
Abstract
Plant–mycorrhizal interactions mediate plant nitrogen (N) limitation and can inform model projections of the duration and strength of the effect of increasing CO2 on plant growth. We present dendrochronological evidence of a positive, but context-dependent fertilization response of Quercus rubra L. to increasing ambient CO2 (iCO2) along a natural soil nutrient gradient in a mature temperate forest. We investigated this heterogeneous response by linking metagenomic measurements of ectomycorrhizal (ECM) fungal N-foraging traits and dendrochronological models of plant uptake of inorganic N and N bound in soil organic matter (N-SOM). N-SOM putatively enhanced tree growth under conditions of low inorganic N availability, soil conditions where ECM fungal communities possessed greater genomic potential to decay SOM and obtain N-SOM. These trees were fertilized by 38 years of iCO2. In contrast, trees occupying inorganic N rich soils hosted ECM fungal communities with reduced SOM decay capacity and exhibited neutral growth responses to iCO2. This study elucidates how the distribution of N-foraging traits among ECM fungal communities govern tree access to N-SOM and subsequent growth responses to iCO2.
Similar content being viewed by others

Introduction
Gross primary productivity (GPP) has been globally stimulated by rising anthropogenic [CO2]1,2 and Earth system models (ESM) suggest this effect could continue to ca. 20703. While global-scale studies infer a moderate historical fertilization effect1,4, evidence for rising CO2 stimulating productivity at the ecosystem scale in mature forests has proven elusive5,6,7. This incongruity has limited accurate constraints of the fertilization effect in ESM3, which is critical to predicting terrestrial carbon feedbacks that may continue to mitigate anthropogenic emissions8,9. Manipulative CO2 enrichment experiments in mature forests record positive, albeit modest, and saturating growth responses to elevated CO2 (eCO2)5,7,10,11,12. Tree-ring studies investigating long-term plant biomass response to increasing ambient CO2 (iCO2) generally observe a weak fertilization effect6,12,13,14,15,16. These observations stand in contrast to the significant responses observed in early successional ecosystems10,12,17, suggesting that any stimulatory effect of CO2 may be transient. Limited nitrogen (N) availability, particularly in mature forests is widely implicated to constrain experimental and ambient growth responses to CO210,12,18.
Plant N limitation is typically linked to the availability of inorganic N, which is made available via the microbial mineralization of soil organic matter (SOM)19. In contrast, N organically bound in soil organic matter (N-SOM), by far the largest ecosystem pool of soil N20, is generally considered inaccessible to plants and is very rarely modeled to contribute to plant N budgets in ESM21. However, there is renewed interest in the possibility that acquisition of N-SOM may allow certain plants to “short-circuit” limiting supply rates of inorganic N22,23. In fact, estimates predict that acquisition of N-SOM, in addition to inorganic N sources, is necessary to mediate a sustained and positive plant growth response to eCO224,25,26,27. Owing to the fact that existing supply rates of inorganic N may be insufficient for a sustained fertilization response28, projections of GPP under iCO2 may be improved by considering the joint contribution of N-SOM and inorganic N to plant growth.
Plant acquisition of N-SOM is contingent on the activity of ectomycorrhizal (ECM) fungal symbionts23,29. ECM fungi may acquire N-SOM using enzymatic and non-enzymatic decay mechanisms retained from their free-living saprotrophic ancestors23,30. Despite their well-established role in providing plants with the majority of their annual N29, ECM communities and their N-foraging traits have rarely been studied in relation to plant response to iCO224,26,31,32. Instead, ECM fungi are often implicitly treated as functionally equivalent in their capacity to decay SOM, leading to untested predictions that all ECM-associated host plants will be fertilized by iCO224.
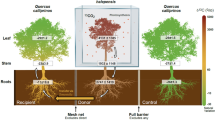
Biological market perspectives emphasizing the metabolic cost of fungal resource capture33 suggest that plants may associate with ECM mutualists that maximize N acquisition and minimize plant carbon (C) expenditure (N return on C investment)34,35. ECM taxa vary widely in their capacity to decay SOM30,36, with greater decay capacity likely carrying a greater C cost to their plant host37. Accordingly, ECM acquisition of N-SOM may be favored under conditions in which inorganic N availability is low38. For example, Cortinarius is a widespread ECM genus with substantial decay capacity39 and is often associated with inorganic N poor soils40. In contrast, ECM taxa that consistently occur in high inorganic N availability soils may specialize in inorganic N acquisition41,42. Although the distribution of ECM taxa are also subject to complex community assembly processes43, we reason that trees associating with ECM communities with greater decay potential (i.e. occurring in inorganic N poor soils) exhibit the largest relativized fertilization response to iCO2 because both N-SOM and inorganic N contribute to tree growth (Fig. 1). In contrast, trees that primarily obtain inorganic N (i.e. occurring in inorganic N rich soils), exhibit a lower relativized growth response to iCO2 (Fig. 1).

These responses occur along a gradient of varying supply rates of inorganic N (net N mineralization rates: x-axis). Dark blue arrows show the relativized fertilization response (arrow width) to iCO2. ECM fungal community composition, morpho-traits, and community aggregated decay traits (CADT) estimated using metagenomic approaches, are hypothesized to vary with soil inorganic N availability. Note hypothesized turnover in the dominance of ECM taxa with extra-radical rhizomorphic hyphae and long-and medium-distance exploration morphologies. White speckles on roots depict hypothesized relative abundance of ECM root-tips. Illustration by Callie R. Chappell, with permission from Reinhard Agerer.
Tree rings represent the outcome of plant growth and are proxies for plant productivity44. Although not without methodological limitations45, dendrochronological studies can be used to study historical responses to iCO26,14,15,16,46. This study had three major goals, (i) elucidate the potential contribution of N-SOM to tree growth, (ii) quantify ECM fungal community aggregated decay traits (CADT)47 along a broad natural soil inorganic N gradient, and (iii) determine if CADT and tree N-SOM acquisition are linked with historical growth responses to iCO2. To accomplish these coupled goals, we studied even-aged (~100 year old) Quercus rubra L. (northern red oak) individuals and their associated ECM communities occurring along a landscape-scale forest mosaic (~50 km) that forms a continuous soil inorganic N gradient (Supplementary Fig. 1)48. Bulk soil properties across this gradient are very similar, however, natural variation in soil inorganic N availability is derived from micro-site differences in nutrient and water retention that have developed following glacial retreat ~10,000 years ago48,49.
In this study we document that Q. rubra exhibits context-dependent growth responses to iCO2. Overall, the largest relative responses occurred under conditions of low inorganic N availability, conditions where N-SOM is likely to contribute to tree growth; acquisition of N-SOM in these conditions is driven by the activity of specialized ECM fungal communities that are enriched in gene families associated with SOM decay. These results highlight the importance of plant-fungal interactions in mediating plant growth response to iCO2 and suggests that ECM activity directly mediates plant access to large and poorly incorporated pools of N-SOM.
