
Abstract
The evolutionary transition from self-incompatible distyly to self-compatible homostyly frequently occurs in heterostylous taxa. Although the inheritance of distyly and homostyly has been deeply studied, our understanding on modifications of the classical simple Mendelian model is still lacking. Primula forbesii, a biennial herb native to southwest China, is a typical distylous species, but after about 20 years of cultivation with open pollination, self-compatible homostyly appeared, providing ideal material for the study of the inheritance of distyly and homostyly. In this study, exogenous homobrassinolide was used to break the heteromorphic incompatibility of P. forbesii. Furthermore, we performed artificial pollination and open-pollination experiments to observe the distribution of floral morphs in progeny produced by different crosses. The viability of seeds from self-pollination was always the lowest among all crosses, and the homozygous S-morph plants (S/S) occurred in artificial pollination experiments but may experience viability selection. The distyly of P. forbesii is governed by a single S-locus, with S-morph dominant hemizygotes (S/−) and L-morph recessive homozygotes (−/−). Homostylous plants have a genotype similar to L-morph plants, and homostyly may be caused by one or more unlinked modifier genes outside the S-locus. Open pollinations confirm that autonomous self-pollination occurs frequently in L-morphs and homostylous plants. This study deepens the understanding of the inheritance of distyly and details a case of homostyly that likely originated from one or more modifier genes.
Similar content being viewed by others

Introduction
Transitions from obligate outcrossing to autonomous self-fertilization frequently occur in angiosperm species (Igic et al. 2006; Shao et al. 2019). However, in environments lacking pollinators and/or pollen donors, adaptive transitions toward selfing are favored and can be fixed in populations because of its reproductive advantage (Darwin 1876; Stebbins 1974; Busch and Delph 2012; Yuan et al. 2017). Changes in mating strategy can lead to reproductive isolation and thus are related to speciation at the population level (Wright et al. 2013). In plants, transitions from outcrossing to selfing are commonly accompanied by heritable directional shifts in floral morphology (e.g. reduced herkogamy and flower diameter) (Morgan and Barrett 1989; Sicard and Lenhard 2011; de Vos et al. 2014; Wu et al. 2017) and physiological traits (e.g. loss of self-incompatibility) (Mable et al. 2005; Igić et al. 2008; Zhong et al. 2019; Liu et al. 2022). Since Darwin’s seminal work, determining the inheritance of traits that structure the variation and evolution of mating systems has been a focus of evolutionary biologists, ecologists and geneticists (Darwin 1876; Shore and Barrett 1985; Barrett et al. 2014; Igic and Busch 2013). After more than a century of study, it has been revealed that the inheritance basis of variation in mating systems includes both some key genes with large effects and some quantitative genes with small effects (Fishman et al. 2002; Matsui et al. 2004; Slotte et al. 2012; Yuan et al. 2019). In the present study, based on crosses of heterostylous plants and novel homostylous plants, which present a paradigmatic model system for studying transitions form outcrossing to selfing, the inheritance of traits related to mating system variation have been explored, providing an empirical basis for further genetic and ecological research of mating system evolution.
Heterostyly is a heritable floral polymorphism that includes two (distyly) or three (tristyly) floral morphs characterized by reciprocal positioning of stigmas and anthers (Darwin 1877; Barrett 1992; Lloyd and Webb 1992). Heterostyly has independently originated in at least 28 angiosperm families and 199 genera, particularly in tubular-flowered species with animal-mediated pollination (Barrett and Shore 2008). The major expression of heterostyly is distyly, in which the flowers of one morph have a high stigma and low anthers (long style morph, L-morph, L), while the other morph expresses the opposite pattern (short style morph, S-morph, S) (Darwin 1877; Barrett 2019). The vast majority of distylous species possess heteromorphic incompatibility systems, in which only inter-morph pollination is compatible, while both intra-morph pollination and self-pollination are incompatible (Barrett and Shore 2008; Brys and Jacquemyn 2015). Distyly has long been regarded as an ideal mechanism to promote outcrossing and improve pollen dispersal efficiency since Darwin’s seminal work (Darwin 1877; Yuan et al. 2017). However, the discovery of numerous self-compatible homostylous plants (H-morph, H) with stigmas and anthers at the same position in distylous populations suggests a new potential evolutionary response to this floral polymorphism (Darwin 1877; Zhou et al. 2017; Barrett 2019; Zhong et al. 2019). Shifts to homostyly frequently occur in alpine environments, in peripheral populations and during colonization, when pollinators and/or mates are not reliably available (Brys and Jacquemyn 2015; Yuan et al. 2017; Shao et al. 2019; Zhang et al. 2021; Liu et al. 2022). Therefore, under conditions averse to outcrossing, distylous populations are forced by selection pressure to abandon obligate outcrossing and transition to self-pollination (Busch and Delph 2012). Although such transitions to selfing have ensured the survival and reproduction of species, they are frequently accompanied by inbreeding depression. Self-fertilization leads to higher homozygosity and the expression of deleterious recessive alleles, which eventually leads to reduced seed mass, germination rates, seedling vigor and reproductive capacity (Luijten et al. 2002; Charlesworth and Willis 2009; Barmentlo et al. 2018).
