
Abstract
Plant survival requires an ability to adapt to differing concentrations of nutrient and toxic soil ions, yet ion sensors and associated signaling pathways are mostly unknown. Aluminum (Al) ions are highly phytotoxic, and cause severe crop yield loss and forest decline on acidic soils which represent ∼30% of land areas worldwide. Here we found an Arabidopsis mutant hypersensitive to Al. The gene encoding a leucine-rich-repeat receptor-like kinase, was named Al Resistance1 (ALR1). Al ions binding to ALR1 cytoplasmic domain recruits BAK1 co-receptor kinase and promotes ALR1-dependent phosphorylation of the NADPH oxidase RbohD, thereby enhancing reactive oxygen species (ROS) generation. ROS in turn oxidatively modify the RAE1 F-box protein to inhibit RAE1-dependent proteolysis of the central regulator STOP1, thus activating organic acid anion secretion to detoxify Al. These findings establish ALR1 as an Al ion receptor that confers resistance through an integrated Al-triggered signaling pathway, providing novel insights into ion-sensing mechanisms in living organisms, and enabling future molecular breeding of acid-soil-tolerant crops and trees, with huge potential for enhancing both global food security and forest restoration.
Similar content being viewed by others
Introduction
Aluminum (Al) is the most abundant metal in the Earth’s crust (∼8% by weight). However, the Al ion is highly toxic to plants. When soils become acidic, part of Al is solubilized from insoluble aluminosilicates or oxides to form soluble ions.1 The resultant phytotoxic Al ions can rapidly enter root cells, and cause a series of cellular damages,2 thus inhibiting root growth and function of most plants at very low micromolar concentrations.3 These effects substantially reduce crop yields, particularly when combined with other stresses, such as drought and nutrient deficiency. Al toxicity is therefore recognized as the major factor limiting agricultural productivity on acid soils which occupy ∼30% of territorial land area and up to 50% of the potential arable lands worldwide, and is exceeded only by drought among abiotic limitations to crop production.4,5 Moreover, Al toxicity is an important contributor to forest decline,6 posing a real threat to the global ecological environment.
Decades of research have established the central role of the secretion of organic acid anions (including malate, citrate and oxalate) in Al resistance in the main crops.3,7,8 These anions chelate and restrict Al ions from entering the root apex, the primary site of Al toxicity.2 Genetically enhancing their biosynthesis or extrusion significantly increases crop Al resistance and growth on acid soils.9,10 In Arabidopsis, Al-ACTIVATED MALATE TRANSPORTER 1 (AtALMT1, the major contributor of Al resistance in Arabidopsis) and MULTI-DRUG and TOXIC COMPOUND EXTRUSION (MATE) respectively encode malate and citrate efflux channels/transporters conferring its resistance to Al toxicity.11,12 The Al-induced expression of both genes is exclusively controlled by the zinc finger transcription factor SENSITIVE TO PROTON TOXICITY 1 (STOP1), the central regulator of Al resistance.12,13 STOP1 has widespread conservation of function in Al resistance in different plant species.14,15 Whilst STOP1 mRNA abundance is largely unresponsive to Al,13 Al promotes STOP1 protein accumulation in root cell nuclei, and its accumulation is regulated by the F-box protein REGULATION OF ATALMT1 EXPRESSION 1 (RAE1), which targets STOP1 degradation via the ubiquitin-26S proteasome pathway.16 Additionally, STOP1 is also regulated by SUMOylation and phosphorylation.17,18 Nevertheless, how Al ions are perceived and then connected to the accumulation of the STOP1 central regulator remains unknown.
Plant receptor-like kinases (RLKs), function as cell surface receptors for steroid hormone, chemical or peptide signals.19,20,21,22,23 We hence wondered whether RLKs might similarly serve in Al perception and/or signaling. A typical RLK consists of a ligand-binding extracellular domain, a single transmembrane domain, and a cytoplasmic serine/threonine kinase domain.24 The Arabidopsis genome contains > 600 genes encoding RLK,24 most of which have not been functionally characterized. In this study, we identified a novel Al resistance gene (here named Al Resistance 1, ALR1) encoding a typical leucine-rich-repeat (LRR) RLK, which was previously known as the phytosulfokine (PSK) peptide receptor kinase (PSKR1) involved in multiple processes,25 such as root growth and biotic stress response.26,27,28 We show here that this kinase is unexpectedly an Al ion receptor, and that the Al sensing conferred is physically and functionally separate from PSK sensing. In essence, our work defines a unique plant Al ion receptor ALR1, and reveals how its perception of Al ions is linked via NADPH oxidase RbohD, reactive oxygen species (ROS), RAE1 and STOP1 to the promotion of organic acid anion extrusion-dependent Al resistance.