Results
Dendrochronological models and plant nitrogen uptake
In order to evaluate the potential contribution of N-SOM to plant growth, we used a change point analysis50 within a dendrochronological Bayesian modeling framework. We asserted that if tree growth, measured as basal area increment (BAI) (cm2/y), is primarily constrained by inorganic N availability51, we can predict a relatively consistent increase in annual growth along a soil gradient of net N mineralization (i.e., inorganic N supply) after accounting for other growth limiting factors (Fig. 2A). However, if tree growth (BAI) is supplemented with N-SOM, the relationship between observed growth rates and the supply of inorganic N should be weakened (smaller slope in Fig. 2A). BAI of focal trees from 1980 to 2017 was analyzed as a function of net N mineralization rates, tree size (to detrend size-age effects), growth in the prior year (to account for lag effects), and as a function of regional yearly climatic conditions (to reflect year to year environmental variability). Finally, BAI was modeled as a function of spatially explicit random effects to account for distances between all sampled focal trees and sites (see Methods). Average minimum May temperature displayed the highest correlation with BAI, whereas annual variability in seasonal and monthly precipitation were only weakly correlated with yearly estimates of BAI and were not included in the final model (Methods). Overall model fit was high (R2 = 0.94; Fig. S2), and we detected a change point in modeled plant growth (BAI) along the N mineralization gradient at 0.53 ± 0.01 μg inorganic N g−1 d−1 (mean ± SD; Fig. 2B).

A Representative analysis of tree growth as a function of net N mineralization rates. The change point analysis identifies the occurrence and location of an inflection point, if any, and the value of the slope parameters on each side. B Basal Area Increment (BAI), from 54 Q. rubra trees along the studied net N mineralization gradient (black circles correspond to an individual growth year). Red and blue lines indicate model estimated BAI mean and 95% PI above and below the identified change-point (BAI change point; estimates were calculated at average values of the other covariates). R2 denotes overall Bayesian model fit. C Slope parameters are significantly different from each other (95% CIs do not overlap; different letters (n = 54). Asterisks indicates parameter is different from zero (95% CI does not overlap with zero).
Consistent with our predictions, there was as significant change-point in the relationship between BAI and rates of net N mineralization, the slope of which was significantly weaker (smaller) to the left of the change point than it was to the right (Fig. 2C; 95% PIs did not overlap). Notably, tree growth below the BAI statistical change-point was significantly greater than predicted based on the contribution of inorganic N alone. After accounting for other growth-limiting variables and in accordance with data presented below, we interpret this higher than predicted growth response as the putative contribution of N-SOM to tree growth.
Ectomycorrhizal community analyses
Concomitant with modeled BAI responses to soil inorganic N availability, there was significant turnover in the composition and morphological attributes of ECM communities. These shifts were generally consistent with the hypothesized context-dependent contribution of N-SOM and inorganic N to tree growth (Fig. 3; Supplementary Fig. 3). The ECM genus Cortinarius dominated in conditions of low inorganic N availability (Fig. 3A)52. Cortinarius, has been widely hypothesized to participate in the acquisition of N-SOM using a potent repertoire of oxidative enzymes that rivals certain free-living saprotrophic fungi42,53. In contrast, the ECM genus Russula which is associated with inorganic N or labile organic N acquisition54, occurred in high relative abundance in soils with high inorganic N availability (Fig. 3B). Other ECM taxa, such as the white-rot derived Hebeloma was found at relatively higher abundance in low inorganic N soils and may also contribute to the functional shifts observed here52,55 (Supplementary Fig. 4). We also examined the relative abundance of ECM taxa forming rhizomorphic hyphae and extraradical emanating hyphae, morpho-traits associated with acquisition of organic N37,56,57. The relative sequence abundance of ECM genera forming rhizomorphic hyphae (F1,110 = 11.32, P = 0.001) and medium-distance exploration types (F1,110 = 14.65, P = 0.0002), was significantly greater for Q. rubra individuals occurring below the statistical BAI change point than in soils above it (Fig. 3C, D).

A, and B relative sequence abundance of the ECM fungal genera Cortinarius and Russula along the gradient of net N mineralization rates (x-axis). Colored bands depict GAM fits. C Box-and whisker plot depicting ECM fungi forming short or medium-distance exploration types. Letters denote statistical significance D. ECM fungi forming rhizomorphic hyphae above and below the BAI statistical threshold (0.53 µgN g soil−1 day−1); letters adjacent to median line of box, denote statistical significance, points are individual communities totaling n = 58. Upper and lower hinges depict 25th and 75th percentiles. E Sum of CAZy gene counts (n = 100 gene families). F Specific gene families (headers) significantly enriched below the BAI change point. AA10, AA11 encode lytic polysaccharide monooxygenases, AA12 is an oxidoreductase, AA3_1: cellobiose dehydrogenase, CE5: acetyl xylan esterase.
In addition to taxonomic and morphological assessments, we employed shotgun metagenomic sequencing of ECM communities inhabiting each of the focal Q. rubra individuals to estimate ECM community aggregated decay traits (CADT)47 and the potential contribution of N-SOM to tree growth. We removed non-fungal sequences and annotated sequences encoding enzymes mediating the decay of SOM47, using the Carbohydrate Active Enzyme and Redoxibase databases (100 gene families studied; Table S2)58,59. We accounted for the compositional nature of the metagenomic data using near-single-copy gene standardization (Methods) and scaled CADT by the number of colonized ECM root-tips encountered on each tree (Supplementary Figs. 5 and 6), reasoning that the net quantity of decay genes hosted on Q. rubra roots is necessary to link N-SOM with host uptake (see Methods).
Q. rubra individuals that exhibited growth responses consistent with N-SOM uptake, i.e., most individuals occurring in low inorganic N soils, hosted ECM communities with distinct decay profiles (Supplementary Figs. 7 and 8). Moreover, these ECM communities were significantly enriched in the total abundance of CAZy genes (F1,54 = 8.38, P = 0.0006; Fig. 3E). Several key gene families that were significantly enriched below the statistically derived BAI change point include cellobiose dehydrogenase (AA3_1) and lytic polysaccharide monooxygenases (LPMO’s: AA9, AA10, AA11) which together act to decay SOM60,61 (Fig. 3F; Supplementary Fig. 9). Many other gene families that are speculated to participate in the decay of SOM also exhibited significant shifts (Supplementary Fig. 9; Supplementary Table 3). Generalized dissimilarity model (GDM) analyses revealed that the compositional abundance of ECM decay genes exhibited a sharp community level threshold response to rates of net N mineralization at approximately 0.5 µg inorganic N g soil−1 day −1 (Supplementary Fig. 9). This threshold response is congruent with the independently derived BAI change point (Fig. 2B), and suggests that ECM communities below this threshold have similar genomic decay potential across a range of soils with low inorganic N availability. Below the modeled BAI change point, differential transcription of certain ECM gene families may explain the greater potential contribution of N-SOM to tree growth with progressively decreasing inorganic N availability. In support of this potential explanation, ECM decay genes are known to be under tight transcriptional regulation, with the expression of key oxidative enzymes inversely correlated with soil inorganic N availability42.