In the past century, many investigations of the inheritance of heterostyly and homostyly have focused on Primula (Bateson and Gregory 1905; Kappel et al. 2017). The genus Primula is comprised of ~430 species, with the vast majority (95%) being distylous and 45 being florally monomorphic (Richards 2003; Mast et al. 2006). Phylogenetic analysis of Primula clearly shows a single independent origin of distyly and multiple transitions to homostyly (Mast et al. 2006; de Vos et al. 2014; Zhong et al. 2019). The early genetic studies in Primula were mainly based on the findings of Bateson and Gregory (1905), who established that distyly was controlled by a single S-locus following Mendelian inheritance, with S-morph dominant heterozygotes (S/s) and L-morph recessive homozygotes (s/s). Then, based on numerous hybridization results of L-morph, S-morph and homostylous plants, Ernst (1955) constructed the S-locus supergene model comprised of at least three tightly linked genes, controlling style elongation, anthers position and pollen morphology, respectively. Therefore, researchers presumed that the homostyly frequently occurring in distylous population may have originated via homologous recombination (Pamela and Dowrick 1956; Lewis and Jones 1992; Barrett and Shore 2008). However, recent studies have demonstrated that the S-locus is actually a hemizygous region only existing in the S-morph (S/−) and absent in the L-morph (−/−) (Li et al. 2016; Kappel et al. 2017; Shore et al. 2019; Mora-Carrera et al. 2023). Further studies showed that the S-locus of Primula contains at least five genes, with CYP controlling style length and female self-incompatibility and GLO controlling anther height (Huu et al. 2016; Huu et al. 2020). Based on the hemizygous genetic model, homostyly in this system cannot have originated via homologous recombination within the S-locus, because there are no homologous sequences on the s haplotype with which it could pair and crossover (Barrett 2019). Consistent with Ernst (1955), the observed homostyly probably arose from mutations of S-locus coding sequence, promoter region variation and/ or gene structure rearrangements (Huu et al. 2016; Huu et al. 2020; Mora-Carrera et al. 2023). Through a less common pathway, homostyly may also arise from unlinked modifier genes outside the S-locus (Richards 1997; Sicard et al. 2011; Arunkumar et al. 2017; Barrett 2019). Under this evolutionary pathway, homostylous plants would exhibit considerable variation in the stigma and anther positions, which is more commonly reported in self-compatible species, rather than ones with strong self-incompatibility (Matsui et al. 2004; Brys and Jacquemyn 2015; Zhang et al. 2021).
Owing to the existence of heteromorphic incompatibility, the vast majority of previous studies on the inheritance of distyly and homostyly have been conducted on a few species that exhibit self-compatibility (Ernst 1955; Pamela and Dowrick 1956; Yuan et al. 2019). Some of the most recent research indicates style length in distyly is mainly developmentally determined by brassinosteroids, with the L-morph and S-morph containing high and low concentrations of brassinosteroids, respectively (Huu et al. 2016). Furthermore, brassinosteroids also play an important role in female self-incompatibility (Huu et al. 2022). The treatment of S-morph flower buds with exogenous brassinosteroids can induce ‘long homostyly,’ characterized by long styles and high anthers, rendering such flowers completely self-compatible (Huu et al. 2022). In the present study, through exogenous brassinosteroid treatment, we provide more comprehensive cross evidence elucidating the inheritance of distyly with strong heteromorphic self-incompatibility.
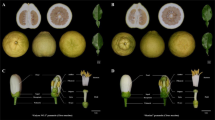
Primula forbesii Franch. (Primulaceae), a typically distylous species with L-morph that is partially self-compatible and S-morph that is completely self-incompatible (Huu et al. 2022; Liu et al. 2022), is a popular horticultural flower because of its pleasant fragrance and abundant purple flowers (Fig. 1A–C). However, preliminary observations revealed that after about 20 years of cultivation with open pollination in a greenhouse, a distylous population of P. forbesii eventually came to exhibit a broader range of floral morphs, producing homostylous plants with stigma and anthers in the same position (Fig. 1D). These variants provide an ideal material for the study of the inheritance of distyly and homostyly in P. forbesii. In order to harvest seeds produced by intra-morph pollination and self-pollination of S-morph plants, exogenous homobrassinolide (homo-BL) was injected into flower buds to render them self-compatible (Huu et al. 2022). In this study, we mainly focused on answering the following three questions. (1) What are the inheritance laws of distyly and homostyly? We aimed to determine whether homostyly had arisen by mutation of key genes at the S-locus or by one or more unlinked modifier genes outside the S-locus. (2) For partially absent heteromorphic incompatibility, does the disassortative mating system mediated by pollinators break down in P. forbesii? We predicted that autonomous self-fertilization would occur frequently in both L-morph and H-morph plants, with the latter being more obvious because of the proximity of the stigma to the anthers. (3) Is inbreeding depression strong in P. forbesii and at what stage is it manifested? Because self-fertilization may lead to the expression of deleterious recessive genes, we predict that the seeds resulting from self-pollination may have lower viability. In addition, owing to the intrinsic hemizygosity of distyly in this system, the S-locus may be linked with recessive lethal genes, such that homozygous S-morph plants (SS) are likely to die or experience severe viability selection. This prediction originates with the work of Richards (1997) and has found only limited support elsewhere (Yuan et al. 2019). In the present work, we tested the hypothesis and found support for this prediction by performing intra-morph pollination and self-pollination of S-morph plants and observing the floral distribution of progeny.

A Flowering plants. B–D Dissected L-morph, S-morph and H-morph flowers, showing differences in style length (blue arrowheads) and anther position (yellow arrowheads) (scale bar = 3 mm).
Material and methods
Primula forbesii is a subalpine biennial herb native to Yunnan, China, belonging to the section Monocarpicae of Primula (Chen and Hu 1990). In spring 2000, we collected flowering plants of P. forbesii from two wild distylous populations at Heilongtan (102°45′14″E, 25°8′38″N) and Xishan (102°37′34″E, 24°58′24″N) in Yunnan, China. All plants were then cultivated in the greenhouse at Sichuan Agricultural University, Chengdu Campus (103°51′44″E, 30°42′17″N). During greenhouse cultivation, no directional breeding was conducted, and the open-pollinated seeds were randomly harvested and sown every year. In spring 2018, we observed homostylous variants for the first time in the previously exclusively distylous population of P. forbesii.
For this study, seeds of P. forbesii were randomly harvested in April 2020 after natural open pollination and were sown into seedling trays in a greenhouse in July. One month later, more than 3000 of the resulting seedlings were transplanted into plastic pots (10 cm × 15 cm) filled with peat soil. Plants were watered every 3 days, and the light, temperature and humidity of the greenhouse were not artificially manipulated. At flowering, we randomly selected 300 plants to investigate the floral morph ratio of the population. The χ2-test implemented in SPSS 23.0 (IBM Corp., Armonk, NY, USA) was used to determine whether the L-morph and S-morph frequencies had deviated from the expected ratio of 1:1.
