
Abstract
Genetically modified conditional lethal strains have been created to improve the control of insect pest populations damaging to human health and agriculture. However, understanding the potential for the genetic breakdown of lethality systems by rare spontaneous mutations, or selection for inherent suppressors, is critical since field release studies are in progress. This knowledge gap was addressed in a Drosophila tetracycline-suppressible embryonic lethality system by analyzing the frequency and structure of primary-site spontaneous mutations and second-site suppressors resulting in heritable survivors from 1.2 million zygotes. Here we report that F1 survivors due to primary-site deletions and indels occur at a 5.8 × 10−6 frequency, while survival due to second-site maternal-effect suppressors occur at a ~10−5 frequency. Survivors due to inherent lethal effector suppressors could result in a resistant field population, and we suggest that this risk may be mitigated by the use of dual redundant, albeit functionally unrelated, lethality systems.
Similar content being viewed by others

Introduction
Genetically modified (GM) conditional lethal insect strains have been developed to improve the sterile insect technique (SIT), with a focus on eliminating females during rearing for sexing, and genetic sterilization of males due to inviable progeny1,2. Two systems achieve non-sex-specific and female-specific conditional lethality by using the Escherichia coli tetracycline (Tet) resistance operon3 to suppress expression (Tet-off)4 of a lethal effector by feeding the antibiotic during rearing. One system, known as RIDL (release of insects carrying a dominant lethal), uses toxic overexpression of the tet-transactivator (tTA) in a self-promoting driver/effector cassette to effect lethality in the absence of Tet5, while a Tet-off embryonic lethality system (TELS) uses a developmentally regulated tTA driver to promote the lethal effector expression of a pro-apoptotic cell death gene in embryos6. Both systems are highly effective in small-scale experimental studies in several insect species7,8,9,10, and the RIDL system has advanced to field release studies for Aedes aegypti11,12. However, in laboratory control studies for RIDL, survival in the medfly, Ceratitis capitata, and the yellow fever mosquito, A. aegypti, were reported at frequencies of 0.5% and 3.5%, respectively7,9, while survival for the Drosophila melanogaster TELS strain was approximately 0.01%6. These levels of initial F1 survival are likely due, primarily, to inherent “leakiness” in the respective systems due to variable transgenic lethal effector expression or function, though heritable survival due to mutations in genetic components of the system have yet to be reported. This is due, in part, to laboratory screening of lethality lines being limited to thousands of adults while spontaneous mutations in Drosophila, resulting in loss-of-function alleles, have been reported in the range of 1–5 × 10−6 per locus per generation13. Thus, if mass-reared GM conditional lethal strains are released in the field at current levels of 106–108 insects/week for SIT14,15, there is a high likelihood that rare primary- or second-site mutations will occur resulting in lethal revertant survivors. This would result in the persistence of GM insects in the field, and for some mutations (especially at second sites), revertants may be resistant to further control by the same or similar lethality system. Indeed, a recent report of A. aegypti populations in Brazil after the release of RIDL mosquitoes indicates introgression of RIDL strain genomic sequences in the surviving population16, which could be associated with inheritance from non-mutant F1 escapers or with inherent resistance to the tTA lethal effector allowing hybrid survival.
To experimentally assess the potential for breakdown of Tet-suppressible lethality, we have tested the frequency and genetic basis of heritable lethal revertant survivors for a D. melanogaster non-sex-specific TELS strain reared on Tet-free diet. Despite not being a significant economically important species, its use as a genetic model provides relevant knowledge relating to the frequency and structure of spontaneous mutations, and it is highly amenable to large-scale laboratory rearing. The binary driver/effector TELS system also provides the same primary tTA target site for mutation that is necessary for lethality in both the TELS and RIDL systems, thus allowing potential breakdown of both Tet-off conditional lethal systems to be evaluated within the same genomic context. In this respect, after screening approximately 1.2 million zygotes on Tet-free diet, 20 heritable lethal revertant survivors were discovered, 7 of which are due to primary-site spontaneous mutations in the driver/effector cassettes while the remaining 13 are due to second-site maternal effect suppressors. Primary-site survivors could still be controlled by the lethality system in the field, though second-site lethal effector suppressors are likely to be resistant. We suggest that the risk of insects resistant to the primary lethality system survive and expand in a field population may be mitigated by the concurrent use of a secondary redundant lethality system that is functionally independent.
Results
Evaluation of a Tet-off embryo-specific lethality line
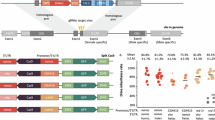
The TELS strain tested included the original piggyBac lethal effector vector, pB{3xP3-ECFP-5’HS4>5’HS4-TREp-hidAla5-5’HS4>5’HS4} (line M5.II) as described by Horn and Wimmer6 (Fig. 1a). This vector is comprised of a phospho-mutated hidAla5 pro-apoptotic cell death lethal effector gene17 linked to a tetracycline response element (TRE), surrounded by two duplicate pairs of 5’ HS4 chicken β-globin insulator sequences18, and the 3xP3-ECFP marker gene (Fig. 1b). The piggyBac serendipity α-regulated tTA driver vector, pB{PUb-DsRed.T3, s1-tTA} (line M18A), was modified from the original embryonic tTA driver (s2-tTA)6,19 by use of the s1 promoter that has an additional 95 upstream promoter nucleotides (Fig. 1a) and includes the polyubiquitin-regulated DsRed.T3 fluorescent marker (Fig. 1b). Both vectors were integrated into the second chromosome of independent D. melanogaster white- mutant host strains, recombined for common linkage, and then inbred to create the double-homozygous driver/effector strain, DH-1.

a Structure of the pBXL{attP_PUbDsRed.T3_fa_s1-tTA_a} tTA driver and pBac{3xP3-ECFPaf; 5’HS4>5’HS4>TREp-hidAla5>5’HS4>5’HS4} lethal effector vectors in the double-homozygous line, DH-1. b Images of an adult DH-1 male and w[m] female under brightfield (BF) and Texas Red (DsRed) and CFP (ECFP) epifluorescence optics. DH-1 males, having a w[m] background, exhibit the white- eye phenotype under brightfield and whole-body PUbDsRed and eye-specific 3xP3-ECFP fluorescent markers for the driver and effector vectors, respectively. Non-transgenic w[m] females exhibit the white- eye phenotype under brightfield and are non-fluorescent for both markers.