Tree growth response to increasing CO2
The same 38 years of ring data were used to estimate the relative response of plant growth (BAI) to iCO2. During the sampled growth period (1980–2017), atmospheric CO2 increased by ~70 μmol mol−1 (www.NOAA.gov), serving as a natural gradient of iCO2. To compare growth rates along the N mineralization gradient, we calculated annual estimates of growth N efficiencies (GNE) for each tree, \(GNE=\frac{BAI}{N\,mineralization}\). These estimates were standardized, and then analyzed as a function of yearly atmospheric [CO2], average minimum May temperature, and spatially explicit random effects (Fig. 4A, D; see Methods). Slope parameters (λ) (i.e., iCO2 effect) for individual trees were then analyzed as a function of net N mineralization rates using a change point method (Fig. 4B, E). Model estimates derived a change point in the relationship between tree growth and the effect of CO2 (λ) along the N mineralization gradient at a value of 0.39 ± 0.01 μg inorganic N g−1 d−1 (Fig. 4E; Supplementary Fig. 2). Trees growing in soils below this statistical change point tended to have increasing GNE with iCO2 concentrations, resulting in a positive CO2 fertilization response. In contrast, trees inhabiting soils with greater inorganic N availability (to the right of the statistical change point) displayed no changes in GNE over the past 38 years (slopes reflecting the effect iCO2 on plant growth were not different from zero; Fig. 4F). The modeled slope parameters (θ), to the left and right of the change point were significantly different from one another (Fig. 4F).

A Representative analysis framework of growth-nitrogen efficiency index (GNE) as a function of increasing concentrations of historic atmospheric CO2 at each point along the net N mineralization gradient (different lines and their relative slopes). B Conceptual diagram of the effects of iCO2 (λ) on plant growth; change point analysis can detect an inflection point along the soil gradient, if any. C Differences in \({{{{{\mathbf{\theta}}}}}}\) derived from the red and blue portion of panel B, indicate distinct slope values. D Dendrochronological data collected from 54 Q. rubra trees from the past 38 years. Individual points represent estimated annual GNE values colored by tree-specific rates of net-N mineralization. E Individual points represent individual trees response to iCO2 (mean model slopes derived from D) over the study period. Red and blue lines denote Bayesian change-point model with plotted 95% PI (dashed lines; estimated at average values of other covariates) (n = 54). F Denotes mean and 95% CI for the red and blue slopes depicted in E; different letters denote significant differences between slopes, asterisks indicate significant differences from zero (95% CI do not include zero; n = 54).
Discussion
Acquisition of N-SOM has been widely implicated as a primary determinant of plant response to rising CO212,24,26,27, but has been challenging to disambiguate in forest ecosystems.
We present evidence suggesting that plant acquisition of N-SOM is contingent on the community N-foraging traits of ECM fungi and secondly that N-SOM contributes to a positive plant growth response to iCO2. In support of our hypotheses, tree growth responses to iCO2 were context dependent and primarily positive for Q. rubra individuals where N-SOM was likely to contribute to tree growth. In contrast, trees occupying soils with relatively high inorganic N availability, modeled as reliant on inorganic N alone, displayed no positive growth response to iCO2. The near convergence of the change-points derived from both dendrochronological analyses (BAI & GNE), supports the role of N-SOM in stimulating a positive CO2 fertilization response. Furthermore, the synchronous shifts in ECM community composition, morpho-traits associated with organic N acquisition and metagenomic estimates of SOM decay capacity (CADT) largely aligns with the independent dendrochronological models, supporting a strong role of ECM fungi in modulating Q. rubra N acquisition and growth responses to iCO2. Additional evidence for a transition from assimilation of N-SOM to predominately inorganic N economies comes from a previous study of these individuals trees, which found δ15N depleted foliage under conditions of low inorganic N availability suggestive of enhanced organic N uptake, and relatively enriched foliage under high inorganic N availability52. Overall, assumptions of plant reliance on inorganic N alone, or alternatively, ubiquitous plant access to N-SOM via ECM symbionts, are each unable to resolve the context dependent growth responses to iCO2 observed here.
Our investigation of ECM communities builds towards a novel perspective on the contribution of N-SOM to plant growth by investigating possible N-foraging trait trade-offs along a soil nutrient gradient38. Shifts in the morphological and genomic traits of ECM communities across the soil mineralization gradient could result from niche based processes and metabolic tradeoffs in the cost of foraging for N-SOM vs. inorganic N (N return on C investment)38. For example, ECM communities inhabiting trees in low inorganic N soils were enriched in C intensive hyphal morphologies (rhizomorphs and medium-distance exploration types) associated with organic N acquisition37. These traits may be especially favored under conditions of low inorganic N availability if they are associated with enhanced photosynthate allocation and persistence on host roots38. Moreover, these ECM communities were enriched in key gene families that together form a lytic polysaccharide monooxygenase (LPMO)61 decay pathway. Notably, previous laboratory studies have shown that ECM fungi express LPMO to decay SOM62. While certain CAZy gene families studied here are very likely implicated in SOM decay, many CAZy have additional intracellular roles or are involved in mycorrhizal initiation, and can therefore be challenging to demonstratively ascribe to SOM decay61,63. Manganese peroxidase is a potent oxidative enzyme that certain ECM fungi employ to degrade SOM42; our results demonstrate that the presence of this gene family decreases across the breadth of the gradient, however, it did not exhibit a significant threshold response to the BAI change-point. Comparative analyses that link the enzymatic acquisition of N-SOM with transcriptomic community-level ECM profiles and plant uptake, would substantially bolster the mechanisms proposed here, but remain technically infeasible under field conditions30. Additionally, metagenomic estimates of absolute functional gene abundance may provide deeper insight into the N-cycling pathways and mechanisms proposed. Future studies studying co-occurring saprotrophic communities may reveal interesting inter-guild interactions that structure plant access to N and SOM dynamics64.
Our results support predictions asserting that supply rates of inorganic N are insufficient to engender a sustained positive plant growth response to rising CO228,65. This may partially explain the results of certain tree-ring studies finding minimal effect of iCO2 on the accumulation of plant biomass6,14,16. Moreover, this interpretation is consistent with evidence from Free Air CO2 Enrichment (FACE) experiments that document negligible effects of eCO2 on plant growth unless additional sources of N are added, typically in the form of inorganic N17,66,67. In the current study, tree association with ECM communities with enhanced capacity to obtain N-SOM may have increased plant N assimilation beyond that made possible by endogenous supply rates of inorganic N alone. In contrast for trees with high existing growth rates, greater C allocation towards ECM symbionts that specialize on inorganic N acquisition may not translate to greater plant N uptake and positive responses to iCO2. This is due to the fact that inorganic N release is mediated by free living soil saprotrophs19, whose activity is largely independent from ECM fungi. In contrast, progressive increases in C allocation to ECM communities with high N-SOM acquisition potential, could provide a direct mechanism for plants to enhance N uptake under iCO2 conditions25,68, but see69. Accordingly, the responses we describe are consistent with previous observations that identify greater N uptake as necessary for positive responses to CO2, and not merely from greater N use efficiency70. We note that the addition of inorganic N in certain previous studies is likely to disfavor ECM decay of SOM54, in these cases precluding accurate assessment of the role of N-SOM in stimulating tree growth under eCO2. Finally, although a recent study suggested that ECM acquisition of N-SOM may reduce soil C stocks under rising CO2 conditions27, our results are not consistent with evidence of asymmetric soil C depletions.