Brassinosteroid treatment
For the brassinosteroid treatment experiment, we selected at least 150 flowers of 30 L-morphs and 150 flowers of 30 S-morphs of P. forbesii. According to the methods of Huu et al. (2016), three experiment groups were established: (1) 10 µM homobrassinolide (homo-BL) (dissolved in dimethyl sulfoxide), (2) mock solution containing 0.1% dimethyl sulfoxide, consistent with the concentration of dimethyl sulfoxide in the 10 µM homo-BL solution, and (3) untreated. We injected flower buds (with petals of flower buds measuring 3 mm in length) every other day with the above solutions until they reached full flowering. After flowering, 50 flowers of each treatment were slit longitudinally and digitally photographed, and then, from those digital images, ImageJ was used to measure the style length, from the top of the ovary to the stigma, and the height of the anthers, from the top of the ovary to the anthers. Student’s t test was used to determine the significance of differences in the height of the stigma and anthers between flowers in the 10 µM homo-BL treatment and control groups in SPSS 23.0.
Controlled pollinations
To investigate the effects of exogenous brassinosteroids on heteromorphic incompatibility in P. forbesii, L-morph and S-morph flower buds were treated according to the above methods. Two experiment groups were established: 10 µM homo-BL application and mock application. After treatment, six artificial pollinations were conducted: inter-morph pollination (i.e., L × S and S × L) and both intra-morph pollination and self-pollination of L-morph and S-morph plants. For each pollination type, we collected approximately 20 plants and more than 150 flowers, and all flowers used were in their second or third day of blossoming. To exclude pollinators, flowers were bagged before blossoming and after artificial pollination, and for crossing treatments, all flowers were emasculated and bagged prior to blossoming. For each cross-pollination, pollen was collected from more than three flowering plants. All pollinations were performed in February 2021, and the fruits and seeds were gathered in April–May, at which time fruit set and seed production per fruit were recorded. Student’s t tests, as implemented in SPSS 23.0, were used to determine the effectiveness of exogenous homo-BL on breaking self-incompatibility of P. forbesii. Furthermore, to investigate the inheritance of distyly in P. forbesii, seeds of different artificial pollinations of L-morphs and S-morphs were gathered and stored at 4 °C after being fully dried in air.
To determine the origination and inheritance of naturally arising homostyly, different hand pollinations of H-morphs were conducted using the above methods, including self-pollination, cross-pollination with other H-morph plants, and crossing with L-morph (i.e., H × L and L × H) and S-morph plants (i.e., H × S and S × H) (Yuan et al. 2019). Finally, the resulting seeds were gathered and stored as described above.
Thousand-seed weight and germination rate
To assay inbreeding depression in P. forbesii, the weight of one thousand seeds and rate of seed germination were measured. To measure the thousand-seed weight of each cross, 500 seeds from each treatment were randomly selected and weighed on an electronic balance, with three replicates of each treatment. To investigate the germination rate (Jiang et al. 2018), 100 healthy and plump seeds of each pollination type were selected. After rinsing them with distilled water three times, seeds were neatly placed in a Petri dish with wet filter paper. The culture dish was then placed in a cabinet, maintained at 20 ± 2 °C, 60% humidity and an illumination level of 3500 lx for 12 h. During the experiment, water was replenished regularly to keep the filter paper wet continuously. Each germination was recorded only if the radicle length exceeded the diameter of the seed. After 1 week, the germination test was ended once no seeds had germinated for 5 consecutive days. There were three replicates of each treatment. We used one-way analysis of variance (ANOVA) to identify significant differences in thousand-seed weight and germination rate among the different pollination types.
Inheritance of distyly and homostyly
The progeny derived from artificial outcrossing and self-pollination were used to determine the inheritance laws of distyly and homostyly in P. forbesii. In the greenhouse, the seeds of each pollination type were germinated on soil-filled seed trays in July 2021. After a month, 100 seedlings of each pollination type were transplanted into plastic pots (10 cm × 15 cm) filled with peat soil, and the plants were maintained using the above-described methods. At flowering, the floral morph ratio of F1 progeny of each pollination type was recorded. We used χ2-tests, as implemented in SPSS 23.0, to determine whether the ratio of L-morph to S-morph progeny had deviated from the expected ratios of 1:1, 1:2 or 1:3. Furthermore, more than 20 flowers were randomly selected from progeny of each pollination. Flowers were slit longitudinally and digitally photographed, and the height of the stigmata and anthers were measured from the top of the ovary using ImageJ.
Investigation of progeny from open pollination
To assess the mating style of different floral morph of P. forbesii under open pollination, 10 L-morphs, 10 S-morphs and 10 H-morphs were selected, and all of them were then naturally pollinated by insects in the greenhouse from February to May 2021. To make the conditions as realistic as possible, the 30 selected plants were not isolated from other plants. The seeds of each floral morph after open-pollination were harvested and stored as described above. We grew these seeds to mature flowering plants using the above methods and recorded the floral morph ratio of progeny of each maternal seed family. We used the χ2-test implemented in SPSS 23.0 to determine whether the frequencies of L-morph and S-morph plants had deviated from the expected ratio of 1:1.
Results
Effects of exogenous brassinosteroid on floral morphology
Treatment with exogenous homo-BL significantly increased the height of the stigma and anthers of P. forbesii (Fig. 2 and Fig. 3). Compared with plants in the control group (3.32 ± 0.03 mm) and the mock treatment group (3.26 ± 0.05 mm), 10 µM homo-BL treatments (4.09 ± 0.07 mm) significantly increased the style length of the S-morph (10 µM homo-BL versus CK, t = −9.80, d.f. = 98, P < 0.001; 10 µM homo-BL versus mock, t = −9.98, d.f. = 98, P < 0.001), and some individuals developed ‘long homostyly’ (Fig. 2B). However, the style length of L-morph plants did not show a similar response, suggesting that exogenous homo-BL especially promotes the growth of S-morph plants. Under exogenous homo-BL treatment, there were different degrees of increase in anther height in both L-morph and S-morph flowers.

