data.worldbank.org/indicator/NY.GDP.MKTP.CD), world governance index (WGI (mean from 1996–2018); https://databank.worldbank.org/source/worldwide- governance-indicators), and human development index (HDI (2019); http://hdr.undp.org/en/content/human-development-index-hdi) as indicators of governance effectiveness. To model both direct and indirect fishing pressure, we used coastal human population size, measured as the number of people living within 100 km of the coast within the distribution of coral reefs in 2020 (https://sedac.ciesin.columbia.edu/data/set/nagdc-population-landscape-climate-estimates-v3), and marine protein consumption (mean from 1961–2013 in g capita−1 day−1;74) to represent the reliance on marine fish products for dietary protein and economic stability. We also included fishing effort, total catch (units), sightings-per-unit-effort (SPUE), and catch-per-unit-effort (CPUE) of all coral reef associated species (all as mean values from 1950–2010)7. We included the 2020 annual mean sea surface temperature75 and the 2020 annual mean primary productivity (measured as chlorophyll-a concentration76) as ecological indicators and included continental shelf area clipped to a bathymetry of 50 m within each nation’s EEZ to the extent that coral reefs are found to denote total habitat availability77.
We used a boosted regression tree (BRT) framework from the XGBoost package78 to model the effect of governance, fishing, and ecological indicators on the percent of threatened species per nation. We separated sharks and rays for analyses due to their differing ecologies and responses to threats. BRTs are a form of machine learning whereby a boosting algorithm is used to fit many decision trees while minimising a loss function. Consequently, BRTs can handle non-linear data and complex interactions and are not limited by collinearity nor missing data79. The correlation (Pearson’s r) among explanatory variables were all below 0.8, and we log-transformed coastal population, GDP, protein consumption, shelf area, primary productivity, fishing effort, catch, SPUE, and CPUE to improve their distributions79. We first used a tuning step to select hyperparameters by varying the learning rate (eta), maximum loss reduction (gamma), the maximum tree depth (max_depth), and the subsample ratio of the training instance (subsample) to minimise the root mean squared error. To suit the 0-1-inflated-beta distribution of our data, our final model was fit with a logistic loss function with the following hyperparameters (sharks, rays): eta = 0.5, 0.2; gamma = 0.5, 0.7; max_depth = 15, 15; subsample = 0.9, 0.3. With this tuning step, we were able to reduce the root mean squared error from 0.093 to 0.078 for sharks and from 0.069 to 0.063 for rays.
We used a form of cross-validation to fit our final model, where we split our data into an 80–20% training-test set split. Due to the stochastic model building process of BRTs, we randomised our data before each iteration and bootstrapped the models for 1000 iterations. We measured the biases of each model by subtracting the predicted percent threatened from the measured value in the test set, whereby a value close to zero would denote minimal prediction bias. We also extracted the relative importance and calculated the marginal effect of each variable for each iteration. All analyses were conducted using the XGBoost package v.1.4.1.178 in R v.4.1.067.
The average bias of the models across 1000 bootstrapped iterations was −2.5 × 10−3 (95% confidence interval: −4.0 × 10−3 to −1.1 × 10−3) for sharks and −1.0 × 10−3 (−2.0 × 10−3 to 1.0 × 10−4) for rays, indicating the models has a negligible bias. The root mean squared error across all iterations was 0.089 (0.088 to 0.089) for sharks and 0.089 (0.089 to 0.089) for rays (Supplementary Fig. 3).
Mapping
To determine global patterns of richness and threat, species maps were produced for all species combined, and sharks and rays separately. All maps were prepared using ArcGIS Pro 2.7.080. We spatially joined the polygons representing species ranges to a hexagonal grid of individual units (cells) that retain their shape and area (~23,322 km2) throughout the globe to generate a species count in each cell. We then projected maps to the Patterson projection. Maps were truncated to 60° latitude because no data existed outside these areas with the exception of Iceland and Alaska, where there has been at least one reported observation of a tiger shark (Galeocerdo cuvier). The tiger shark is reported as a seasonal vagrant and not a resident of these nations, therefore, their removal from the map did not affect the tiger shark’s resident distribution81. For the proportion of threatened species maps, only cells with greater than five species were included to avoid disproportionately high threat in cells with very few species.