While the evidence we have accumulated provides strong support for the role of ECM fungi and N-SOM in mediating plant response to iCO225, it simultaneously challenges expectations that ECM associated trees will display a positive growth response to rising CO224,26,32. Because the positive growth responses to iCO2 reported here are relativized and primarily occurred for the individuals with the smallest initial BAI, our study suggests that positive biomass responses to iCO2 may be modest, and if they can be extrapolated, suggests that ECM associated tree response to iCO2 is overestimated28,32. In fact our findings overall range from a modest to neutral growth response to iCO2 which are in line with available evidence for eCO2 experiments conducted in mature ecosystems7. Together, our work further highlights the importance of nutrient limitation in iCO2 responses, suggesting that studies derived from aggrading or early successional ecosystems may be poor predictors of forest C sequestration potential under future CO2 regimes12. Further studies on a wider range of ECM and AM communities and host trees, distributed across forest biomes, particularly the tropics, are needed to determine the generality of our findings.
Our field-based study is not without certain caveats. To address some of them, our modeling framework directly accounted for historic increases in temperature, thereby allowing us to isolate the effect of iCO2 on individual-level plant growth. It is also possible that other limiting nutrients such as phosphorus (P)32 could account for the distinct growth patterns found here. This is unlikely, however, as prior study of soils in this region show small differences in P availability71; moreover, P is unlikely to be a key limiting nutrient in these young soils (<10,000 yrs). Similarly, previous studies in this region also suggest that free-living and symbiotic N-fixation are unlikely to provide additional sources of inorganic N that we did not measure72. While historic rates of N deposition are inaccessible at the scale of individual focal trees or plots studied here, they are also unlikely to vary significantly across plots, as our study area is small and uniformly distant from anthropogenic point sources of N pollution. Additionally, the relative differences in inorganic N availability among forest plots studied here have remained stable since at least the mid 1980’s48,49, and the minimal disturbances in this late-successional ecosystem also suggests relative equilibrium in the distinct N-cycling pathways. A key consideration of our proposed mechanism is that N-SOM has differentially contributed to plant growth for the duration of the study period. While we cannot directly test this assumption, previous analyses of ECM communities occupying these soils suggest that communities have remained relatively stable, supporting this possibility73. Further study on the mechanistic role of rhizosphere priming under iCO2 conditions is needed31, but alone does not seem capable of driving the context-dependent results reported here. Finally, we acknowledge that there are considerable biases associated with sample design that affect the interpretation of dendrochronological data such as measuring only a subset of dominant trees in an area which if non-randomly selected can easily bias towards the preferential selection of large diameter age classes45. We address some of these issues by employing relativized analyses of host trees, selected at random, at small geographic scales45.
Plant productivity responses to iCO2 remain one of the largest uncertainties in projections of the terrestrial C sink8,9,74. We provide dendrochronological evidence of a context-dependent stimulation of tree growth by iCO2 in a mature temperate forest ecosystem; this response is putatively driven by the uneven contribution of N-SOM to tree growth. Functional trait trade-offs in the N-foraging attributes of ECM fungal communities may govern plant growth responses to iCO2; this finding provides important nuance to predictions asserting that all ECM associated trees will respond positively to iCO2. Moreover, we highlight that singular emphasis on inorganic N in existing ESM may lead to spurious conclusions regarding the strength and duration of the CO2 fertilization effect28,75. In these respects, our findings add further evidence that projections of substantial increases in terrestrial C storage driven by CO2 fertilization of GPP are likely overestimated76. While incorporation of ECM communities into existing ESM is not feasible, further work will determine if inorganic N availability can represent a reliable proxy for plant access to N-SOM in ecosystems dominated by ECM fungi. In conclusion, the accurate representation of plant growth response to iCO2 and the terrestrial C sink in ESM appears to be contingent on incorporating N-SOM, a previously unrecognized and cryptic source of plant N.
Methods
Site conditions
Our study took place across a regional network of 12 forest sites comprising a natural N soil gradient in Manistee National Forest in northwestern Lower Michigan (Supplementary Fig. 1). These sites have been described in full elsewhere52. Briefly, annual rates of net N mineralization range from 38 to 120 kg N ha−1 y−1 (calculated from48), which broadly spans soil inorganic N availability in the upper Lake States region77,78. The forest stands are even-aged (~100 years old), resulting from regrowth following forest clearing in the early 20th century. Relative differences in soil nutrient availability have persisted for decades due to lack of disturbance48,49. Micro-site climatic differences in nutrient retention have developed in the past ~10,000 yrs resulting in variation in nutrient cycling (Supplementary Fig. 1). An interlobate moraine transects the study region, and a network of outwash plains with slightly coarser textured soils are associated with lower rates of net N mineralization; in contrast, soils occurring in more upland positions on moraines are associated with greater rates of net N mineralization. Soils across the study region are derived from sandy (~85% sand) glacial drift, and range from Typic Udipsamments to Entic Haplorthods. Comparison with previously published rates of net N mineralization for soils from this region suggest that regional variation in inorganic N availability has persisted since at least the mid 1980’s48,49, and likely across their historical development. Moreover, comparison of rates of net N mineralization revealed that the underlying inorganic N gradient studied here is stable across the duration of the growing season (Pearson r = 0.77; Supplementary Fig. 10).
Tree core sampling and measurement
In May 2018, at each of the 12 sites, we randomly selected five mature Q. rubra individuals that were at least 10-m apart, and measured tree diameter at breast height (1.3 m; DBH) in cm. We then extracted growth cores (to the pith) from the North and South aspect of each tree at DBH using 5.15 mm Haglöf increment borers. The samples were dried overnight at 100 °C. We mounted tree cores on cradles and progressively sanded them by hand, from 100 to 600 grit. The mounted cores were digitized on a flatbed scanner at a resolution of 1200 dpi. We measured yearly ring width (growth) of the scanned tree cores using the Cybis CooRecorder program at a precision of 0.001 mm (Cybis Elektronik 2010). We then used the program Cybis CDendro for individual cross dating and chronology assembly by site. Crossdating was achieved when TTest values were greater than 5 for matching target samples using the P2YrsL normalization method79. We created master ring width lists that were summed by stem to reflect the average yearly growth of each individual using cross dated North & South aspects. We estimated historical DBH of focal trees at each year using the yearly ring width from our master chronologies. All overstory plant stems greater than 10 cm DBH and within 10 m radius of each focal Q. rubra individual were measured and identified (Supplementary Figs.: 11-12). Understory plant communities are reported elsewhere52, and no N-fixing taxa in the overstory or understory were encountered.