A L-morph flowers. B S-morph flowers.

A L-morph flowers. B S-morph flowers. Differences were not significant (ns), significant (*P < 0.05) orvery significant (**P < 0.01; ***P < 0.001) based on Student’s t test. Box edges indicate the lower and upper quartiles, respectively, the white middle line represents the median and the black dot and squares represent the mean value and abnormal values, respectively. The vertical bars indicate upper and lower limits of the data or extend to 1.5 times beyond the upper or lower quartile.
Effects of exogenous homobrassinolide on heteromorphic self-incompatibility
The L-morph was partially self-compatible, with about 60% fruit set and 100 seeds per fruit under intra-morph pollination and self-pollination, and exogenous homo-BL treatments had little effect on the self-incompatibility of L-morphs (Fig. 4A, B). On the contrary, S-morph plants were completely self-incompatible, and 10 µM homo-BL treatments induced a significant break down of the self-incompatibility system (Fig. 4C, D). For inter-morph pollinations, the homo-BL treatments significantly decreased the fruit set (0.77 ± 0.03) and seed production (172.80 ± 9.30) compared with plants subjected to mock treatments (fruit set 0.92 ± 0.19, t = 4.07, d.f. = 38, P < 0.001; seed production 199.80 ± 7.28, t = 2.29, d.f. = 18, P = 0.04). In contrast, unlike the mock treatment pollinations, in which almost no seeds were produced under intra-morph pollination and self-pollination, the S-morph plants treated with 10 µM homo-BL had become fully self-compatible, with high fruit set (intra-morph pollination 0.80 ± 0.03; self-pollination 0.82 ± 0.03) and seed production (intra-morph pollination, 204.40 ± 6.95; self-pollination, 211.70 ± 8.60).

A-B L-morph plants. C-D S-morph plants. Differences were not significant (ns), significant (*P < 0.05) or very significant (**P < 0.01; ***P < 0.001) based on Student’s t test.
Thousand-seed weight and germination rate
As shown in Table 1, the seed mass and vigor of the three morphs of P. forbesii were decreased to different degrees when seeds were obtained by self-pollination. For maternal L-morphs, the thousand-seed weight of intra-morph pollinations and self-pollinations were significantly reduced compared with L × S crosses (F3,8 = 4.77, P = 0.03), while the germination rate did not differ significantly among the different pollinations (F3,8 = 1.45, P = 0.30). In maternal S-morphs, there were no significant differences in the thousand-seed weight (F3,8 = 3.20, P = 0.08), while germination rates of seeds from intra-morph pollinations and self-pollinations, at only 62.67 ± 6.36 and 52.67 ± 1.76%, respectively, were significantly lower than those of S × L and S × H seeds (F3,8 = 14.34, P = 0.01). For maternal H-morphs, self-pollination was associated with the lowest thousand-seed weight (87.33 ± 3.19 mg) and germination rate (50.00 ± 7.57%) among all 12 pollination types.
Inheritance of distyly and homostyly
Inheritance of distyly in P. forbesii was consistent with a standard Mendelian inheritance model (Table 2 and Fig. 5). The progeny of L-morph intra-morph pollinations and self-pollinations were all L-morphs. The floral morph frequency of progeny of S-morph intra-morph pollinations and self-pollinations did not deviate significantly from an L:S ratio of 1:3 (S × S, χ2 = 0.42, d.f. = 1, P = 0.56; S-selfing, χ2 = 0.13, d.f. = 1, P = 0.81), although this frequency also did not deviate significantly from 1:2 (S × S, χ2 = 1.32, d.f. = 1, P = 0.28; S-selfing, χ2 = 1.80, d.f. = 1, P = 0.22). Meanwhile, inter-morph pollinations yielded an L:S ratio of progeny that did not deviate from 1:1 (S × L, χ2 = 0.04, d.f. = 1, P = 0.92; L × S, χ2 = 0.51, d.f. = 1, P = 0.54). Moreover, for the progeny derived from outcrossing and selfing, the measurement of flower sex organs confirmed that there was an obvious bimodal pattern in stigma and anther height (Fig. 5).

A S-morph × L-morph; B L-morph × S-morph; C S-morph × H-morph; D H-morph × S-morph; E L-morph × H-morph; F H-morph × L-morph; G S-morph × S-morph outcross; H L-morph × L-morph outcross; I H-morph × H-morph outcross; J S-morph selfing; K L-morph selfing; L H-morph selfing.
For seeds from selfing and outcrossing with other homostylous plants, almost all progeny were also homostylous plants. The progeny derived from crossing the H-morph and S-morph exhibited a roughly 1:1 floral ratio of L-morph and S-morph plants (S × H, χ2 = 0.65, d.f. = 1, P = 0.48; H × S, χ2 = 0.65, d.f. = 1, P = 0.48), while the progeny of crosses between the H-morph and L-morph were mainly comprised of L-morphs and a few H-morphs. Furthermore, for the progeny resulting from selfing and outcrossing of H-morphs, measurements of floral sex-organ position showed that there was substantial variation in stigma and anther height (e.g., Fig. 5I, L).
Progeny of open pollination
As shown in Table 3, the floral morph ratio of the cultivated population of P. forbesii was an L:S:H ratio of 0.52:0.40:0.08, in which the floral frequency of L-morphs and S-morphs significantly deviating from the expected L:S ratio of 1:1 (χ2 = 4.94, d.f. = 1, P = 0.03). For the L-morph seed families from open pollination, the progeny included L-morph, S-morph and H-morph plants, with an L:S ratio of 0.6:0.34, significantly deviating from 1:1 (χ2 = 7.19, d.f. = 1, P = 0.01). In the S-morph seed families, the progeny only included L-morph and S-morph plants, with an L:S ratio of 0.52:0.48, and the deviation from 1:1 was not significant (χ2 = 0.16, d.f. = 1, P = 0.76). In the H-morph seed families, many homostylous plants were observed, and the L:S:H floral morph ratio was 0.31:0.38:0.30.