Results
ALR1 confers Al resistance
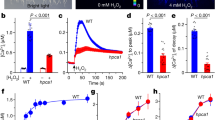
Screening a library of RLK T-DNA insertion mutants, we identified SALK_008585 (alr1-1) displaying remarkably reduced Al resistance (Fig. 1a, b). Whilst ALR1 is known to be required for root growth,28 the morphological development of alr1-1 appears largely identical to that of wild type (WT) (Supplementary information, Fig. S1a). Despite the relatively shorter roots of alr1-1 (vs WT) in control conditions, Al-mediated root growth inhibition is substantially greater in alr1-1 and alr1-2 (SALK_071659C) than in WT (Fig. 1a, b; Supplementary information, Fig. S1b, c). The reduced Al resistance of alr1-1 was restored to normal by transgenic expression of ALR1 (driven by the native ALR1p promoter; ALR1/alr1-1#1 and #2), and Al resistance was promoted by overexpression of ALR1 (ALR1ox1 and ox2; Fig. 1a, b; Supplementary information, Fig. S1d). This increased Al sensitivity of alr1-1 is not attributed to its deficit in root development, as other root-deficient mutants showed WT-like Al response (Supplementary information, Fig. S1e, f). The reduced Al resistance of alr1-1 was next confirmed in hydroponic and soil conditions (Supplementary information, Fig. S1g–k). Further analyses indicated that ALR1 is expressed in root and shoot, and that ALR1 mRNA level (in WT plants) is relatively unaffected by low pH or Al treatments (Supplementary information, Fig. S2a, b). Additionally, the lack of ALR1 did not significantly affect plant responses to other metal ions (Supplementary information, Fig. S2c, d). We conclude that ALR1 confers ion-specific resistance to Al stress.

a, b Root growth under control and Al (1 mM) treatments (a), and their relative quantification (b) (n = 17–21). The average length of each genotype was set to 100%, and the relative root length was expressed as a percentage (root length with Al treatment/root length without Al × 100). almt1 was used as a positive control. c–e Al content in 1 cm root tips (c), root sap (d) and root cell walls (e) under Al (25 µM) treatment (n = 9 in c, n = 8 in d, e). f Expression of ALMT1 in roots under control and Al (25 µM) treatments (n = 3). g Malate secretion from roots under control and Al (50 µM) treatments (n = 4). h Venn diagram showing the overlap of differentially expressed genes (foldchange > 1.5) between WT vs alr1-1 and WT vs stop1 under Al treatment. i, j GFP-STOP1 fluorescence signals in roots (i) and their relative quantification under Al treatment (j) (n = 20). Bars = 1 cm (a), 100 µm (i). All data were analyzed by unpaired t-test (b–f, g, j), or two-way ANOVA (f) (ns non-significance, **P < 0.01, ***P < 0.001, ****P < 0.0001).
We next found both root and shoot Al contents of Al-treated alr1-1 seedlings were significantly increased vs WT, so were the symplastic and apoplastic Al contents in root, whilst those of ALR1ox1 seedlings were significantly decreased (Fig. 1c–e; Supplementary information, Fig. S2e), suggesting that ALR1 confers Al resistance via promoting Al exclusion. These effects of ALR1 are Al-specific, because the accumulation of other ions is largely unaffected in roots lacking ALR1 (Supplementary information, Fig. S2f). ALMT1- and MATE-mediated organic acid anion (malate and citrate) secretion is pivotal for Arabidopsis Al exclusion and resistance,11,12 and we accordingly found that ALMT1- and MATE-encoding mRNA abundances are reduced in alr1-1 roots, but increased in ALR1ox1 roots compared to WT upon Al treatment (Fig. 1f; Supplementary information, Fig. S3a). In consequence, malate and citrate secretion were reduced in alr1-1, but increased in ALR1ox1 vs WT upon Al treatment (Fig. 1g; Supplementary information, Fig. S3b). We conclude that ALR1 confers Al resistance by promoting the exclusion of Al from roots via the enhancement of Al-induced organic acid anion secretion.