Characterization of soil properties
Five soil cores, 5-cm diameter and 10-cm deep, were collected both May and August 2018, were taken radially around the dripline of each focal Q. rubra individual following our previous work in these forests52. Soil net N mineralization rates were quantified as an estimate of soil inorganic N availability for soil samples collected in both May and August 2018. Soil inorganic N was extracted from fresh sieved soil using 2 M KCl, followed by a 14-day aerobic incubation in order to measure rates of soil inorganic N mineralization80. NO3− and NH4+ in soil extracts were analyzed colorometrically (AQ2; Seal Analytical, Mequon, WI) (Supplementary Methods). Total free primary amines (TFPA) in soil (primarily amino acids and amino sugars) was measured using fresh sieved soil extracted with 2 M KCl81 (Supplementary Fig. 12). Total C and N and soil pH were processed as described in52.
Analysis of BAI response to N mineralization rates
Annual tree radial growth and diameter at breast height (DBH) at the time of sampling were combined to calculate past DBH by subtracting radial growth each year to the previous year’s DBH. We then calculated annual tree basal area (BA), BA = π(DBH/2)2 and basal area increments (BAI; cm2/y). BAIs were estimated for each tree (i) and year (y) as the difference in BA between two consecutive years:
Fifty four trees yielded high quality ring widths for BAI estimates. We then analyzed these BAI estimates as a function of tree-level N mineralization rates applying a change point analysis that would identify if at any point in the N mineralization gradient the relationship with growth changed (Fig. 2A; Supplementary Table 5). We also accounted for the relationship between tree size (DBH) and growth by including DBH in the analysis of BAI (detrending). Furthermore, because tree growth at any particular year may also be affected by lag effects (growth in previous years)82 we included previous year growth (standardized) as a covariate after exploring how many years back previous growth could have affected current growth, and a lag of one showed the best relationship (Table S5). We carried out exploratory data analysis to identify climatic variables (monthly temperature and precipitation records from the closest NOAA climate station in Cadillac, MI [data was retrieved June 7, 2020, https://www.ncdc.noaa.gov/cdo-web]). Average May minimum temperature was the variable with the highest correlation with BAI (r: 0.11); we included this variable in the analysis (standardized). To also account any spatial autocorrelation, e.g., trees close to each other and site level factors, we included spatially explicit random effects (SERE). For any particular tree i and year y BAI analysis likelihood and process models were:
We followed a Bayesian approach to estimate parameters. Parameter J is an indicator, with value 0 before the change point, and value 1 after. This change point parameter was estimated as: \(Change\,point \sim Uniform(0,1.25),\,\) allowing the change point to fall outside the range of N mineralization sampled (0.06–1.19 μg g−1 d−1). Variability around growth estimates (σ2) was estimated as a function of DBH, since this seems to vary with size83. The rest of the parameters were estimated from non-informative prior distributions. In Eq. (6), d represents the distance between sites i and j.
Characterization of Ectomycorrhizal fungi
In August 2018, ECM root-tips were collected radially around the dripline of each focal Q. rubra individual as previously described52; briefly, five cores were taken around each tree, each core was 10-cm deep and 11 × 11 cm in area. The soil was removed from roots using sequential washing using tap water and ECM root-tips with high turgor were manually excised using a dissecting microscope after visually eliminating non-Quercus roots. In total, 14,944 individual ECM root-tips were excised. DNA was extracted from lyophilized root-tips using the Qiagen DNeasy Plant Mini Kit (Hilden, Germany) and DNA pools were split for amplicon and metagenomic sequencing (see below). The ITS2 fragment of rRNA was amplified using PCR, following Taylor et al.84 (Table S1) and sequenced using Illumina Mi-Seq (2 × 250; San Diego, CA). Sequences were processed using DADA2 v1.16, ASV were assigned taxonomy using the UNITE dynamic database (v.8; 97–99% sequence similarity) with the scikit naive bayes algorithem (v.0.21.0)85,86. We used the DEEMY (characterization and DEtermination of EctoMYcorrhizae) database (http://www.deemy.de/) to gather morphological information on the exploration type (hyphal foraging distance) and rhizomorph occurrence of ECM taxa present in our dataset at >0.5% relative abundance52. We assigned morphological hyphal trait data for 28 ECM genera, comprising more than 93% of all identified ECM sequences.
Metagenomic sequence generation, processing, and annotation
Shotgun metagenomic sequencing was conducted using a NovaSeq 6000 instrument (2 × 150 bp) at the University of Michigan Advanced Genomics Core. In total, 23,203,326,006 metagenomic sequences were generated, and reads were left unmerged. In order to remove non-fungal sequences, we removed sequences that mapped to the UniVec database (bacterial, archaeal, human, viral) sequences, as well as Quercus rubra87 and Qurcus lobata genome assemblies88 using Kraken2 (v. 0.9.29)89. On average, 22% of sequences per sample were removed during this filtering step, yielding a mean of 307,041,274 putative fungal sequences per sample (Fig. S12). Next, we used a direct mapping approach to annotate remaining sequences against the CAZy and Peroxibase reference databases (100 total gene families; Table S2) using ‘sensitive’ DIAMOND (v. 0.9.29)90 and BWA-MEM (v.0.7.17)91 respectively, following best practice for unmerged reads92. The compiled decay gene database primarily contained ‘core’ gene families found to be actively expressed during fungal decay of SOM and microbial biomass93,94 (CAZy: http://www.cazy.org; http://peroxibase.toulouse.inra.fr/), and is hereafter referred to as the ‘CAZy database’ (Table S2). We tabulated the number of near-single copy genes, as a proxy for the number of Dikaryotic fungal genomes present in each sample, using the OrthoDB v.9 gene database, which comprised 1,312 near-single copy gene variants95. Further methodological details are presented in the Supplementary Methods.
Statistical analysis of Ectomycorrhizal fungal composition and metagenomic function
We compared the relative sequence abundance of ECM ASV forming rhizomorphic and medium-distance hyphal morphologies above and below the statistical BAI change point using two-way ANOVA. To account for the compositional nature of the metagenomic decay gene data96 we calculated the logarithm of the number of sequences mapped to a given decay gene family divided by the geometric mean number of orthologous near single-copy features present in the sample (single-copy genes); note that this is identical to an additive log-ratio transformation96. We incorporated potential underlying shifts in the biomass of ECM communities by multiplying single-copy standardized gene counts by the standardized number of colonized ECM root-tips recovered from focal Q. rubra root-systems (Supplementary Fig. 5). To isolate the effect of mineralization rate and other environmental variables in driving shifts in the compositional abundance of the 100 decay gene families, we used generalized dissimilarity models (GDM)97,98. Environmental predictors initially included in the model were net N mineralization rates, pH, soil C and N, C:N, TFPA and gravimetric soil moisture. All abiotic measurements were calculated at the individual tree basis. This model additionally incorporated geographic distances between individual focal trees and Bray-Curtis distance matrices of the abundance of the 100 gene families were used. We used backwards model selection99, and confirmed the significance of remaining predictors using matrix permutation (nperm = 500) (Table S4). To determine the identity of the ECM gene families that were significantly enriched in communities inhabiting trees that exhibited putative uptake of N-SOM, we compared the mean log abundance of individual gene families occurring in ECM communities above and below the statistical BAI changepoint using one-way ANOVA with Bonferroni correction (Table S3). Vegan (v 2.5.6)100 and tidyverse (1.3)101 were also employed for analysis.