The applied use of TELS in the field would require the mass release of double-homozygous P1 males reared on Tet-diet, with the expectation that their double-heterozygous F1 progeny would be inviable by the first larval instar in the absence of Tet. Thus F1 survivors would be considered to be putative lethal revertants due to spontaneous primary-site point mutations, deletions or insertions in the driver or effector cassettes, or second-site modifiers or variations in the genome that would somehow suppress the otherwise functional TELS. However, fertile adult survivors expressing the DsRed driver (Red) and ECFP effector (Cyan) cassette markers would require verification as true heritable lethal revertants from a backcross to w[m] that yields double-marked Red/Cyan F2 surviving progeny on Tet-free diet (Fig. 1b).
F1 survivors from large-scale rearing
Based on typical spontaneous mutation rates of 10−6 to 10−5 per locus in Drosophila13, we presumed that the recovery of a significant number of heritable lethal revertants from the two loci TELS strain would require a minimal screen of ~106 F1 adults from large-scale rearing. This was achieved by establishing 6 control mating groups, each having 100 homozygous DH-1 males crossed to 400 w[m] virgin females reared on permissive Tet-diet, with two successive transfers to fresh media, which yielded a total of ~15,000 F1 adult progeny from the three broods for each mating group (Table 1). The same crosses were then established for 80 experimental mating groups reared on non-permissive Tet-free diet that would account, based on control matings, for the screening of approximately 1,200,000 double-heterozygous F1 progeny for adult survival. From the experimental matings, a total of 109 Red/Cyan (R/C) double-marked adults, for the driver and lethal effector vector cassettes (Fig. 1b), eclosed on Tet-free diet yielding an initial survival rate of 0.0091% (Table 1), consistent with preliminary small-scale population survival rates.
Heritable F2 lethal revertants identified from F1 survivors
From the 109 F1 survivors, 88 remained viable allowing backcross matings to w[m] to test for heritability of disruption or suppression of the lethality system (Table 1). Seventy-three F1 survivors were fertile, of which 20 resulted in R/C F2 progeny on Tet-free diet, indicating a heritable survival frequency of 18.3% relative to the initial number of survivors, and a survival frequency of 1.67 × 10−5 relative to the total number of adults screened (Table 1). Seven of the 20 survivor lines exhibited an approximate 1:1 ratio of unmarked to double-heterozygous R/C marked F2 descendants on Tet-diet, with approximately 11% of the progeny expressing either the R or C marker (Table 2). Independent marker segregation was presumably due to recombination between the driver and lethal effector cassettes at their respective chromosomal 2R loci of 56C8/9 and 53C4 (Supplementary Fig. 1), limited to the five F1 female lines due to the absence of male recombination in D. melanogaster20. These results were consistent with complete disruption of the lethality system due to spontaneous primary-site genetic alterations in either, or both, the tTA driver or lethal effector cassettes at a frequency, relative to the population screened, of 5.8 × 10−6 events per generation. The significance of this frequency is supported by a power calculation21 to test for the optimal sample size required for detecting primary-site genetic alterations based on the typical null mutation rate of 10−6. This resulted in a power of 0.93 at the 0.05 significance level for the screened sample size of ~1,200,000 double-heterozygous F1 progeny (Supplementary Fig. 2).
Second-site maternal effect lethality suppression lines
The 13 additional F2 R/C survivor lines were heritable but exhibited significantly lower survival frequencies for R/C marked progeny relative to their unmarked siblings (Table 2), ranging from 1.4% to 32.4% survival, with 11 lines being <11%. These variable R/C frequencies are more consistent with lethal suppression by second-site modifiers, either distantly or un-linked to the driver/effector cassettes, and possibly the result of different suppressors.
Notably, all of these lines originated from F1 females, although F2 survivors included both sexes, suggesting the possibility of maternal effect heritability of lethal suppression. This was tested by backcrosses of R/C revertant males and females to w[m] flies in successive generations. For all 13 lines, F3 R/C survivors only arose on Tet-free diet from maternal R/C F2 females, and not from R/C F2 males, while both F2 males and females gave rise to F3 R/C progeny on Tet-diet. For two lines (5-f1 and 14-f2) having the highest levels of survival, the maternal effect was observed for 11 generations and confirmed by 10 to 12 individual backcross matings of F10 males and females to w[m] (Table 3).
Molecular analysis of lethal revertant survivor lines
To determine the molecular integrity of the tTA driver and lethal effector cassettes in the lethal revertant survivor lines, F2 R/C adults for each line were inbred to homozygosity on Tet-free diet. PCR products from the s1-tTA driver and TREp-hidAla5 lethal effector cassettes from the revertant lines were sequenced (Supplementary Table 2 and Supplementary Fig. 3) using adjacent and internal primers, though reliable PCR amplification of the highly repetitive 5’ HS4 insulator sequences was not possible. For some difficult sequences, more distant primers or TAIL-PCR were utilized. For the seven putative primary-site revertant lines, relatively short deletions were identified within the tTA and hidAla5 genes, in addition to larger deletions and indels that included TREp-hidAla5 and adjacent insulators (Fig. 2). Notably, mutations or alterations were not evident in the driver or effector cassettes for any of the 13 putative modifier lines, consistent with second-site mutations or variations.

a Line 18-f1 has a 27-bp deletion and 3 nucleotide substitutions (blue vertical lines and nucleotides) in the tTA gene and b line 45-f1 has a 26-bp deletion in the hidAla5 cell death gene (maroon dashes and triangles indicate deleted sequences). Lines 9-f1, 62-f1, and 65-m2 have identical deletions of the TREp-hidAla5 lethal effector and two 5’ HS4 insulator sequences, lines 44-m1 and 21-f1 exhibit deletions (blue dotted lines), and insert sequences (blue solid lines) from within the ECFP-linked SV40 extending downstream through the 3’ genomic insertion site sequence (blue arrows indicate unknown indel termination sites).