Comparison of threat status and global FinPrint BRUVS abundance estimates
BRUVS estimates of MaxN (highest count of species in view at once) were downloaded from the publicly available data in the MacNeil paper27 at (https://doi.org/10.1038/s41586-020-2519-y). Sightings-per-unit-effort was calculated by summing the total MaxN per nation and dividing it by the total hours of footage analysed. Species lists per nation were created using the IUCN Red List assessment geographic ranges. The proportion of species threatened was calculated by dividing the total number of CR, EN, and VU species in a nation over the total number of coral reef sharks and rays in the nation. A beta regression linear model was run to compare the proportion of coral reef sharks and rays threatened in each nation and SPUE of coral reef sharks from the MacNeil paper (betareg package82).
Red list index
The Red List Index (RLI) values from subsequent assessments show changes in relative extinction risk of various taxa over time31,53. The RLI was calculated by experts retrospectively assigning an IUCN Red List category to each species for the years 1970, 1980, and 2005 based on current status and known history of threat severity. In particular, we used reconstructed catch data from the Sea Around Us, which includes the dates, locations, and species groups exploited. With this information, along with an understanding of species-specific traits and response to fishing, we could estimate that if fishing pressure increased throughout the 1970s and reached a peak in the early 2000s, the species in question was likely Least Concern in 1970, possibly Near Threatened by 1980, and likely in a threatened category by 2005. We built a picture of the history of fishing pressure using the Sea Around Us data and worked backwards from the current category to determine if the previous category (e.g., from 2020 to 2005, then 2005 to 1980, then 1980 to 1970) was likely to be: (i) the same, (ii) better by one, or (iii) two categories. Thus, the range of possible choices is highly constrained at each timestep. Though some assessments were completed around the year 2005, using our current understanding of threats and trends, these may not have accurately represented the IUCN Red List status at the time. Therefore, the 2005 RLI may differ from published IUCN Red List assessments. Eight species underwent taxonomic revision and thus, the initial IUCN Red List assessment may no longer be relevant. However, for these eight species, the old taxonomic concept covers most of their range and, therefore, these were not considered to have previously been Not Evaluated (Supplementary Table 6). Once IUCN Red List categories were assigned, the RLI for each year (t) was calculated by multiplying the number of species included (s) in each IUCN Red List category by their respective category weights (Wc; 0 for LC, 1 for NT, 2 for VU, 3 for EN, 4 for CR, and 5 for EX). As per Eq. 2, the product was then summed and divided by the maximum possible product (number of species (N) multiplied by the maximum weight (i.e. W=5)), and subtracted from 1 to achieve a final index ranging between 0 (meaning all species are Extinct) and 1 (meaning all species are Least Concern83):
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
IUCN Red List of Threatened Species assessments are publicly available with links to specific assessments in Supplementary Data 1. All species traits used in analyses are available in Supplementary Data 2. All covariate values used for the national trait boosted regression tree analysis and the source of these values are available in Supplementary Data 4. Source data are provided with this paper.
Code availability
Code used to reproduce the analysis can be accessed at https://github.com/sammsherman27/CoralReefSharkRayIUCN and at https://doi.org/10.5281/zenodo.726790484.
References
Plaisance, L., Caley, M. J., Brainard, R. E. & Knowlton, N. The diversity of coral reefs: what are we missing? PLoS One. 6, e25026 (2011).
Hoegh-Guldberg, O., Poloczanska, E. S., Skirving, W. & Dove, S. Coral reef ecosystems under climate change and ocean acidification. Front. Mar. Sci. 4, 158 (2017).
Mora, C. et al. Global human footprint on the linkage between biodiversity and ecosystem functioning in reef fishes. PLoS Biol. 9, e1000606 (2011).
Burke, L., Reytar, K., Spalding, M. & Perry, A. Reefs at Risk Revisited. 130 pp. (World Resources Institute, Washington, D.C., 2011).
Hicks, C. C., Graham, N. A. J., Maire, E. & Robinson, J. P. W. Secure local aquatic food systems in the face of declining coral reefs. One Earth. 4, 1214–1216 (2021).
Cinner, J. E. et al. Gravity of human impacts mediates coral reef conservation gains. PNAS 115, E6116–E6125 (2018).