Analysis of BAI response to atmospheric CO2
To compare trends in growth across trees we first calculated an index of growth nitrogen efficiency, GNE. This index was estimated for each tree (i) and year (y) as:
To facilitate the analyses, we standardized GNE:
Afterwards, we analyzed GNES as a function of annual atmospheric CO2 (obtained from NOAA 2019), average May minimum temperature (see above), and spatially explicit random effects (Table S6-7). For each tree i and year y:
We then analyzed the effect of CO2, slope parameters λ (mean and variance, \({\bar{\lambda }}_{i}\) and \({\sigma }_{\lambda }^{2}\)), as a function of N mineralization rate; this analysis tested how much of the variability found in this parameter could be attributed to differences in inorganic N availability. We tried several analyses, including exponential decay and logarithmic functions, and change point analysis with two different intercepts. The best model fitting the data, based on Deviance Inference Criterion (DIC)102 was a simple change point analysis (Table S6-7). For each tree i:
Parameter J is an indicator, with value 0 before the change point, and value 1 after. This change point parameter was estimated as: \(Change\,point \sim Uniform(0,1.25).\) Remaining parameters were estimated from non-informative prior distributions.
See supplement for additional information and all analysis code.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Raw DNA sequences associated with the ITS2 amplicon sequencing are deposited in NCBI Sequence Read Archive: SRR14164239-SRR14164298. Metagenomic sequences are deposited under accession codes: SRR15377920-SRR15377978. Associated soil metadata are available in Dryad (https://doi.org/10.5061/dryad.4f4qrfjbt). Access to wood cores will be available upon written request. Publicly available datasets used in this study include CAZy and Redoxibase http://www.cazy.org; http://peroxibase.toulouse.inra.fr/). Univec database: https://ftp.ncbi.nlm.nih.gov/pub/UniVec/. UNITE database: https://unite.ut.ee/. NOAA climatic data https://www.ncdc.noaa.gov/cdo-web]) was used, and in addition, OrthoDB database was used (https://www.orthodb.org/). Finally, the publicly available DEEMY database was accessed at http://www.deemy.de.
Code availability
Custom code for dendrochronological analyses is reported in the Supplementary Methods.
References
Campbell, J. E. et al. Large historical growth in global terrestrial gross primary production. Nature 544, 84–87 (2017).
Schwalm, C. R. et al. Modeling suggests fossil fuel emissions have been driving increased land carbon uptake since the turn of the 20th Century. Sci. Rep. 10, 9059 (2020).
Wenzel, S., Cox, P. M., Eyring, V. & Friedlingstein, P. Projected land photosynthesis constrained by changes in the seasonal cycle of atmospheric CO2. Nature 538, 499–501 (2016).
Piao, S. et al. Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 1, 14–27 (2020).
Ellsworth, D. S. et al. Elevated CO2 does not increase eucalypt forest productivity on a low-phosphorus soil. Nat. Clim. Change 7, 279–282 (2017).
Hararuk, O., Campbell, E. M., Antos, J. A. & Parish, R. Tree rings provide no evidence of a CO2 fertilization effect in old-growth subalpine forests of western Canada. Glob. Change Biol. 25, 1222–1234 (2019).
Jiang, M. et al. The fate of carbon in a mature forest under carbon dioxide enrichment. Nature 580, 227–231 (2020).
Friedlingstein, P. et al. Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks. J. Clim. 27, 511–526 (2014).
Koven, C. D. et al. Controls on terrestrial carbon feedbacks by productivity versus turnover in the CMIP5 earth system models. Biogeosciences 12, 5211–5228 (2015).
Norby, R. J. & Zak, D. R. Ecological lessons from free-air CO2 enrichment (FACE) experiments. Annu. Rev. Ecol. Evol. Syst. 42, 181–203 (2011).
Sigurdsson, B. D., Medhurst, J. L., Wallin, G., Eggertsson, O. & Linder, S. Growth of mature boreal Norway spruce was not affected by elevated [CO2] and/or air temperature unless nutrient availability was improved. Tree Physiol. 33, 1192–1205 (2013).
Walker, A. P. et al. Integrating the evidence for a terrestrial carbon sink caused by increasing atmospheric CO2. N. Phytol. 229, 2413–2445 (2021).
Gedalof, Z. & Berg, A. A. Tree ring evidence for limited direct CO2 fertilization of forests over the 20th century. Glob. Biogeochem. Cycles 24, (2010).
van der Sleen, P. et al. No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nat. Geosci. 8, 24–28 (2015).
Girardin, M. P. et al. No growth stimulation of Canada’s boreal forest under half-century of combined warming and CO2 fertilization. Proc. Natl Acad. Sci. USA 113, E8406–E8414 (2016).
Giguère-Croteau, C. et al. North America’s oldest boreal trees are more efficient water users due to increased [CO2], but do not grow faster. Proc. Natl Acad. Sci. USA 116, 2749–2754 (2019).
Walker, A. P. et al. Decadal biomass increment in early secondary succession woody ecosystems is increased by CO2 enrichment. Nat. Commun. 10, 454 (2019).
Du, E. et al. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. https://doi.org/10.1038/s41561-019-0530-4 (2020).
Schimel, J. P. & Bennett, J. Nitrogen mineralization: challenges of a changing paradigm. Ecology 85, 591–602 (2004).
Vitousek, P. M. & Howarth, R. W. Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry 13, 87–115 (1991).
Taylor, K. E., Stouffer, R. J. & Meehl, G. A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 93, 485–498 (2012).
Näsholm, T., Kielland, K. & Ganeteg, U. Uptake of organic nitrogen by plants. N. Phytol. 182, 31–48 (2009).
Lindahl, B. D. & Tunlid, A. Ectomycorrhizal fungi – potential organic matter decomposers, yet not saprotrophs. N. Phytol. 205, 1443–1447 (2015).
Terrer, C., Vicca, S., Hungate, B. A., Phillips, R. P. & Prentice, I. C. Mycorrhizal association as a primary control of the CO2 fertilization effect. Science 353, 72–74 (2016).
Terrer, C. et al. Ecosystem responses to elevated CO2 governed by plant–soil interactions and the cost of nitrogen acquisition. N. Phytol. 217, 507–522 (2018).
Sulman, B. N. et al. Diverse Mycorrhizal associations enhance terrestrial C storage in a global model. Glob. Biogeochem. Cycles 33, 501–523 (2019).
Terrer, C. et al. A trade-off between plant and soil carbon storage under elevated CO2. Nature 591, 599–603 (2021).
Wieder, W. R., Cleveland, C. C., Smith, W. K. & Todd-Brown, K. Future productivity and carbon storage limited by terrestrial nutrient availability. Nat. Geosci. 8, 441–444 (2015).
Smith, S. E. & Read, D. J. Mycorrhizal symbiosis. (Academic Press, 2010).
Pellitier, P. T. & Zak, D. R. Ectomycorrhizal fungi and the enzymatic liberation of nitrogen from soil organic matter: why evolutionary history matters. N. Phytol. 217, 68–73 (2018).