For the primary-site mutations, two relatively short spontaneous deletions were detected in lines 18-f1 and 45-f1. In line 18-f1, the TREp-hidAla5 lethal effector sequence was unaltered, but a 27-bp deletion (nts 221–247 from the ATG) and three adjacent nucleotide substitutions were identified in the tTA driver coding sequence (Fig. 2 and Supplementary Fig. 4a). While the tTA reading frame was maintained, we presume the deleted and substituted peptides resulted in loss of tTA function. In line 45-f1, the tTA driver was unaltered, but a 26-bp deletion (nts 27–52 from the ATG) occurred in the 1.2-kb hidAla5 lethal effector gene, resulting in an early frameshift that likely eliminated gene product function (Fig. 2 and Supplementary Fig. 4b).
For the primary-site lines 21-f1 and 44-m1, the tTA driver sequence was unaltered, but a series of PCR primer pairs flanking or internal to the 5’HS4_5’HS4-TREp-hidAla5-5’HS4_5’HS4 lethal effector cassette failed to yield amplicons, consistent with a large deletion or indel (see “Methods” and Supplementary Fig. 3b, c2–7). Control PCRs were positive for the unaltered presence of the pB3’ to 3xP3-ECFP sequence (AH1577–AH1636) in both lines (Supplementary Fig. 3b, c1) consistent with the expression of the marker gene. Internal PCR (AH1540–AH1652) indicated the presence of the effector pB5’ sequence in both lines (Supplementary Fig. 3b, c8); however, PCR from pB5’ to adjacent 3’ genomic sequences were not positive (Supplementary Fig. 3b, c6) suggesting a transposition or rearrangement of sequences. Partial insert sequences downstream from ECFP-SV40 were identified by TAIL-PCR, which included a 180-bp incomplete insert of unknown origin in line 21-f1 (Supplementary Fig. 5a), except for several short sequences identical to those found on the X and 2R chromosomes, the 3xP3 promoter, and a 5’ HS4 insulator. Line 44-m1 revealed a 540-bp incomplete insert (Supplementary Fig. 5b) having high identity to telomeric HeT-A non-LTR retrotransposon sequences, with 87% identity to the X chromosome HeT element (ID: M81595.1)22 (Supplementary Fig. 6). Notably, retrotransposon insertions, among other mobile elements, have been associated with deletions that may provide the breakpoints for their insertion resulting in spontaneous mutations in Drosophila and other eukaryotes23,24.
For the three remaining heritable survivor lines (9-f1, 62-f1, and 65-m2), PCR with primers in ECFP and pB5’ and ECFP-linked SV40 and pB5’ yielded 2.9 kb (Supplementary Fig. 3c7) and 2.6 kb (Supplementary Fig. 7a) products, respectively, that included partial 5’ HS4 insulator sequences and the absence of the internal TREp-hidAla5 sequence. The actual sequence in this region was then inferred by AflII and BglII digests of the 2.6-kb product that resulted in fragment sizes consistent with a deletion of the lethal effector and two insulators (Supplementary Fig. 7a). If the two remaining insulator sequences were originally upstream and downstream of TREp-hidAla5, a parsimonious explanation for the deletion would be a single-strand inverted-loop meiotic pairing of the duplicate insulator tandem pairs having a single internal recombination event. The resulting segregation products would thus consist of the two remaining insulators due to a circular DNA deletion of 5’HS4_TREphidAla5_5’HS4 (Supplementary Fig. 7b).
Discussion
After nearly 20 years since Tet-off conditional lethality systems for pest insect population control were first reported5,6,25, a quantitative and structural assessment for the spontaneous breakdown of the genetic components that comprise the D. melanogaster Tet-off embryonic lethality system has now been described. After screening approximately 1.2 million heterozygous TELS F1 adults on a restrictive Tet-free diet, 20 heritable revertant lethal survivors were recovered, yielding a frequency of 1.7 × 10−5 events per generation resulting in genetic breakdown. However, primary-site mutations disrupting the tTA driver or hidAla5 lethal effector sequences were verified for seven of the survivor lines that included deletions and indels of various length, including a retrotransposon-associated indel, resulting in a spontaneous primary-site mutation rate of approximately 5.8 × 10−6 events per generation.
Given that three of the lethal revertant lines were likely due to deletions resulting from cis-recombinations between the highly repetitive 5’ HS4 chicken globin insulators, a more relevant revertant frequency that includes only the remaining four mutant lines would be ~3.3 × 10−6 events per generation (Table 1). Since this mutation rate includes genetic alterations from both primary-site targets, it more accurately reflects the overall rate of genetic breakdown for this specific lethality system, whereas the approximate spontaneous mutation rate per locus for the 1.0-kb tTA driver and 1.2-kb hidAla5 lethal effector coding regions would be 0.83 × 10−6 and 2.5 × 10−6 events per locus, respectively. Various factors can affect the mutation rate of specific loci, including the method of phenotypic screening and, indeed, our revertant lethal screens depended on detecting dual DsRed/ECFP marked survivors that would have missed revertants due to deletions including critical sequences for either marker gene. With these caveats in mind, the observed spontaneous mutation rates for the driver and effector alleles remain generally consistent with previous spontaneous mutation studies for a variety of alleles in Drosophila, having a mean spontaneous specific locus rate of 1–5 × 10−6 as estimated by Ashburner13 and additional analyses24,26,27,28,29.
The recovery of putative insulator recombinations also highlights an important consideration for the design of lethality systems since the insulator pairs surrounding both driver and effector cassettes in the original TELS strain did improve lethal gene expressivity6. However, this was achieved at the cost of increasing the primary-site mutation frequency by >40% (Table 1). Thus an important caveat for the use of the 5’ HS4 insulator, and possibly other repetitive sequences, is the possibility for internal cis-recombinations resulting in increased lethality strain breakdown due to deletions in a significant percentage of F1 progeny.