Eddy, T. D. et al. Global decline in capacity of coral reefs to provide ecosystem services. One Earth. 4, 1278–1285 (2021).
Graham, N. A. J. et al. Human disruption of coral reef trophic structure. Curr. Biol. 27, 231–236 (2017).
Sherman, C. S., Heupel, M. R., Moore, S. K., Chin, A. & Simpfendorfer, C. A. When sharks are away rays will play: effects of top predator removal in coral reef ecosystems. Mar. Ecol. Prog. Ser. 641, 145–157 (2020).
Ruppert, J. L. W., Travers, M. J., Smith, L. L., Fortin, M.-J. & Meekan, M. G. Caught in the middle: combined Impacts of shark removal and coral loss on the fish communities of coral reefs. PLoS One. 8, e74648 (2013).
Last, P. R. et al. Rays of the World. (CSIRO Publishing, 2016).
Ebert, D. A., Dando, M. & Fowler, S. Sharks of the World. 2nd edn, 608 (Princeton University Press, 2021).
Heupel, M. R., Lédée, E. J. I. & Simpfendorer, C. A. Telemetry reveals spatial separation of co-occurring reef sharks. Mar. Ecol. Prog. Ser. 589, 179–192 (2018).
Heupel, M. R., Papastamatiou, Y. P., Espinoza, M., Green, M. E. & Simpfendorfer, C. A. Reef shark science - key questions and future directions. Front. Mar. Sci. 6, 12 (2019).
Roff, G., Brown, C. J., Priest, M. A. & Mumby, P. J. Decline of coastal apex shark populations over the past half century. Commun. Biol. 1, 223 (2018).
Williams, J. J., Papastamatiou, Y. P., Caselle, J. E., Bradley, D. & Jacoby, D. M. P. Mobile marine predators: an understudied source of nutrients to coral reefs in an unfished atoll. Proc. R. Soc. B. 285, 20172456 (2018).
Heithaus, M. R., Wirsing, A. J. & Dill, L. M. The ecological importance of intact top-predator populations: a synthesis of 15 years of research in a seagrass ecosystem. Mar. Freshw. Res. 63, 1039–1050 (2012).
Peel, L. R. et al. Stable isotope analyses reveal unique trophic role of reef manta rays (Mobula alfredi) at a remote coral reef. R. Soc. Open Sci. 6, 190599 (2019).
O’Shea, O. R., Thums, M., van Keulen, M. & Meekan, M. Bioturbation by stingrays at Ningaloo Reef, Western Australia. Mar. Freshw. Res. 63, 189–197 (2012).
Takeuchi, S. & Tamaki, A. Assessment of benthic disturbance associated with stingray foraging for ghost shrimp by aerial survey over an intertidal sandflat. Continental Shelf Res. 84, 139–157 (2014).
Burkholder, D. A., Heithaus, M. R., Fourqurean, J. W., Wirsing, A. & Dill, L. M. Patterns of top-down control in a seagrass ecosystem: could a roving apex predator induce a behaviour-mediated trophic cascade? J. Anim. Ecol. 82, 1192–1202 (2013).
Creel, S. & Christianson, D. Relationships between direct predation and risk effects. TRENDS Ecol. Evolution. 23, 194–201 (2008).
Ward-Paige, C. A. et al. Large-scale absence of sharks on reefs in the greater-Caribbean: a footprint of human presence. PLoS One. 5, e11968 (2010).
Espinoza, M., Cappo, M., Heupel, M. R., Tobin, A. J. & Simpfendorfer, C. A. Quantifying shark distribution patterns and species-habitat associations: implications of marine park zoning. PLoS One. 9, e106885 (2014).
Graham, N. A., Spalding, M. D. & Sheppard, C. R. Reef shark declines in remote atolls highlight the need for multi-faceted conservation action. Aquat. Conserv.: Mar. Freshw. Ecosyst. 20, 543–548 (2010).
Nadon, M. O. et al. Re-creating missing population baselines for Pacific reef sharks. Conserv. Biol. 26, 493–503 (2012).
MacNeil, M. A. et al. Global status and conservation potential of reef sharks. Nature 583, 801–806 (2020).