Phillips, R. P. et al. Roots and fungi accelerate carbon and nitrogen cycling in forests exposed to elevated CO2. Ecol. Lett. 15, 1042–1049 (2012).
Terrer, C. et al. Nitrogen and phosphorus constrain the CO2 fertilization of global plant biomass. Nat. Clim. Change 9, 684–689 (2019).
Christian, N. & Bever, J. D. Carbon allocation and competition maintain variation in plant root mutualisms. Ecol. Evol. 8, 5792–5800 (2018).
Hortal, S. et al. Role of plant–fungal nutrient trading and host control in determining the competitive success of ectomycorrhizal fungi. ISME J. 11, 2666–2676 (2017).
Bogar, L. et al. Plant-mediated partner discrimination in ectomycorrhizal mutualisms. Mycorrhiza 29, 97–111 (2019).
Bödeker, I. T. M., Nygren, C. M. R., Taylor, A. F. S., Olson, Å. & Lindahl, B. D. ClassII peroxidase-encoding genes are present in a phylogenetically wide range of ectomycorrhizal fungi. ISME J. 3, 1387–1395 (2009).
Hobbie, E. A. & Agerer, R. Nitrogen isotopes in ectomycorrhizal sporocarps correspond to belowground exploration types. Plant Soil 327, 71–83 (2010).
Koide, R. T., Fernandez, C. & Malcolm, G. Determining place and process: functional traits of ectomycorrhizal fungi that affect both community structure and ecosystem function. N. Phytol. 201, 433–439 (2014).
Lindahl, B. D. et al. A group of ectomycorrhizal fungi restricts organic matter accumulation in boreal forest. Ecol. Lett. 24, 1341–1351 (2021).
van der Linde, S. et al. Environment and host as large-scale controls of ectomycorrhizal fungi. Nature 558, 243–248 (2018).
Read, D. J. & Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems: a journey towards relevance? N. Phytol. 157, 475–492 (2003).
Bödeker, I. T. M. et al. Ectomycorrhizal Cortinarius species participate in enzymatic oxidation of humus in northern forest ecosystems. N. Phytol. 203, 245–256 (2014).
Bogar, L. & Peay, K. Processes maintaining the coexistence of ectomycorrhizal fungi at a fine spatial scale. in Biogeography of Mycorrhizal Symbiosis (ed. Tedersoo, L.) vol. 230 79–105 (Springer, 2017).
Xu, K. et al. Tree-ring widths are good proxies of annual variation in forest productivity in temperate forests. Sci. Rep. 7, 1945 (2017).
Nehrbass‐Ahles, C. et al. The influence of sampling design on tree-ring-based quantification of forest growth. Glob. Change Biol. 20, 2867–2885 (2014).
Mathias, J. M. & Thomas, R. B. Disentangling the effects of acidic air pollution, atmospheric CO2, and climate change on recent growth of red spruce trees in the Central Appalachian Mountains. Glob. Change Biol. 24, 3938–3953 (2018).
Fierer, N., Barberán, A. & Laughlin, D. C. Seeing the forest for the genes: using metagenomics to infer the aggregated traits of microbial communities. Front. Microbiol. 5, 614 (2014).
Zak, D. R. & Pregitzer, K. S. Spatial and temporal variability of nitrogen cycling in northern lower Michigan. Science 36, 367–380 (1990).
Zak, D. R., Pregitzer, K. S. & Host, G. E. Landscape variation in nitrogen mineralization and nitrification. Can. J. Res. 16, 1258–1263 (1986).
Chen, J. & Gupta, A. K. Parametric Statistical Change Point Analysis: With Applications to Genetics, Medicine, and Finance. (Springer Science & Business Media, 2011).
Thomas, R. Q., Canham, C. D., Weathers, K. C. & Goodale, C. L. Increased tree carbon storage in response to nitrogen deposition in the US. Nat. Geosci. 3, 13–17 (2010).
Pellitier, P. T., Zak, D. R., Argiroff, W. A. & Upchurch, R. A. Coupled shifts in ectomycorrhizal communities and plant uptake of organic nitrogen along a soil gradient: an isotopic perspective. Ecosystems (2021).
Sterkenburg, E., Clemmensen, K. E., Ekblad, A., Finlay, R. D. & Lindahl, B. D. Contrasting effects of ectomycorrhizal fungi on early and late stage decomposition in a boreal forest. ISME J. 12, 2187–2197 (2018).
Lilleskov, E. A., Hobbie, E. A. & Fahey, T. J. Ectomycorrhizal fungal taxa differing in response to nitrogen deposition also differ in pure culture organic nitrogen use and natural abundance of nitrogen isotopes. N. Phytol. 154, 219–231 (2002).
Kohler, A. et al. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 47, 410–415 (2015).
Moeller, H. V., Peay, K. G. & Fukami, T. Ectomycorrhizal fungal traits reflect environmental conditions along a coastal California edaphic gradient. FEMS Microbiol. Ecol. 87, 797–806 (2014).
Defrenne, C. E. et al. Shifts in Ectomycorrhizal fungal communities and exploration types relate to the environment and fine-root traits across interior douglas-fir forests of Western Canada. Front. Plant Sci. 10, 643 (2019).
Fawal, N. et al. PeroxiBase: a database for large-scale evolutionary analysis of peroxidases. Nucleic Acids Res. 41, D441–D444 (2013).
Lombard, V., Golaconda Ramulu, H., Drula, E., Coutinho, P. M. & Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 42, D490–D495 (2014).
Garajova, S. et al. Single-domain flavoenzymes trigger lytic polysaccharide monooxygenases for oxidative degradation of cellulose. Sci. Rep. 6, 28276 (2016).
Janusz, G. et al. Lignin degradation: microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 41, 941–962 (2017).
Shah, F. et al. Ectomycorrhizal fungi decompose soil organic matter using oxidative mechanisms adapted from saprotrophic ancestors. N. Phytol. 209, 1705–1719 (2016).
Baldrian, P. Fungal laccases – occurrence and properties. FEMS Microbiol. Rev. 30, 215–242 (2006).
Fernandez, C. W. & Kennedy, P. G. Revisiting the ‘Gadgil effect’: do interguild fungal interactions control carbon cycling in forest soils? N. Phytol. 209, 1382–1394 (2016).
Reich, P. B. et al. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 440, 922–925 (2006).
Oren, R. et al. Soil fertility limits carbon sequestration by forest ecosystems in a CO2-enriched atmosphere. Nature 411, 469–472 (2001).
Norby, R. J., Warren, J. M., Iversen, C. M., Medlyn, B. E. & McMurtrie, R. E. CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc. Natl Acad. Sci. USA 107, 19368–19373 (2010).
Andrew, C. & Lilleskov, E. A. Productivity and community structure of ectomycorrhizal fungal sporocarps under increased atmospheric CO2 and O3. Ecol. Lett. 12, 813–822 (2009).
Näsholm, T. et al. Are ectomycorrhizal fungi alleviating or aggravating nitrogen limitation of tree growth in boreal forests? N. Phytol. 198, 214–221 (2013).