The mutation recovered in this study most relevant to the RIDL system, for which A. aegypti RIDL lines have already been tested in open field release studies9,30, was the short internal deletion within the tTA driver that acts as both driver and lethal effector in the self-promoting RIDL system. Given the 0.83 × 10−6 mutation frequency of this locus from a single F1 TELS revertant, it is possible to expect a RIDL insect to survive from at least every 1.2 million F1 embryos oviposited in the field. Not unexpectedly, the additional hidAla5 effector cassette critical for TELS function, compared to RIDL, provides an additional target for mutation and thus a higher frequency for disruption of lethality based on this study.
In addition to primary-site mutations, this study revealed a class of second-site maternal effect variations, found in 13 of the revertant lethality lines, that effectively suppress expression or function of the hidAla5 pro-apoptotic lethal effector gene. While it remains to be determined how ectopic hidAla5 could be negatively influenced by a maternally derived factor, there is precedence for a maternally inherited mitochondrial role in pro-apoptotic gene product function. In mammalian systems, cytochrome c is converted into holocytochrome c by a heme attachment within the mitochondrial intermembrane space that facilitates pro-apoptotic caspase activation resulting in cell death31. While this specific function has not been observed in invertebrates, a mitochondrial role in Drosophila cell death has been inferred by the inhibition of apoptosis when pro-apoptotic protein localization within the mitochondria is blocked32. Although the observed maternal effect may be limited to the particular D. melanogaster w[m] host strain tested, if ectopic pro-apoptosis is similarly subject to suppressor gene function due to variation in a small subset of an insect population, then the potential for selection of these suppressors should be a concern for the use of pro-apoptotic lethal effectors. Indeed, the hid gene has already been tested as a TELS conditional lethal effector in four insect pest species8,10,33,34,35, some of which are in consideration for release.
Another mechanism for maternal effect suppression, since the sry-alpha promoter functions transiently in early embryogenesis19, is a pre-zygotic maternal contribution of molecules that suppress ectopic tTA or hidAla5 expression or function at that developmental period. Since the RIDL system relies on the toxic accumulation of tTA protein throughout development for lethality, neither of these types of maternal effect suppression may be relevant to RIDL, though it is possible that tTA toxicity ultimately results in induced apoptosis.
More broadly, however, is the realization that regardless of the lethal effector employed for population control, or its mode of action, it may be subject to suppression by a pre-existing inherent variation in the targeted field population36 or a newly acquired modifier mutation in the mass-reared lethality strain. In either case, such effects could have a significant impact on lethality system effectiveness and implications for its use in population control programs. Importantly, this would require evaluation by contained large-scale preliminary testing as performed in this study but, ideally, using field population adults for P1 matings. Another important consideration is that primary-site revertant survivors, due to cis-acting mutations, of both TELS and RIDL systems should still be susceptible to control by the same lethality system. In contrast, a dominant-acting second-site suppressor of lethal effector expression or function, due to trans-acting modifiers, would more likely result in GM insects resistant to the lethality system, and relatively small numbers of GM resistant survivors could be expected to rapidly re-populate an ecosystem where the susceptible population has been suppressed. Certain factors will affect the rate of introgression of the resistant population and its ability to reach fixation. These include the number of suppressor alleles selected for and whether they are recessive lethal, recessive or dominant fitness costs, maternal effect heritability, and the proportional population density of successive releases among other variables having positive or negative effects37. All of the TELS suppressor lines were inbred to homozygosity (and share the same mitochondrial genome), and none exhibited recessive lethality or apparent semi-lethality due to compromised fitness. Nevertheless, unless the negative costs to resistance are considerable, the eventual establishment of a lethality-resistant population may be expected from programs that rely on continuous mass releases (especially preventative release programs38) in the absence of an efficient secondary control program. The potential for resistance to bi-sexual and female-specific RIDL release programs, and its spread within a population, has been modeled taking into account many of the variables affecting resistance introgression for this system37, but this remains theoretical in the absence of large-scale population tests and knowledge of the actual basis for tTA lethality. The importance of this knowledge for the RIDL system, in particular, is heightened by recent studies showing significant introgression of genomic sequences from the A. aegypti OX513A RIDL strain into the native Brazilian (Jacobina) mosquito population 6 months to 2.5 years after the initiation of lethal strain releases16. Notably, sequences related to the lethality system were not reported, nor could they necessarily be identified by the single-nucleotide polymorphism analysis performed, and the sequences identified could have simply been inherited from non-mutant escapers that comprised >70% of the F1 TELS survivors in our tests. But until this is resolved, one explanation for at least some of the observed introgression is survival due to inherent resistance to the tTA lethal effector. Alphey et al. suggest early and effective monitoring of release programs to detect resistance in time for remediation37, which is certainly important on a continuing basis, as it is for most current SIT programs38. However, the additional use of preliminary large-scale contained population tests previous to a new release program would provide the advantage of assessing the potential for resistance and a determination as to whether concurrent or sequential releases of a secondary control system, or other modifications, would be necessary.
Considering the possibility that resistance to current insect conditional lethality systems might occur, strategies to prevent or mitigate the survival of lethality-resistant individuals have been considered. Two strategies are based on the potential for redundant, yet independent, dual lethality (or reproductive sterility) systems mitigating the survival of revertant individuals in a mass release program39,40. Thus, based on the multiplicative law of probability [P(AB) = P(A)P(B)], an approximate 10−6 lethal revertant frequency estimate for each system (A and B), as demonstrated here for a Drosophila TELS strain, could result in a frequency as low as ~10−12 for both to fail concurrently in the same genome, presuming that the lethality systems are functionally independent and do not differentially affect host physiology and fitness. In application, at a modest rate of release for current SIT programs (~107 males per week), use of a single lethality system might result in ≥100 heritable revertant survivors per week assuming each male yields 10 F1 progeny, whereas the coincident breakdown of a dual lethality strain might yield a single revertant survivor only after several decades.