ALR1-promoted Al resistance is STOP1-dependent
Al-induced ALMT1 and MATE expression is controlled by STOP1, the central transcriptional regulator of Al resistance.12,13 We next found ALR1 and STOP1 expression to be coordinated (Supplementary information, Fig. S3c), and that the enhanced Al resistance of ALR1ox1 was completely suppressed by a stop1 knockout mutant (Supplementary information, Fig. S3d, e). Moreover, an alr1-1 stop1 double mutant displayed an Al sensitivity comparable to that of both single mutants (Supplementary information, Fig. S3f, g). Further analyses revealed that the promotion of Al-induced expression of ALMT1 and MATE by ALR1 is dependent on STOP1, as is the promotion of Al-induced secretion of malate and citrate (Fig. 1f, g; Supplementary information, Fig. S3a, b). An additional transcriptome profiling showed that the expression of 64.5% genes regulated by STOP1 was likewise regulated by ALR1 under Al treatment (Fig. 1h). We conclude that ALR1-promoted Al resistance is STOP1-dependent.
Although STOP1 mRNA levels were comparable in WT and alr1-1 (Supplementary information, Fig. S3h), we found that Al-induction of GFP-STOP1 accumulation in root apex nuclei was greatly reduced in alr1-1 (vs WT), but promoted by ALR1 overexpression (Fig. 1i, j; Supplementary information, Fig. S3i), by using a STOP1p:GFP-STOP1 transgenic line.
However, unlike stop1 which is insensitive to phosphate (Pi) starvation in root growth,29 alr1-1 showed a WT-like response (Supplementary information, Fig. S2g, h), indicating that ALR1 is not involved in response to Pi starvation. Furthermore, since the ALR1 orthologue PSKR2 functions redundantly with ALR1 in PSK signaling,30 we detected a relatively minor (relative to that of ALR1) contribution of PSKR2 to Al resistance (Supplementary information, Fig. S3j, k). Nevertheless, as in alr1-1, Al-induced STOP1 accumulation was not completely abolished in an alr1-1 pskr2 double mutant (Supplementary information, Fig. S3l, m), perhaps suggesting additional functional redundancy or the existence of minor ALR1-independent mechanisms via which Al promotes STOP1 accumulation. We conclude that ALR1 is predominantly required for the Al-induced accumulation of STOP1.
RbohD-dependent ROS promote STOP1 accumulation
With ALR1 failing to interact with STOP1 (Supplementary information, Fig. S4a), upon screening of ALR1 interactants in a split-ubiquitin membrane yeast two-hybrid system, we identified 18 potential interactors (Supplementary information, Table S1). Testing mutants lacking these potential interactors for Al sensitivity, we found that loss of RbohD function (rbohD-1 and rbohD-2) showed substantially reduced Al resistance, a phenotype rescued by transgenic complementation with RbohD (Fig. 2a, b), suggesting that RbohD is a likely ALR1 substrate. We then confirmed the ALR1–RbohD interaction in yeast and planta (Supplementary information, Fig. S4b–d). Further experiments with additional root-expressed Rboh family members detected relatively weak ALR1–RbohA and ALR1–RbohE interactions (Supplementary information, Fig. S4e), and almost no effect on Al sensitivity (data not shown). Furthermore, the Al-induced accumulation of STOP1 was substantially compromised in the rbohD mutant (Fig. 2c, d; Supplementary information, Fig. S4f), as was Al-induced expression of ALMT1 and MATE (Supplementary information, Fig. S4g, h), resulting in decreased Al-induced malate secretion (Supplementary information, Fig. S4i). These results in aggregate indicate that RbohD promotes Al resistance by enhancing the Al-induced accumulation of STOP1.

a, b Root growth under control and Al (1 mM) treatments (a), and their relative quantification (b) (n = 20). The average length of each genotype was set to 100%, and the relative root length was expressed as percentage (root length with Al treatment/root length without Al × 100). c–f GFP-STOP1 fluorescence signals in roots (c, e) and their relative quantification under indicated treatments (d, f) (n = 20). g, h ROS visual signals in roots detected by H2DCF-DA (g), and their relative quantification under 10 min Al treatment (h) (n = 20). Bars = 1 cm (a), 100 µm (c, e, g). Data were analyzed by unpaired t-test (b, d, f, h) (ns non-significance, ****P < 0.0001).