Dulvy, N. K. et al. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 1–15 (2021).
Walls, R. H. L. & Dulvy, N. K. Eliminating the dark matter of data deficiency by predicting the conservation status of Northeast Atlantic and Mediterranean Sea sharks and rays. Biol. Conserv. 246, 108459 (2020).
Yan, H. F. et al. Overfishing and habitat loss drives range contraction of iconic marine fishes to near extinction. Science Adv. 7, eabb6026, (2021).
Butchart, S. H. M. et al. Using Red List Indices to measure progress towards the 2010 target and beyond. Philos. Trans. R. Soc. B 360, 255–268 (2005).
Sherman, C. S. et al. Taeniura lymma. The IUCN Red List of Threatened Species, eT116850766A116851089 (2021). 10.2305/IUCN.UK.2021-1.RLTS.T116850766A116851089.en
Pacoureau, N. et al. Half a century of global decline in oceanic sharks and rays. Nature 589, 567–571 (2021).
Cardeñosa, D. et al. Small fins, large trade: a snapshot of the species composition of low-value shark fins in the Hong Kong markets. Anim. Conserv. 23, 203–211 (2019).
Haque, A. B. & Spaet, J. L. Y. Trade in threatened elasmobranchs in the Bay of Bengal, Bangladesh. Fish. Res. 243, 106059 (2021).
Alcala, A. C. & Russ, G. R. A direct test of the effects of protective management on abundance and yield of tropical marine resources. ICES J. Mar. Sci. 47, 40–47 (1990).
Serrano, A. et al. Effects of anti-trawling artificial reefs on ecological indicators of inner shelf fish and invertebrate communities in the Cantabrian Sea (southern Bay of Biscay). J. Mar. Biol. Assoc. U. Kingd. 91, 623–633 (2011).
Cortés, E. Perspectives on the intrinsic rate of population growth. Methods Ecol. Evolution. 7, 1136–1145 (2016).
McClenachan, L., Cooper, A. B. & Dulvy, N. K. Rethinking trade-driven extinction risk in marine and terrestrial megafauna. Curr. Biol. 26, 1–7 (2016).
Tamburello, N., Cote, I. M. & Dulvy, N. K. Energy and the scaling of animal space use. Am. Naturalist 186, 196–211 (2015).
Dulvy, N. K. et al. Challenges and priorities in shark and ray conservation. Curr. Biol. 27, R565–R572 (2017).
Davidson, L. N. K. & Dulvy, N. K. Global marine protected areas to prevent extinctions. Ecol. Evolution. 1, 1–6 (2017).
Pauly, D., Zeller, D. & Palomares, M. L. D. Sea Around Us Concepts, Design and Data, <http://seaaroundus.org> (2021).
Simpfendorfer, C. A. & Dulvy, N. K. Bright spots of sustainable shark fishing. Curr. Biol. 27, R83–R102 (2017).
Booth, H., Squires, D. & Milner-Gulland, E. J. The mitigation hierarchy for sharks: a risk-based framework for reconciling trade-offs between shark conservation and fisheries objectives. Fish. Fish. 21, 269–289 (2019).
Grorud-Colvert, K. et al. The MPA Guide: A framework to achieve global goals for the ocean. Science 373, eabf0861 (2021).
Enright, S. R., Meneses-Orellana, R. & Keith, I. The Eastern Tropical Pacific Marine Corridor (CMAR): The emergence of a voluntary regional cooperation mechanism for the conservation and sustainable use of marine biodiversity within a fragmented regional ocean governance landscape. Front. Mar. Sci. 8, 674825 (2021).
Chin, A., Kyne, P. M., Walker, T. I. & McAuley, R. B. An integrated risk assessment for climate change: analysing the vulnerability of sharks and rays on Australia’s Great Barrier Reef. Glob. Change Biol. 16, 1936–1953 (2010).
Dwyer, R. G. et al. Individual and population benefits of marine reserves for reef sharks. Curr. Biol. 30, 480–489 (2020).
Speed, C. W., Cappo, M. & Meekan, M. G. Evidence for rapid recovery of shark populations within a coral reef marine protected area. Biol. Conserv. 220, 308–319 (2018).