From these considerations, we would suggest that large-scale population screens for spontaneous mutations or pre-existing genetic variations, resulting in the genetic breakdown of conditional lethal systems developed for mass field release, be a critical component in the risk assessment evaluation of these programs. This could be achieved most efficiently in rearing facilities where mass-reared P1 males and females are mated under non-permissive conditions for survival, whereupon the frequency and structure of mutations and suppressor/modifiers could be evaluated in surviving F1 progeny. Given that most current facilities for SIT mass-rearing generate a minimum of 107–108 insects per week, a more significant and informative analysis could be achieved in insects of interest relative to large-scale laboratory studies with Drosophila. Importantly, host females for P1 matings should come from the targeted field populations to test for inherent suppressors or modifiers of the lethality system as a prelude to actual releases, though such evaluations may prove to be less critical with the use of redundant lethality systems.
This study has focused on a conditional lethal strain created by transposon-mediated transformation, yet most insect genetic modifications should be subject to the same types of alteration, disruption, or suppression regardless of the methodology used to create them. However, the frequency of specific types of mutations and suppressor/modifiers may vary with the genomic structure of the host species (e.g., genome size, chromatin structure, repetitive DNA content, etc.) and the specific modification. This is especially relevant to the variety of manipulations envisioned by gene editing, and the development of gene drive (GD) systems in particular. Indeed, despite the great potential for GD systems to modify or replace highly pestiferous insect strains in the field, it is well recognized that control or containment systems must be integrated into GD systems before the field release of these self-propagating strains can be considered41,42. A variety of containment strategies proposed, and in some cases tested, include drives that are self-limiting43 such as multi-component daisy-chained drives44, split drives45,46 and spatially restricted drives45 and small molecule-dependent drives47, among others. However, similar to the lethality strain breakdown demonstrated here all of these strategies should be subject to mutation and modification and, potentially, at higher frequencies due to multi-generational GD strategies and larger target sites for some. Thus it would seem imperative that similar large-scale studies for GD integrity be initiated for systems already created in Drosophila, and if warranted, redundancy for GD containment be considered and tested for mitigation. Similarly, the desired systems for introgression should be equally subject to genetic breakdown for which redundancy may also be required, at the risk of a GD population regaining its pestiferous function, or losing its beneficial function, and expanding in nature. In this respect, and in general, care should be taken in the design of all genetic modifications for specific applications, in terms of understanding and testing the potential for mutation, suppression, and the possible need for redundancy to maintain strain stability.
Methods
Insect strains, rearing, and screening
All D. melanogaster transgenic lines, the white-eyed w[m] mutant line, and large-scale matings were reared under standard laboratory conditions at 25 °C and 60% humidity on a 12-h light:12-h dark cycle48. Flies were anesthetized with CO2 for screening, fly collections, and matings. Transgenic flies were screened by epifluorescence microscopy using a Leica MZFLIII fluorescence microscope and filter sets, TxRed for DsRed (ex: 560/40; em: 610 LP) and CFP for ECFP (ex: 436/20; em: 480/40). The University of Florida Institutional Biosafety Committee provided oversight and regulatory approval for the creation and rearing of the GM D. melanogaster strains used in this study.
Tet-off tTA driver and hid Ala5 lethal effector lines
The D. melanogaster transgenic embryonic tTA driver M18A strain was transformed with the piggyBac transgene vector, pBXL{attP_PUbDsRed.T3_fa_s1-tTA_a} (GenBank accession no.: MT453110). The vector was created by isolating the s1-tTA driver sequence, as an AscI fragment, from pBac{3xP3-EYFPafm; s1-tTA}6 and inserting it into the AscI site in pBXL{attP_PUbDsRed_fa}10. Unlike the tTA driver vector used in the original embryonic lethality strain6, the serendipity α s1 promoter has an additional 95 upstream nucleotides compared to the s2 promoter, the 5’ HS4 insulator sequences that surround the s2-tTA driver construct are eliminated, an attP recombination site is added for subsequent phiC31-mediated integrations, and a polyubiquitin-regulated DsRed (PUbDsRed) whole-body marker replaces the eye-specific 3xP3-EYFP marker. The vector was transformed into the w[m] strain by piggyBac-mediated germline transformation49 using a 500 ng/µl transgene vector plasmid and 200 ng/µl phspBac transposase helper plasmid mixture for preblastoderm injections.
The transgenic M5.II lethal effector strain, pBac{3xP3-ECFPaf_5’HS>5’HS>TREp-hidAla>5’HS>5’HS4} was created and kindly provided by Horn and Wimmer6. This effector vector is marked with the eye-specific marker, 3xP3-ECFP, and carries a Tet-suppressible lethal effector construct having the phospho-mutated hidAla5 pro-apoptotic cell death gene linked to a TRE. In the absence of tetracycline, the tet-transactivator (tTA-VP16 originally isolated from the pTet-Off Vector plasmid; Takara BIO USA, Inc.) from the driver construct binds to the tetO operator sequences within the TRE to promote hidAla5 transcription. Included within the vector construct is an attP recombination site for subsequent phiC31-mediated integrations, and two tandem repeat 2.4 kb 5’ HS4 chicken β-globin insulator repeat sequences flank both sides of the effector construct.
Vector insertion site mapping
The effector cassette in strain M5.II was previously mapped to chromosome 2 but not to a specific locus6. Segregation patterns from the mating of the M18A and M5.II homozygous strains and subsequent inbreeding suggested that the effector cassette in M18A also maps to chromosome 2. Therefore, the M18A and M5.II homozygous strains were inter-mated and double-heterozygous progeny were outcrossed to w[m] to select recombinant progeny having the driver and effector cassettes linked on the same chromosome. Recombinants were inbred to create the double-homozygous driver/effector line, DH-1, that was mated to the D. melanogaster white mutant strain w[m] in large-scale rearing (Fig. 1).