Because RbohD is an NADPH oxidase responsible for ROS generation,31 we next determined whether ROS promotes Al-induced STOP1 accumulation, finding that the NADPH oxidase inhibitors diphenyleneiodonium chloride (DPI) and imidazole and the ROS scavenger N-acetyl-L-cysteine (NAC) all suppressed Al-induced STOP1 accumulation (Fig. 2e, f; Supplementary information, Fig. S4j). In contrast, methylviologen (MV), which stimulates intracellular ROS generation, and hydrogen peroxide (H2O2) both induced STOP1 accumulation in the absence of Al (Fig. 2e, f; Supplementary information, Fig. S4j). In addition, ALR1ox-promoted STOP1 accumulation was substantially suppressed by DPI and NAC, whilst MV and H2O2 largely restored STOP1 accumulation in alr1-1 and rbohD mutants (Supplementary information, Fig. S4k, l). Accordingly, H2O2 and MV induced the expression of ALMT1 and MATE in the absence of Al (Supplementary information, Fig. S4m, n). Furthermore, we found ROS levels to be significantly increased 10 min after onset of Al treatment in WT root apices, but not in roots of plants lacking ALR1 or RbohD (Fig. 2g, h). We conclude that RbohD-dependent ROS production is necessary for Al-induced STOP1 accumulation, and thus for Al resistance.
ALR1 phosphorylates RbohD to boost ROS production
Because ALR1 can interact with RbohD (Fig. S4b–d), we next tested if ALR1 could phosphorylate RbohD. We found that the recombinant ALR1 cytoplasmic domain (fused with a trigger factor tag, TF-ALR1CD; the TF tag is a 48 kDa chaperone that helps to decrease protein misfolding; Supplementary information, Fig. S5a) indeed phosphorylated an N-terminal fragment of RbohD (TF-RbohDN) in vitro (Supplementary information, Fig. S5a, b). Although initial mass spectrum analyses failed to identify the RbohD phosphorylation site, we subsequently found that a small region of RbohDN had been inadvertently omitted from these analyses (Supplementary information, Fig. S5c). This region contains Ser39 (S39), a phosphosite that plays a key role in RbohD activation during the immune reaction.32 Focusing therefore on S39, we found that a S39 to Ala substitution (RbohDN-S39A) substantially reduced ALR1CD-mediated phosphorylation of RbohDN (Supplementary information, Fig. S5b), indicating S39 to be a major potential target for ALR1-dependent RbohD phosphorylation. We next employed previously described pS39 antibodies specially recognizing the phospho-S39 form of RbohD,32 finding that RbohDN, but not RbohDN-S39A, was in vitro phosphorylated by ALR1CD (Supplementary information, Fig. S5d).
Further analyses revealed that Al treatment promotes ALR1CD-mediated phosphorylation of RbohD S39, and that this promotion increases with increasing Al concentration (Fig. 3a). This effect is relatively specific to Al, because other metal ions, in particular lanthanum (La3+; an ion structurally similar to Al3+), do not promote ALR1-dependent RbohD S39 phosphorylation (Supplementary information, Fig. S5e, f). Finally, phosphorylation was abolished when RbohDN was incubated with a kinase-dead variant ALR1CD (ALR1CD-K762R; Supplementary information, Fig. S5g). These observations suggest that the ALR1 cytoplasmic domain, likely mainly the kinase domain, specifically senses Al ion concentration and proportionately phosphorylates RbohD in response.

a in vitro phosphorylation of RbohD in response to different concentrations of Al (0, 1, 10, 20 and 100 nM) detected with pS39 antibodies. CBS indicates coomassie blue staining. b in vivo phosphorylation of RbohD in seedlings following control or Al (50 µM) treatments. c, d Root growth under control and Al (1 mM) treatments (c), and their relative quantification (d) (n = 20). The average length of each genotype was set to 100%, and the relative root length was expressed as a percentage (root length with Al treatment/root length without Al × 100). e, f GFP-STOP1 fluorescence signals in roots (e) and their relative quantification under Al treatment (f) (n = 20). Bars = 1 cm (c), 100 µm (e). Data were analyzed by unpaired t-test (d, f) (ns non-significance, ****P < 0.0001).