Discussion
The present study mainly focused on the inheritance of distyly and homostyly of P. forbesii and obtained the following key findings. First, confirming the results of previous research (Huu et al. 2022), the style length and heteromorphic incompatibility of the S-morph of P. forbesii could be changed by exogenous homo-BL, and thus, intra-morph pollination and self-pollination progeny of S-morphs can be obtained. Second, owing to inbreeding depression, the thousand-seed weight and germination rate of each floral morph decreased to different degrees after self-pollination. In addition, based on a 1:3–1:2 ratio of L-morph and S-morph progeny of intra-morph pollinations and self-pollinations of S-morphs, mature homozygous S-morphs (S/S) may experience viability selection. Third, based on the hemizygous inheritance model of Primula, distyly of P. forbesii is controlled by a single S-locus, in which the L-morph is homozygous recessive (−/−) and the S-morph is hemizygous dominant (S/−). Consistent with Matsui et al. (2004), the homostyly may possess a genotype similar to that of the L-morph and arise via one or more unlinked modifier genes outside the S-locus. Finally, we determined that the autonomous self-pollination of L-morph and H-morph plants occurred frequently in the cultivated population, leading to the deviation from the equilibrium isocline of L-morphs and S-morphs. These key results of this study are discussed in detail below.
Exogenous homobrassinolide changes style length and female self-incompatibility
Self-incompatibility has been deeply studied in homomorphic plants, including sporophytic and gametophytic self-incompatibility (McCubbin 2008). In the former system, the pollen barrier site occurs on the stigmatic surface, while in the latter, it mainly occurs within the style (Allen and Hiscock 2008; Lu et al. 2018). For a heteromorphic incompatibility system, the incompatible pollen tube on the L-morph styles can partially enter the ovary but cannot germinate on the S-morph stigma (Huang et al. 2015; Jiang and Li 2017; Liu et al. 2022). For a long time, the female incompatibility of S-morphs had been considered entirely sporophytic, while such incompatibility was considered gametophytic in L-morphs (Lewis and Jones 1992; McCubbin 2008). Recently, Huu et al. (2022) provided a comprehensive and deep discussion on heteromorphic incompatibility in P. forbesii. They found that the gene CYP734A50, which encodes a brassinosteroid-inactivating enzyme, determined both style elongation and female self-incompatibility of S-morph plants. In the present study, consistent with previous results, by injecting exogenous homo-BL into S-morph flower buds, we successfully obtained ‘long homostyly’ and overcame self-incompatibility. Thus, the progeny of intra-morph pollination and self-pollination of S-morphs were successfully obtained.
Inbreeding depression and sheltered load
Under reproductive pressure caused by unreliable pollinator service and/or a limited number of mates, many strictly outcrossing species evolve self-compatibility (Stebbins 1974; Busch and Delph 2012; Igic and Busch 2013). Although the evolutionary transitions ensure the normal survival and reproduction of the population, it is commonly accompanied by strong inbreeding depression (Lande and Schemske 1985). Frequent self-fertilization in a population will result in increased homozygosity and the expression of recessive deleterious alleles, ultimately decreasing the fitness of progeny (Luijten et al. 2002; Charlesworth and Willis 2009; Barmentlo et al. 2018). Consistent with previous studies, we observed that the three floral morphs of P. forbesii exhibited declines in fitness after self-pollination, with seed masses and germination rates significantly lower than those of inter-morph pollination.
Owing to the forced hemizygosity of the S-locus in distylous species, it is postulated that the S-locus was tightly linked to one or more recessive deleterious alleles, a putative example of a phenomenon termed ‘sheltered load’ (Richards 1997; Strobeck 1972). According to this hypothesis, homozygous S-morphs (S/S) would die or exhibit reduced viability in natural conditions. However, the empirical evidence for a ‘sheltered load’ associated with a S-locus are mixed (Mather and De Winton 1941; Barrett et al. 1989; Kurian and Richards 1997). In the present study, the L-morph to S-morph ratio was distributed between 1:3 and 1:2 in progeny of intra-morph pollinations and self-pollinations of S-morph plants. Consistent with Yuan et al. (2019), the mature homozygous S-morph plants may exist in progeny but also probably experience strong viability selection owing to linked loads.
Inheritance of distyly and homostyly
Heterostyly presents a classic model for the study of the shift from outcrossing to selfing. Hitherto, the inheritance of heterostyly has been investigated in c. 11 families, with the vast majority of studies focusing on distylous taxa (Lewis and Jones 1992). Distyly is controlled by a single S-locus linkage group, with two exceptions in which the dominance relations are reversed, with S-morph dominant heterozygotes (S/s) and L-morph recessive homozygotes (s/s) (Charlesworth and Charlesworth 2009; Barrett 2019). Based on molecular data, the S-haplotype of the S-morph actually contains a hemizygous region consisting of at least five genes, which is absent from the s-haplotype of L-morphs (Li et al. 2016; Kappel et al. 2017). The hemizygous characteristics of the S-locus have been demonstrated in Primula, Turnera, Fagopyrum, and Linum (Yasui et al. 2012; Li et al. 2016; Shore et al. 2019; Gutiérrez-Valencia et al. 2022). In the present study, the results of different cross experiments are consistent with previous inheritance models, indicating that the S-morph is dominant hemizygous (S/−) and the L-morph is recessive homozygote (−/−) in P. forbesii. It is unsurprising that numerous Primula species share the same inheritance pattern, because phylogenetic analysis of Primula has demonstrated a single origination of distyly (Mast et al. 2006).