Both the effector and driver cassettes in DH-1 were mapped to specific loci by inverse PCR insertion-site sequencing to one arm of each vector. Inverse PCR10 was performed by digestion of the DH-1 strain with AvaII or RsaI and amplifying circularized fragment sequences with primer sets AH1537–AH1579 for the driver cassette 3’ flanking sequence and AH1566–AH1542 for the effector cassette 5’ flanking sequence (Supplementary Table 2). The flanking genomic sequences were subjected to a BLASTn search of the D. melanogaster genome assembly indicating an insertion of the driver cassette at the Tab2 gene (CG7417) and the effector cassette at the Sema2b gene (CG33960), both on chromosome 2R (Supplementary Fig. 1). However, while both transgenes inserted into a TTAA site (duplicated upon insertion) consistent with all piggyBac insertions, the genomic sequences immediately flanking both the 5’ and 3’ piggyBac termini were not present in the D. melanogaster NCBI database. The driver cassette 5’ flanking sequence and the effector cassette 3’ flanking sequence were then re-confirmed by direct PCR using primer sets, AH1536–AH1585 and AH1543–AH1570 (Supplementary Table 2), to internal vector and genomic sequences and were also confirmed by direct internal sequencing in the genomes of w[m] and another white strain, w1118. A straightforward explanation for the anomalous sequences is that many white strains have been maintained as inbred laboratory colonies for many years (at least 60 years for w1118) and may have been subjected to genetic drift from the wild-type strains used for recent whole-genomic sequencing.
Tetracycline tests
To generate an embryonic lethality strain exhibiting the lowest survival frequency on Tet-free diet and the highest level of survival on the lowest Tet-diet concentration (considering potential fitness costs at high concentrations), F1 survival of progeny from small-scale matings of DH-1 males and w[m] females was first tested on Tet-free diet and diet supplemented with tetracycline at concentrations of 10, 50, and 100 μg/ml (Supplementary Table 1). Eggs were collected on medium containing the corresponding Tet concentration, with pupae and adult eclosion rates recorded. Since no significant difference was observed in eclosion rates between 10 and 50 μg/ml, a concentration of 20 μg/ml tetracycline was used in media for large-scale rearing tests.
Large-scale rearing tests
For large-scale rearing tests, w[m] virgin females and double-homozygous DH-1 males were collected <8 h after eclosion and reared separately for 2 days before P1 mating of 400 females and 100 males for brood I in 1-liter jars on 20 μg/ml Tet-diet for control tests and Tet-free media for experimental tests. Brood I adults were transferred to fresh media after 4 days for brood II, with another transfer after 4 days for brood III, with brood III adults discarded after 6 days. Six control matings were performed to determine the mean total number of surviving adults from the three broods for each mating, which were counted individually. This resulted in a mean number of 15,001 ± 389 standard error of the mean F1 adults per mating (Table 1). A total of 80 experimental matings (240 jars) were screened for pupal survivors that were collected and placed in individual vials with Tet-free media and monitored for adult eclosion. F1 adults were counted and screened for the fluorescent markers, and survivors double-marked for the driver (DsRed) and effector (ECFP) cassettes were backcrossed individually to w[m] adults on Tet-free diet for 3–5 days. F1 female survivor matings were then transferred to Tet-diet for 4–5 days, while three w[m] females mated to F1 male survivors were left on Tet-free diet and three females were transferred to Tet-diet. Rearing on Tet-diet ensured that a line from fertile F1 adults was maintained and provided a control for the expected percentage of Red/Cyan F2 progeny. If F2 R/C progeny survived on Tet-diet, but not on Tet-free diet, then the F1 individual was considered to be a non-heritable survivor and the line was discarded, but if the F2 R/C progeny survived on Tet-free diet, it was considered to be a putative heritable lethal revertant survivor. F2 R/C males and females were inbred to create double-homozygous lines by single sib-pair matings with selection of the strongest fluorescing progeny (presumed to be homozygous) for matings in successive generations until all progeny were strongly fluorescent, typically by the third generation. Homozygosity was then verified by backcrosses to w[m] that resulted in 100% heterozygous progeny.
Heritable lethal revertant survivor lines
Double-heterozygous F2 progeny, expressing both DsRed and ECFP (R/C) fluorescent markers, from backcrossed F1 survivors reared on Tet-free diet, were considered to be lethal revertants due to a heritable mutation or variant modifier allele. Primary-site mutations in either the driver or effector sequences, causing breakdown of the lethality system, were expected in approximately one-half of the F2 progeny with the remaining progeny consisting of unmarked w[m] flies and single-marked progeny resulting from recombination between the markers from F1 female survivors (since recombination does not occur in D. melanogaster males). Second-site modifiers were expected to have a dominant-acting function and would also be expected to suppress lethality in one-half of the double-heterozygous F2 progeny only if the alleles had relatively close second chromosome linkage to the driver/effector cassettes and were fully expressed. Lower frequencies of F2 survival would be expected if second-site modifiers were unlinked or distantly linked to the D/E cassettes or were variably expressed possibly due to independent suppressor alleles. F1 and successive generation backcrosses on Tet-diet provided a control for the expected frequencies of marked and unmarked F2 progeny. Heterozygous lethal revertant survivor lines were maintained by continuing backcrosses to w[m] on Tet-diet, in addition to inbred homozygous lines maintained on Tet-free diet for molecular analysis.
Putative second-site survivor lines, based on low F2 survival frequencies, were tested for putative maternal effect modifiers by three replicate single pair backcrosses of F2 double-heterozygous males and females to w[m] reared on Tet-free diet for 3 days and then Tet-diet for 3 days. Adult progeny on each diet were screened for markers, with the ratio of double-marked to unmarked F3 descendants compared between the two diets. F3 double-heterozygous survivors arising only from F2 females, and not from F2 males, was considered to be consistent with a second-site maternal effect modifier that was verified by continued backcrosses in successive generations and verifying that the sequence integrity of the driver and effector cassettes was unaltered (Table 2 and Supplementary Fig. 3a, b). For two putative maternal effect lines, 5f1 and 14f2, exhibiting relatively higher frequencies of survival, backcrosses were continued for 10 generations at which time 10–12 replicate single pair backcrosses of both F10 double-heterozygous males and females was performed (Table 3).