Finzi, A. C. et al. Increases in nitrogen uptake rather than nitrogen-use efficiency support higher rates of temperate forest productivity under elevated CO2. Proc. Natl Acad. Sci. USA 104, 14014–14019 (2007).
Merkel, D. Soil Nutrients in Glaciated Michigan Landscapes: Distribution of Nutrients and Relationships with Stand Productivity. (Doctoral Thesis Submitted to Michigan State University, 1988).
Host, G. E. & Pregitzer, K. S. Geomorphic influences on ground-flora and overstory composition in upland forests of northwestern lower Michigan. Can. J. Res. 22, 1547–1555 (1992).
Edwards, I. P. & Zak, D. R. Phylogenetic similarity and structure of Agaricomycotina communities across a forested landscape. Mol. Ecol. 19, 1469–1482 (2010).
Schimel, D., Stephens, B. B. & Fisher, J. B. Effect of increasing CO2 on the terrestrial carbon cycle. Proc. Natl Acad. Sci. USA 112, 436–441 (2015).
Medlyn, B. E. et al. Using ecosystem experiments to improve vegetation models. Nat. Clim. Change 5, 528–534 (2015).
Peñuelas, J. et al. Shifting from a fertilization-dominated to a warming-dominated period. Nat. Ecol. Evol. 1, 1438–1445 (2017).
McClaugherty, C. A., Pastor, J., Aber, J. D. & Melillo, J. M. Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology 66, 266–275 (1985).
Pastor, J., Aber, J. D., McClaugherty, C. A. & Melillo, J. M. Aboveground production and N and P cycling along a nitrogen mineralization gradient on Blackhawk Island, Wisconsin. Ecology 65, 256–268 (1984).
Serra-Maluquer, X., Mencuccini, M. & Martínez-Vilalta, J. Changes in tree resistance, recovery and resilience across three successive extreme droughts in the northeast Iberian Peninsula. Oecologia 187, 343–354 (2018).
Vitousek, P. Nutrient cycling and nutrient use efficiency. Am. Nat. 119, 553–572 (1982).
Darrouzet-Nardi, A., Ladd, M. P. & Weintraub, M. N. Fluorescent microplate analysis of amino acids and other primary amines in soils. Soil Biol. Biochem. 57, 78–82 (2013).
Ibáñez, I., Zak, D. R., Burton, A. J. & Pregitzer, K. S. Anthropogenic nitrogen deposition ameliorates the decline in tree growth caused by a drier climate. Ecology 99, 411–420 (2018).
Lines, E. R., Zavala, M. A., Purves, D. W. & Coomes, D. A. Predictable changes in aboveground allometry of trees along gradients of temperature, aridity and competition. Glob. Ecol. Biogeogr. 21, 1017–1028 (2012).
Taylor, D. L. et al. Accurate estimation of fungal diversity and abundance through improved lineage-specific primers optimized for illumina amplicon sequencing. Appl. Environ. Microbiol. 82, 7217–7226 (2016).
Callahan, B. J. et al. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583 (2016).
Nilsson, R. H. et al. The UNITE database for molecular identification of fungi: handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 47, D259–D264 (2019).
Konar, A. et al. High-quality genetic mapping with ddRADseq in the non-model tree Quercus rubra. BMC Genomics 18, 417 (2017).
Sork, V. L. et al. First draft assembly and annotation of the genome of a California Endemic oak. Genes|Genomes|Genet. 6, 3485–3495 (2016).
Wood, D. E., Lu, J. & Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 20, 257 (2019).
Buchfink, B., Xie, C. & Huson, D. H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60 (2015).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Treiber, M. L., Taft, D. H., Korf, I., Mills, D. A. & Lemay, D. G. Pre- and post-sequencing recommendations for functional annotation of human fecal metagenomes. BMC Bioinforma. 21, 74 (2020).
Peng, M. et al. Comparative analysis of basidiomycete transcriptomes reveals a core set of expressed genes encoding plant biomass degrading enzymes. Fungal Genet. Biol. 112, 40–46 (2018).
Floudas, D. et al. Uncovering the hidden diversity of litter-decomposition mechanisms in mushroom-forming fungi. ISME J. https://doi.org/10.1038/s41396-020-0667-6 (2020).
Kriventseva, E. V. et al. OrthoDB v10: sampling the diversity of animal, plant, fungal, protist, bacterial and viral genomes for evolutionary and functional annotations of orthologs. Nucleic Acids Res. 47, D807–D811 (2019).
Quinn, T. P., Erb, I., Richardson, M. F. & Crowley, T. M. Understanding sequencing data as compositions: an outlook and review. Bioinformatics 34, 2870–2878 (2018).
Ferrier, S., Manion, G., Elith, J. & Richardson, K. Using generalized dissimilarity modelling to analyse and predict patterns of beta diversity in regional biodiversity assessment. Divers. Distrib. 13, 252–264 (2007).
Duhamel, M. et al. Plant selection initiates alternative successional trajectories in the soil microbial community after disturbance. Ecol. Monogr. 89, e01367 (2019).
Qin, C., Zhu, K., Chiariello, N. R., Field, C. B. & Peay, K. G. Fire history and plant community composition outweigh decadal multi-factor global change as drivers of microbial composition in an annual grassland. J. Ecol. 108, 611–625 (2020).
Oksanen, J., et al. Package vegan.
Wickham, H. et al. Welcome to the Tidyverse. J. Open Source Softw. 4, 1686 (2019).
Spiegelhalter, D. J., Best, N. G., Carlin, B. P. & Linde, A. V. D. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. B Stat. Methodol. 64, 583–639 (2002).
Acknowledgements
We especially thank R. Upchurch, E. Herrick, K. Seguin, N. Gudal, N. Ahmad and B. VanDusen valuable laboratory and field support. V. Denef, M. Coon, J. Evans and T. James provided essential sequencing and bioinformatic support.
Author information
Authors and Affiliations
Contributions
P.P., I.I. and D.Z. designed the study. P.P. and I.I. performed the analyses. W.A. and K.A. contributed intellectually to this project as well as to field and laboratory work. P.P drafted the paper, and all authors contributed to the final copy.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Communications thanks Mohammad Bahram and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pellitier, P.T., Ibáñez, I., Zak, D.R. et al. Ectomycorrhizal access to organic nitrogen mediates CO2 fertilization response in a dominant temperate tree. Nat Commun 12, 5403 (2021). https://doi.org/10.1038/s41467-021-25652-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-021-25652-x
This article is cited by
-
Role of ectomycorrhizal colonization in enhancement of nutrients for survival of plants collected from mountainous cold stress areas
BMC Microbiology (2024)
-
A risk assessment framework for the future of forest microbiomes in a changing climate
Nature Climate Change (2024)
-
Fungal community composition predicts forest carbon storage at a continental scale
Nature Communications (2024)
-
Microbes modify soil nutrient availability and mediate plant responses to elevated CO2
Plant and Soil (2023)
-
Conversion of native grassland to coniferous forests decreased stocks of soil organic carbon and microbial biomass
Plant and Soil (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