Self-compatible homostyly occurs frequently in distylous populations under reproductive pressure. Homostylous flowers commonly combine the sex organs of distyly, in which long homostyly occurs as a chimera of the styles of L-morphs and anthers of S-morphs, and short homostyly exhibits the opposite combination (Barrett and Shore 2008). Therefore, homostyly for a long time had been considered to originate from homologous recombination within the S-locus region (Pamela and Dowrick 1956; Lewis and Jones 1992). However, this hypothesis has now been rejected after the dominant S-haplotype was demonstrated to be hemizygous, because the s-haplotype could not provide homologous sequences with which to pair and cross-over during meiosis (Li et al. 2016; Kappel et al. 2017). Homostyly probably has arose via mutations of the key genes controlling either style length or anther height within the S-locus, which has been supported by molecular data from many distylous species (Huu et al. 2016; Shore et al. 2019; Huu et al. 2020). However, Mora-Carrera et al. (2023) found in P. vulgaris that the S-morph and some long-homostylous plants share exactly the same CYP allele, which could inhibit style elongation by inactivating brassinosteroid and thus produce short styles. These results suggested that in addition to loss-of-function mutation of the coding region of specific genes, homostyly may also arise via other substitution mechanisms, such as promoter region mutations, structural rearrangements and mutations to unlinked modifier genes outside the S-locus (Mather and De Winton 1941; Ganders 1979; Kappel et al. 2017). In the present study, the progeny of intra-morph pollination and self-pollination of homostylous plants were almost all homostylous, indicating that the homostylous variants of P. forbesii were homozygous. Consistent with the findings of Matsui et al. (2004), no or only a few homostylous plants were observed to result from cross experiments of homostylous plants with S-morphs and L-morphs, indicating that homostyly could not originate from the mutations of key genes within the S-locus. Based on crossing data alone, we speculate that homostylous plants may have a genotype similar to that of L-morphs (−/−) and have arisen via one or more recessive modifier genes outside the S-locus. There were a few homostylous plants among the progeny of crosses between homostylous plants and L-morphs, which may indicate that some heterozygous L-morphs provide recessive modifier genes. However, it is still difficult to explain why there were a small number of L-morphs among the progeny of intra-morph pollinations and self-pollinations of homostylous plants. Environmental factors may play a role in the expression of polygenic modifiers based on the considerable fluctuation of sex-organ height in progeny. Thus, it will be necessary to sequence the genes that control the style length and anthers height (e.g., CYP734A50 and GLO2), which will provide direct evidences for the origination and inheritance of homostyly in P. forbesii.
Mating patterns in open-pollinated conditions
Heteromorphic incompatibility has great significance in maintaining the disassortative mating and equilibrium isocline of floral morphs in heterostylous species (Ganders 1979; Brys and Jacquemyn 2015; Yuan et al. 2017; Yuan et al. 2019). For distylous species partially or completely lacking heteromorphic incompatibility, the distribution of progeny for each floral morph after open pollination is an effective method to evaluate mating style, although more quantitative evaluation requires the use of molecular markers (Barrett and Shore 1987; Yuan et al. 2019). In previous studies, the absence of heteromorphic incompatibility was shown to seriously disrupt the disassortative mating system and lead to a deviation from a 1:1 ratio of L-morphs and S-morphs (Barrett 2003; Yuan et al. 2017; Shao et al. 2019; Mora-Carrera et al. 2023). In the cultivated population of P. forbesii, the open-pollinated progeny of S-morphs did not different from a 1:1 morph ratio. However, open-pollinated L-morphs apparently produced more progeny of L-morph, about twice the number of S-morphs. This pattern is likely caused by frequent occurrences of intra-morph fertilization and self-fertilization in L-morph plants owing to the weakened heteromorphic incompatibility. Owing to the loss of heteromorphic incompatibility and herkogamy, homostylous plants would have a higher incidence of autonomous self-pollination compared with distylous plants. In the present study, nearly 1/3 of the open-pollinated progeny of homostylous plants were homostylous. The open-pollination results accord with the inheritance pattern described above, as self-pollination of homostylous plants would produce exclusively homostylous plants, indicating that autonomous self-pollination occurs frequently in homostylous plants. Furthermore, the biased morph ratio of the cultivated population of P. forbesii corresponded well with above results, indicating the frequent occurrence of self-pollination in L-morph and homostylous plants in the open pollination conditions. Therefore, theory can predict, although the direct evidence remains lacking (Curtis and Curtis 1985), that S-morphs may be further reduced and ultimately result in a homostylous population with a few L-morph plants (Crosby 1949; Crosby 1960; Mora-Carrera et al. 2023). Studies on large time scales are necessary, which will provide direct evidence to elucidate the effects of shifts in mating systems on floral morph composition and evolutionary fate of heteromorphic populations.
Data availability
All data needed to evaluate the conclusions in the paper are present in the paper.