Molecular analysis of lethal revertant survivor lines
The s1-tTA driver and TREp-hidAla5 lethal effector cassette sequences were isolated by PCR for all heritable homozygous survivor lines and sequenced to identify genetic alterations that would eliminate or diminish lethal effector expression. Primer pairs AH1601–AH1631 and AH1554–AH1582 were used to amplify the driver cassette, while the primer pairs AH1616–AH1562 and AH1610–AH1581 were used to amplify the effector cassette (Supplementary Table 2 and Supplementary Fig. 3). Another primer pair, AH1566–AH1615, that amplified the effector cassette and the flanking 5′ HS4 insulator tandem repeat sequences was used if the effector cassette could not be amplified with the more proximal primers. Owing to the highly repetitive 5′ HS4 sequence, the use of primers within the sequence or PCR through the sequence with external primers was not always reliable. PCR cycling conditions were: 30 s at 98 °C; 5 cycles of 10 s at 98 °C, 30 s at 58 °C, 2 min at 72 °C; 30 cycles of 10 s at 98 °C, 30 s at 55 °C, 2 min at 72 °C; and a final extension for 2 min at 72 °C. If primer pairs flanking the driver or effector cassettes failed to generate amplicons, possibly due to large indel aberrations, TAIL-PCR was performed according to Liu and Whittier50 using specific primers (SPs) SP1 (AH1557), SP2 (AH1635), SP3 (AH1638), and an arbitrary degenerate primer (AH1117). All PCR reactions and PCR product digestions resulting in the same profile or sequence were replicated a minimum of two or more times.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The data and materials that support the findings of this study are available from the corresponding author upon reasonable request. The piggyBac driver vector, pBXL{attP_PUbDsRed.T3_fa_s1-tTA_a}, annotated sequence is available from the NCBI GenBank database, accession no. MT453110.
References
Knipling, E. F. Sterile-male method of population control: successful with some insects, the method may also be effective when applied to other noxious animals. Science 130, 902–904 (1959).
Handler, A. M. Prospects for using genetic transformation for improved SIT and new biocontrol methods. Genetica 116, 137–149 (2002).
Gossen, M. & Bujard, H. Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc. Natl Acad. Sci. USA 89, 5547–5551 (1992).
Bello, B., Resendez-Perez, D. & Gehring, W. J. Spatial and temporal targeting of gene expression in Drosophila by means of a tetracycline-dependent transactivator system. Development 125, 2193–2202 (1998).
Alphey, L. Re-engineering the sterile insect technique. Insect Biochem. Mol. Biol. 32, 1243–1247 (2002).
Horn, C. & Wimmer, E. A. A transgene-based, embryo-specific lethality system for insect pest management. Nat. Biotechnol. 21, 64 (2003).
Gong, P. et al. A dominant lethal genetic system for autocidal control of the Mediterranean fruitfly. Nat. Biotechnol. 23, 453 (2005).
Schetelig, M. F., Caceres, C., Zacharopoulou, A., Franz, G. & Wimmer, E. A. Conditional embryonic lethality to improve the sterile insect technique in Ceratitis capitata (Diptera: Tephritidae). BMC Biol. 7, 4 (2009).
Harris, A. F. et al. Field performance of engineered male mosquitoes. Nat. Biotechnol. 29, 1034 (2011).
Schetelig, M. F. & Handler, A. M. Strategy for enhanced transgenic strain development for embryonic conditional lethality in Anastrepha suspensa. Proc. Natl Acad. Sci. USA 109, 9348–9353 (2012).
Harris, A. F. et al. Successful suppression of a field mosquito population by sustained release of engineered male mosquitoes. Nat. Biotechnol. 30, 828 (2012).
Lacroix, R. et al. Open field release of genetically engineered sterile male Aedes aegypti in Malaysia. PLoS ONE 7, e42771 (2012).
Ashburner, M. Drosophila. A Laboratory Handbook (Cold Spring Harbor Laboratory Press, 1989).
Robinson, A. S. Genetic sexing strains in the medfly, Ceratitis capitata: development, mass rearing and field application. Trends Entomol. 2, 81–104 (1999).
Carvalho, D. O. et al. Mass production of genetically modified Aedes aegypti for field releases in Brazil. J. Vis. Exp. e3579 (2014).
Evans, B. R. et al. Transgenic Aedes aegypti mosquitoes transfer genes into a natural population. Sci. Rep. 9, 1–6 (2019).
Grether, M. E., Abrams, J. M., Agapite, J., White, K. & Steller, H. The head involution defective gene of Drosophila melanogaster functions in programmed cell death. Genes Dev. 9, 1694–1708 (1995).
Chung, J. H., Whiteley, M. & Felsenfeld, G. A 5′ element of the chicken β-globin domain serves as an insulator in human erythroid cells and protects against position effect in Drosophila. Cell 74, 505–514 (1993).
Ibnsouda, S., Schweisguth, F., de Billy, G. & Vincent, A. Relationship between expression of serendipity alpha and cellularisation of the Drosophila embryo as revealed by interspecific transformation. Development 119, 471–483 (1993).
McKee, B. D., Yan, R. & Tsai, J. Meiosis in male Drosophila. Spermatogenesis 2, 167–184 (2012).
Ryan, T. Sample Size Determination and Power (Wiley, Hoboken, 2013).
Danilevskaya, O., Slot, F., Pavlova, M. & Pardue, M. Structure of the Drosophila HeT-A transposon: a retrotransposon-like element forming telomeres. Chromosoma 103, 215–224 (1994).
Vogt, J. et al. SVA retrotransposon insertion-associated deletion represents a novel mutational mechanism underlying large genomic copy number changes with non-recurrent breakpoints. Genome Biol. 15, R80 (2014).
Watanabe, Y., Takahashi, A., Itoh, M. & Takano-Shimizu, T. Molecular spectrum of spontaneous de novo mutations in male and female germline cells of Drosophila melanogaster. Genetics 181, 1035–1043 (2009).
Heinrich, J. C. & Scott, M. J. A repressible female-specific lethal genetic system for making transgenic insect strains suitable for a sterile-release program. Proc. Natl Acad. Sci. USA 97, 8229–8232 (2000).
Tobari, Y. N. & Kojima, K. A study of spontaneous mutation rates at ten loci detectable by starch gel electrophoresis in Drosophila melanogaster. Genetics 70, 397–403 (1972).
Mukai, T. & Cockerham, C. C. Spontaneous mutation rates at enzyme loci in Drosophila melanogaster. Proc. Natl Acad. Sci. USA 74, 2514–2517 (1977).