Mizrahi, M. I., Diedrich, A., Weeks, R. & Pressey, R. L. A systematic review of the socioeconomic factors that influence how marine protected areas impact on ecosystems and livelihoods. Soc. Nat. Resour. 32, 4–20 (2019).
IPBES. Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. 1148 (Bonn, Germany, 2019).
Butchart, S. H. M. et al. Global biodiversity: indicators of recent declines. Science 328, 1164–1168 (2010).
Hanh, T. T. H. & Boonstra, W. J. What prevents small-scale fishing and aquaculture households from engaging in alternative livelihoods? A case study in the Tam Giang lagoon, Viet Nam. Ocean Coast. Manag. 182, 104943 (2019).
Ahmed, N., Troell, M., Allison, E. H. & Muir, J. F. Prawn postlarvae fishing in coastal Bangladesh: challenges for sustainable livelihoods. Mar. Policy. 34, 218–227 (2010).
Prasetyo, A. P. et al. Shark and ray trade in and out of Indonesia: addressing knowledge gaps on the path to sustainability. Mar. Policy. 133, 104714 (2021).
McClanahan, T., Polunin, N. & Done, T. Ecological states and the resilience of coral reefs. Conserv. Ecol. 6, 18 (2002).
Bellwood, D. R., Hughes, T. P. & Hoey, A. S. Sleeping functional group drives coral-reef recovery. Curr. Biol. 16, 2434–2439 (2006).
Cinner, J. E. et al. Vulnerability of coastal communities to key impacts of climate change on coral reef fisheries. Glob. Environ. Change. 22, 12–20 (2012).
Víe, J.-C., Hilton-Taylor, C. & Stuart, S. N. Wildlife in a Changing World - An analysis of the 2008 IUCN Red List of Threatened Species. 180 (Gland, Switzerland, 2009).
Mace, G. M. et al. Quantification of extinction risk: IUCN’s system for classifying threatened species. Conserv. Biol. 22, 1424–1442 (2008).
Sherley, R. B. et al. Estimating IUCN Red List population reduction: JARA - A decision-support tool applied to pelagic sharks. Conserv. Lett. 13, e12688 (2019).
IUCN Red List. Threats Classification Scheme (Version 3.2), <https://www.iucnredlist.org/resources/threat-classification-scheme> (2021).
Salafsky, N. et al. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conserv. Biol. 22, 897–911 (2008).
Moore, A. Chiloscyllium arabicum. The IUCN Red List of Threatened Species 2017, e.T161426A109902537 (2017). 10.2305/IUCN.UK.2017-2.RLTS.T161426A109902537.en
Sadovy de Mitcheson, Y. J. et al. Valuable but vulnerable: Over-fishing and under-management continue to threaten groupers so what now? Mar. Policy. 116, 103909 (2020).
R: A language and environment for statistical computing (R Foundation for Statistical Computing, Vienna, Austria, 2021).
Regression Models for Ordinal Data v. 2019.12.10 (CRAN, 2019).
Econometric Tools for Performance and Risk Analysis v. 2.0.4 (2020).
Dormann, C. F. et al. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46 (2013).
Akinwande, M. O., Dikko, H. G. & Samson, A. Variance inflation factor: As a condition for the inclusion of suppressor variable(s) in regression analysis. Open J. Stat. 5, 754–767 (2015).
Burnham, K. P. & Anderson, D. R. Multimodel inference: understanding AIC and BIC in model selection. Sociological Methods Res. 33, 261–304 (2004).
Plots Coefficients from Fitted Models v. 1.2.8 (2022).
Fisheries and Aquaculture Software. FishStatJ - Software for Fishery and Aquaculture Statistical Time Series., <http://www.fao.org/fishery/> (2020).
Reynolds, R. W., Rayner, N. A., Smith, T. M., Stokes, D. C. & Wang, W. An improved in situ and satellite SST analysis for climate. J. Clim. 15, 1609–1625 (2002).
NASA Ocean Biology (OB.DAAC). Mean annual sea surface chlorophyll-a concentration for the period 2009-2013 (composite dataset created by UNEP-WCMC). Data obtained from the Moderate Resolution Imaging Spectroradiometer (MODIS) Aqua Ocean Colour website (NASA OB.DAAC, Greenbelt, MD, USA), <http://oceancolor.gsfc.nasa.gov/cgi/l3> (2014).