References
Allen AM, Hiscock SJ (2008) Evolution and Phylogeny of Self-Incompatibility Systems in Angiosperms. In: Franklin-Tong VE (ed) Self-Incompatibility in Flowering Plants: Evolution, Diversity, and Mechanisms. Springer, Berlin, p 73–101
Arunkumar R, Wang W, Wright SI, Barrett SC (2017) The genetic architecture of tristyly and its breakdown to self‐fertilization. Mol Ecol 3:752–765
Barmentlo SH, Meirmans PG, Luijten SH, Triest L, Oostermeijer JGB (2018) Outbreeding depression and breeding system evolution in small, remnant populations of Primula vulgaris: consequences for genetic rescue. Conserv Genet 19:545–554
Barrett SCH, Morgan MT, Husband BC (1989) The dissolution of a complex genetic polymorphism: the evolution of self-fertilization in tristylous Eichhornia paniculata (Pontederiaceae). Evolution 43:1398–1416
Barrett SCH (1992) Evolution and function of heterostyly. Springer-Verlag, Berlin
Barrett SCH (2003) Mating strategies in flowering plants: the outcrossing–selfing paradigm and beyond. Philos Trans R Soc Lond, Ser B: Biol Sci 358:991–1004
Barrett SCH (2019) ‘A most complex marriage arrangement’: recent advances on heterostyly and unresolved questions. N. Phytol 224:1051–1067
Barrett SCH, Arunkumar R, Wright SI (2014) The demography and population genomics of evolutionary transitions to self-fertilization in plants. Philos Trans R Soc Lond, Ser B: Biol Sci 369:20130344
Barrett SCH, Shore JS (1987) Variation and evolution of breeding systems in the Turnera ulmifolia L. complex (Turneraceae). Evolution 41:340–354
Barrett SCH, Shore JS (2008) New Insights on Heterostyly: Comparative Biology, Ecology and Genetics. In: Franklin-Tong VE (ed) Self-Incompatibility in Flowering Plants: Evolution, Diversity, and Mechanisms. Springer, Berlin, p 3–32
Bateson W, Gregory RP (1905) On the inheritance of heterostylism in Primula. Proc R Soc B-Biol Sci 76:581–586
Brys R, Jacquemyn H (2015) Disruption of the distylous syndrome in Primula veris. Ann Bot 115:27–39
Busch JW, Delph LF (2012) The relative importance of reproductive assurance and automatic selection as hypotheses for the evolution of self-fertilization. Ann Bot 109:553–562
Charlesworth B, Charlesworth D (2009) Darwin and genetics. Genetics 183:757–766
Charlesworth D, Willis JH (2009) The genetics of inbreeding depression. Nat Rev Genet 10:783–796
Chen F, Hu Q (1990) Flora of China. Science Press, Beijing
Crosby JL (1949) Selection of an unfavourable gene-complex. Evolution 3:212–230
Crosby JL (1960) The use of electronic computation in the study of random fluctuations in rapidly evolving populations. Philos Trans R Soc B-Biol Sci 242:551–573
Curtis J, Curtis CF (1985) Homostyle primroses re-visited. I. Variation in timeand space. Heredity 54:227–234
Darwin C (1876) The effects of cross and self fertilisation in the vegetable kingdom. John Murray, London
Darwin C (1877) The different forms of flowers on plants of the same species. John Murray, London
Ernst A (1955) Self-fertility in monomorphic Primulas. Genetica 27:391–448
Fishman L, Kelly AJ, Willis JH (2002) Minor quantitative trait loci underlie floral traits associated with mating system divergence in Mimulus. Evolution 56:2138–2155
Ganders FR (1979) Heterostyly in Lithospermum cobrense (Boraginaceae). Am J Bot 66:746–748
Gutiérrez-Valencia J, Fracassetti M, Berdan EL, Bunikis I, Soler L, Dainat J et al. (2022) Genomic analyses of the Linum distyly supergene reveal convergent evolution at the molecular level. Curr Biol 32:4360–4371. e6
Huang Y, Li NW, Ren ZX, Chen G, Wu ZK, Ma YP (2015) Reproductive biology of Primula beesiana (Primulaceae), an alpine species endemic to Southwest China. Plant Ecol Evol 148:289–296
Huu CN, Kappel C, Keller B, Sicard A, Takebayashi Y, Breuninger H et al. (2016) Presence versus absence of CYP734A50 underlies the style-length dimorphism in primroses. Elife 5:e17956
Huu CN, Keller B, Conti E, Kappel C, Lenhard M (2020) Supergene evolution via stepwise duplications and neofunctionalization of a floral-organ identity gene. Proc Natl Acad Sci USA 117:23148–23157
Huu CN, Plaschil S, Himmelbach A, Kappel C, Lenhard M (2022) Female self-incompatibility type in heterostylous Primula is determined by the brassinosteroid-inactivating cytochrome P450 CYP734A50. Curr Biol 32:671–676. e5
Igic B, Bohs L, Kohn JR (2006) Ancient polymorphism reveals unidirectional breeding system shifts. Proc Natl Acad Sci USA 103:1359–1363
Igić B, Lande R, Kohn JR (2008) Loss of self-incompatibility and its evolutionary consequences. Int J Plant Sci 169:93–104
Igic B, Busch JW (2013) Is self‐fertilization an evolutionary dead end? N Phytol 198:386–397
Jiang XF, Zhu XF, Chen LL, Li QJ (2018) What ecological factors favor the shift from distyly to homostyly? A study from the perspective of reproductive assurance. J Plant Ecol 11:645–655
Jiang XF, Li QJ (2017) Self- and intra-morph incompatibility and selection analysis of an inconspicuous distylous herb growing on the Tibetan plateau (Primula tibetica). Ecol Evol 7:5746–5753
Kappel C, Huu CN, Lenhard M (2017) A short story gets longer: recent insights into the molecular basis of heterostyly. J Exp Bot 68:5719–5730
Kurian V, Richards AJ (1997) A new recombinant in the heteromorphy ‘S’ supergene in Primula. Heredity 78:383–390
Lande R, Schemske DW (1985) The Evolution of Self-Fertilization and Inbreeding Depression in Plants. II. Empirical observations. Evolution 39:41–52
Lewis D, Jones D (1992) The genetics of heterostyly. In: Barrett SCH (ed) Evolution and function of heterostyly. Springer, Berlin, p 129–150
Li J, Cocker JM, Wright J, Webster MA, McMullan M, Dyer S et al. (2016) Genetic architecture and evolution of the S locus supergene in Primula vulgaris. Nat Plants 2:1–7