Woodruff, R. C. & Thompson, J. N. Jr Genetic factors that affect rates of spontaneous mutation and chromosome aberrations in Drosophila melanogaster. Cytogenet. Genome Res. 33, 152–159 (1982).
Yang, H. P., Tanikawa, A. Y. & Kondrashov, A. S. Molecular nature of 11 spontaneous de novo mutations in Drosophila melanogaster. Genetics 157, 1285–1292 (2001).
Winskill, P. et al. Dispersal of engineered male Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 9, e0004156 (2015).
Wang, C. & Youle, R. J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 43, 95–118 (2009).
Abdelwahid, E. et al. Mitochondrial disruption in Drosophila apoptosis. Dev. Cell 12, 793–806 (2007).
Schetelig, M. F. & Handler, A. M. A transgenic embryonic sexing system for Anastrepha suspensa (Diptera: Tephritidae). Insect Biochem. Mol. Biol. 42, 790–795 (2012).
Schetelig, M. F., Targovska, A., Meza, J. S., Bourtzis, K. & Handler, A. M. Tetracycline‐suppressible female lethality and sterility in the Mexican fruit fly, Anastrepha ludens. Insect Mol. Biol. 25, 500–508 (2016).
Yan, Y., Linger, R. J. & Scott, M. J. Building early-larval sexing systems for genetic control of the Australian sheep blow fly Lucilia cuprina using two constitutive promoters. Sci. Rep. 7, 2538 (2017).
Knudsen, K. E. et al. Genetic variation and potential for resistance development to the tTA overexpression lethal system in insects. G3 (Bethesda) 10, 1271–1281 (2020).
Alphey, N., Bonsall, M. B. & Alphey, L. Modeling resistance to genetic control of insects. J. Theor. Biol. 270, 42–55 (2011).
Hendrichs, J., Robinson, A. S., Cayol, J. P. & Enkerlin, W. R. Medfly areawide sterile insect technique programmes for prevention, suppression or eradication: the importance of mating behavior studies. Florida Entomol. 85, 1–13 (2009).
Eckermann, K. N. et al. Perspective on the combined use of an independent transgenic sexing and a multifactorial reproductive sterility system to avoid resistance development against transgenic sterile insect technique approaches. BMC Genet. 15, S17 (2014).
Handler, A. M. Enhancing the stability and ecological safety of mass‐reared transgenic strains for field release by redundant conditional lethality systems. Insect Sci. 23, 225–234 (2016).
Akbari, O. S. et al. Safeguarding gene drive experiments in the laboratory. Science 349, 927–929 (2015).
Noble, C., Adlam, B., Church, G. M., Esvelt, K. M. & Nowak, M. A. Current CRISPR gene drive systems are likely to be highly invasive in wild populations. Elife 7, e33423 (2018).
Dhole, S., Vella, M. R., Lloyd, A. L. & Gould, F. Invasion and migration of spatially self-limiting gene drives: a comparative analysis. Evol. Appl. 11, 794–808 (2018).
Noble, C., Min, J. & Olejarz, J. Daisy-chain gene drives for the alteration of local populations. Proc. Natl Acad. Sci. USA 116, 8275–8282 (2019).
Dhole, S., Lloyd, A. L. & Gould, F. Tethered homing gene drives: a new design for spatially restricted population replacement and suppression. Evol. Appl. 12, 1688–1702 (2019).
Li, M. et al. Development of a confinable gene drive system in the human disease vector Aedes aegypti. Elife 9, e51701 (2020).
Basgall, E. M. et al. Gene drive inhibition by the anti-CRISPR proteins AcrIIA2 and AcrIIA4 in Saccharomyces cerevisiae. Microbiology 164, 464–474 (2018).
Handler, A. M. & Harrell, R. A. II Germline transformation of Drosophila melanogaster with the piggyBac transposon vector. Insect Mol. Biol. 8, 449–457 (1999).
Schetelig, M. F. & Handler, A. M. A functional comparison of the 3xP3 promoter by recombinase-mediated cassette exchange in Drosophila and a tephritid fly, Anastrepha suspensa. G3 (Bethesda) 3, 687–693 (2013).
Liu, Y. & Whittier, R. F. Thermal asymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking. Genomics 25, 674–681 (1995).
Acknowledgements
We thank Dr. Carsten Horn and Dr. Ernst Wimmer for sharing the M5.II lethal effector strain and the s1-tTA driver construct plasmid, and Dr. Rodney Nagoshi and Dr. Daniel Hahn for comments on the manuscript. This project was supported by the USDA-Biotechnology Risk Assessment Program competitive grant no. 2015-33522-24094 from the USDA National Institute of Food and Agriculture (to A.M.H.) and the Emmy Noether Program SCHE 1833-1/1 of the German Research Foundation (to M.F.S.). This study benefited from discussions at meetings for the Coordinated Research Project, “Comparing Rearing Efficiency and Competitiveness of Sterile Male Strains Produced by Genetic, Transgenic or Symbiont-based Technologies,” funded by the International Atomic Energy Agency (IAEA).
Author information
Authors and Affiliations
Contributions
Y.Z. and A.M.H. designed experiments and analyzed data; Y.Z. and M.F.S. performed research; and all authors wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Communications thanks Bruce Hay and the other anonymous reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhao, Y., Schetelig, M.F. & Handler, A.M. Genetic breakdown of a Tet-off conditional lethality system for insect population control. Nat Commun 11, 3095 (2020). https://doi.org/10.1038/s41467-020-16807-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-020-16807-3
This article is cited by
-
Sequence and expression analysis of the spermatogenesis-specific gene cognates, wampa and Prosα6T, in Drosophila suzukii
Genetica (2023)
-
Differences in barriers for controlled learning about safety between biotechnology and chemistry
Nature Communications (2022)
-
A transgenic female killing system for the genetic control of Drosophila suzukii
Scientific Reports (2021)
-
Identification and characterization of four Drosophila suzukii cellularization genes and their promoters
BMC Genetics (2020)
-
Functional characterization of the Drosophila suzukii pro-apoptotic genes reaper, head involution defective and grim
Apoptosis (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