General Bathymetric Chart of the Oceans. GEBCO_2014 Grid. version 20150318. www.gebco.net (2015).
XGBoost: A Scalable Tree Boosting System v. 1.4.1.1 (In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining (pp. 785–794). New York, NY, USA: ACM, 2016).
Elith, J., Leathwick, J. R. & Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 77, 802–813 (2008).
ArcGIS Pro 2.7.0 (Environmental Systems Research Institute) (2020).
Ferreira, L. C. & Simpfendorer, C. Galeocerdo cuvier. The IUCN Red List of Threatened Species 2019, e.T39378A2913541 (2019).
Beta Regression v. 3.1-4 (2021).
Butchart, S. H. et al. Improvements to the Red List Index. PLoS ONE. 2, e140 (2007).
Sherman, C. S. et al. Half a century of rising extinction risk of coral reef sharks and rays, sammsherman27/CoralReefSharkRayIUCN: Data and Code Used in Sherman et al. Half a century of rising extinction risk of coral reef sharks and rays v1.0.0. https://doi.org/10.5281/zenodo.7267904 (2022).
Acknowledgements
We thank all members of the IUCN Species Survival Commission Shark Specialist Group and other experts who contributed to the data collation and, in particular, R. Barreto, A. Cevallos, C. Dudgeon, D. Ebert, A. González, P.M. Kyne, A. Maung, J.M. Morales-Saldaña, L. Seyha, A. Sianipar, D. Tanay, and V.Q. Vo (Supplementary Data 3). We thank W.W.L. Cheung and V.W.Y. Lam for their contributions to the coral reef catch and effort data. We thank C.M. Pollock and C. Hilton-Taylor for quality-controlling data submission through the IUCN Red List. The scientific results and conclusions, as well as any views or opinions expressed herein, are those of the author(s) and do not necessarily reflect those of institutions or data providers. This project was funded by the Shark Conservation Fund, a philanthropic collaborative pooling expertise and resources to meet the threats facing the world’s sharks and rays. The Shark Conservation Fund is a project of Rockefeller Philanthropy Advisors. This work was funded by the Shark Conservation Fund as part of the Global Shark Trends Project to N.K.D. and C.A.S. N.K.D. was supported by Natural Science and Engineering Research Council Discovery and Accelerator Awards and the Canada Research Chairs Program.
Author information
Authors and Affiliations
Contributions
C.S.S., C.A.S., and N.K.D. conceptualized the paper and the analysis. C.S.S., C.A.S., N.P., J.M., H.F.Y., R.W., and N.K.D. conducted the statistical analyses and visualised the data. J.M., J.C., D.D., and K.H. created and compiled species maps. C.A.S. and N.K.D. acquired the funding. C.S.S., C.A.S., and N.K.D. wrote the first draft. C.S.S., C.L.R., W.J.V., R.P., C.A.S., J.C., P.C., R.J., A.B.A., F., C.G.A.C., B.K., M.P.B.P., D., M.E., D.F., A.B.H, P.A.M.F., A.F.N., J.C.P.J., J.U., R.Y., and N.K.D. contributed significantly to Red List assessments that were used in this paper. T.E. and M.L.D.P. provided catch and effort data for coral reefs. All authors contributed to writing the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Imanol Miqueleiz and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sherman, C.S., Simpfendorfer, C.A., Pacoureau, N. et al. Half a century of rising extinction risk of coral reef sharks and rays. Nat Commun 14, 15 (2023). https://doi.org/10.1038/s41467-022-35091-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-022-35091-x
This article is cited by
-
Management and research efforts are failing dolphins, porpoises, and other toothed whales
Scientific Reports (2024)
-
Bioturbation by Benthic Stingrays Alters the Biogeomorphology of Tidal Flats
Ecosystems (2024)
-
A wandering wedgefish illustrates the need for cooperative elasmobranch conservation in the Arabian Gulf
Environmental Biology of Fishes (2024)
-
Extinction risk of the world’s chondrichthyan fishes: a global assessment of the interplay between anthropogenic factors and marine protected areas
Reviews in Fish Biology and Fisheries (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