Liu CL, Jia Y, Li YF, Xiang YF, Pan YZ, Liu QL et al. (2022) The rapid appearance of homostyly in a cultivated distylous population of Primula forbesii. Ecol Evol 12:e9515
Lloyd D, Webb C (1992) The evolution of heterostyly. In: Barrett SCH (ed) Evolution and function of heterostyly. Springer, Berlin, p 151–178
Lu W, Bian X, Yang W, Cheng T, Wang J, Zhang Q et al. (2018) Transcriptomics investigation into the mechanisms of self-incompatibility between pin and thrum morphs of Primula maximowiczii. Int J Mol Sci 19:1840
Luijten SH, Kéry M, Oostermeijer JGB, Nijs HCD (2002) Demographic consequences of inbreeding and outbreeding in Arnica montana: a field experiment. J Ecol 90:593–603
Mable BK, Robertson AV, Dart S, Berardo CD, Witham L, Fenster C (2005) Breakdown of self-incompatibility in the perennial Arabidopsis lyrata (Brassicaceae) and its genetic consequences. Evolution 59:1437–1448
Mast AR, Kelso S, Conti E (2006) Are any primroses (Primula) primitively monomorphic? N. Phytol 171:605–616
Mather K, De Winton D (1941) Adaptation and counter-adaptation of the breeding system in Primula. Ann Bot 5:297–311
Matsui K, Nishio T, Tetsuka T (2004) Genes outside the S supergene suppress S functions in buckwheat (Fagopyrum esculentum). Ann Bot 94:805–809
McCubbin A (2008) Heteromorphic self-incompatibility in Primula: twenty-first century tools promise to unravel a classic nineteenth century model system. In: Franklin-Tong EV (ed) In Self-incompatibility in flowering plants. Springer, Berlin, p 289–308
Mora-Carrera E, Stubbs R, Keller B, Léveillé-Bourret T, Conti E (2023) Different molecular changes underlie the same phenotypic transition: origins and consequences of independent shifts to homostyly within species. Mol Ecol 32:61–78
Morgan MT, Barrett SCH (1989) Reproductive correlates of mating system evolution in Eichhornia paniculata (Spreng.) Solms (Pontederiaceae). J Evol Biol 2:183–203
Pamela V, Dowrick J (1956) Heterostyly and homostyly in Primula obconica. Heredity 10:219–236
Richards AJ (1997) Plant breeding systems. Chapman and Hall, London
Richards AJ (2003) Primula. Timber Press, Portland
Shao JW, Wang HF, Fang SP, Conti E, Chen YJ, Zhu HM (2019) Intraspecific variation of self-incompatibility in the distylous plant Primula merrilliana. AoB Plants 11:plz030
Shore JS, Barrett SC (1985) The genetics of distyly and homostyly in Turners ulmifolia L.(Turneraceae). Heredity 55:167–174
Shore JS, Hamam HJ, Chafe PD, Labonne JD, Henning PM, McCubbin AG (2019) The long and short of the S‐locus in Turnera (Passifloraceae). N Phytol 224:1316–1329
Sicard A, Lenhard M (2011) The selfing syndrome: a model for studying the genetic and evolutionary basis of morphological adaptation in plants. Ann Bot 107:1433–1443
Sicard A, Stacey N, Hermann K, Dessoly J, Neuffer B, Bäurle I et al. (2011) Genetics, evolution, and adaptive significance of the selfing syndrome in the genus Capsella. Plant Cell 23:3156–3171
Slotte T, Hazzouri KM, Stern D, Andolfatto P, Wright SI (2012) Genetic architecture and adaptive significance of the selfing syndrome in Capsella. Evolution 66:1360–1374
Stebbins GL (1974) Flowering plants: evolution above the species level. Belknap Press, Cambridge
Strobeck C (1972) Heterozygosity in pin-thrum plants or with partial sex linkage. Genetics 72:667–678
Wright SI, Kalisz S, Slotte T (2013) Evolutionary consequences of self-fertilization in plants. Proc R Soc Lond, Ser B: Biol Sci 280:20130133
de Vos JM, Wueest R, Conti E (2014) Small and ugly? Phylogenetic analyses of the ‘selfing syndrome’ reveal complex evolutionary fates of monomorphic primrose flowers. Evolution 68:1042–1057
Wu LY, Wang B, Schoen DJ, Huang SQ (2017) Transitions from distyly to homostyly are associated with floral evolution in the buckwheat genus (Fagopyrum). Am J Bot 104:1232–1240
Yasui Y, Mori M, Aii J, Abe T, Matsumoto D, Sato S et al. (2012) S-LOCUS EARLY FLOWERING 3 Is Exclusively Present in the Genomes of Short-Styled Buckwheat Plants that Exhibit Heteromorphic Self-Incompatibility. Plos ONE 7:e31264
Yuan S, Barrett SC, Duan T, Qian X, Shi M, Zhang D (2017) Ecological correlates and genetic consequences of evolutionary transitions from distyly to homostyly. Ann Bot 120:775–789
Yuan S, Barrett SC, Li C, Li X, Xie K, Zhang D (2019) Genetics of distyly and homostyly in a self-compatible Primula. Heredity 122:110–119
Zhang W, Hu YF, He X, Zhou W, Shao JW (2021) Evolution of Autonomous Selfing in Marginal Habitats: Spatiotemporal Variation in the Floral Traits of the Distylous Primula wannanensis. Front Plant Sci 12:781281
Zhong L, Barrett SC, Wang XJ, Wu ZK, Sun HY, Li DZ et al. (2019) Phylogenomic analysis reveals multiple evolutionary origins of selfing from outcrossing in a lineage of heterostylous plants. N Phytol 224:1290–1303
Zhou W, Barrett SC, Li HD, Wu ZK, Wang XJ, Wang H et al. (2017) Phylogeographic insights on the evolutionary breakdown of heterostyly. N Phytol 214:1368–1380
Acknowledgements
The authors thank Kehang Ma, Xilin Wang, Xing Lin and Minjie Jiang for their assistance during the pollination work. This work was supported by the National Natural Science Foundation of China (32001356).
Author information
Authors and Affiliations
Contributions
YJ, CL, and YP planned and designed the research. CL, YL, YX, and ZW performed the experiments. XY and QL analyzed the data. CL, YJ, and SG wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Jia, Y., Liu, C., Li, Y. et al. Inheritance of distyly and homostyly in self-incompatible Primula forbesii. Heredity 130, 259–268 (2023). https://doi.org/10.1038/s41437-023-00598-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41437-023-00598-6