We next found that Al enhances RbohD S39 phosphorylation in WT, but not in alr1-1 (Fig. 3b), indicating that ALR1 is required for in vivo Al-activated RbohD S39 phosphorylation. In addition, transgenic expression of mutant RbohDS39A did not restore Al resistance in rbohD plants (vs WT; Fig. 3c, d), whilst lack of RbohD or expression of RbohDS39A suppressed Al resistance conferred by overexpression of ALR1 (Fig. 3c, d). In contrast, plants expressing a constitutive RbohD phosphorylation mimic (RbohDS39D) exhibited increased Al resistance in the rbohD background vs WT, and rescued Al sensitivity of alr1-1 (Supplementary information, Fig. S6a–d). Furthermore, Al-induced STOP1 accumulation and malate secretion was reduced by RbohDS39A expression (Fig. 3e, f; Supplementary information, Figs. S4i, 5h), but increased by RbohDS39D expression (Supplementary information, Fig. S6e–g). These observations confirm that RbohD is epistatic to ALR1, and that Al-induced ALR1-mediated phosphorylation of RbohD S39 promotes STOP1 accumulation, thus conferring Al resistance.
Further analyses revealed that Al treatment stimulated NADPH oxidase activity in WT plants, but not in alr1-1 or rbohD mutants, or RbohDS39A-expressing plants (Supplementary information, Fig. S5i). Accordingly, Al-promoted ROS generation was abolished in rbohD and RbohDS39A-expressing plants (vs WT; Supplementary information, Fig. S5j, k), but increased in RbohDS39D-expressing plants (Supplementary information, Fig. S6h, i). We conclude that ALR1 confers Al resistance via Al-dependent phosphorylation of RbohD, which in turn stimulates RbohD-dependent ROS generation and associated STOP1 accumulation.
ALR1 function requires SERK co-receptors
Because the interaction of ALR1 with somatic embryogenesis receptor-like kinase (SERK) co-receptors is required for PSK signaling,28 we next found that lack of SERK3/BAK1 (serk3/bak1-4 mutant), but not of SERK1 or SERK2, obviously reduced Al resistance vs WT, although this reduction was less than that conferred by serk1+/– serk2–/– serk3–/– triple mutant (Fig. 4a, b). Moreover, the Al resistance and Al-induced ALMT1 expression and malate secretion were all reduced in bak1-4 and serk1+/– serk2–/– serk3–/– triple mutants vs WT, and the reduced levels in the triple mutant were comparable to those in alr1-1 mutant (Fig. 4c–f). With BAK1 being a major SERK in Al response, we next found that the Al-induced STOP1 accumulation was substantially reduced in bak1-4 vs WT, although the reduction was less than in alr1-1 (Fig. 4g, h). Further experiments demonstrated that Al promotes the interaction between ALR1 and BAK1, and does so in an ALR1 cytoplasmic domain-dependent manner (Fig. 4i–k), thus boosting ALR1-BAK1 inter-phosphorylation (Fig. 4l). Additionally, BAK1, whilst unable to itself phosphorylate RbohD, facilitates Al-promoted ALR1-dependent phosphorylation of RbohD (Fig. 4m). In accord with the previous demonstration that the PSK peptide promotes in vivo ALR1–BAK1 interactions,28 we also found that PSK application increases Al-dependent RbohD phosphorylation (Fig. 4n). We conclude that the ALR1–BAK1 interaction is required for a complete Al-promoted ALR1-dependent RbohD phosphorylation, and is thus involved in Al resistance.

a–d Root growth under control and Al treatments (a, c), and their relative quantification (b, d) (n = 19–24). The average length of each genotype was set to 100%, and the relative root length was expressed as a percentage (root length with Al treatment/root length without Al × 100). e Expression of ALMT1 in roots under control and Al (25 µM) treatments for 6 h (n = 3). f Malate secretion from roots under control and Al (25 µM) treatments for 24 h (n = 3). g, h GFP-STOP1 fluorescence signals in roots (g) and their relative quantification (h) (n = 15). i Bimolecular fluorescence complementation (BiFC) mediated detection of interaction between ALR1 and BAK1, ALR1ED-TM-PSYR1CD (a chimeric ALR1 comprised of ALR1 extracellular and transmembrane domains and the cytoplasmic domain of another RLK PSY1R) and BAK1, or ALR1 and BAK1ED-TM (BAK1 extracellular domain and transmembrane domain), respectively, following control, Al (50 µM) or La (50 µM) treatments for 10 min. j Relative fluorescence intensity in i (n = 40). k Co-immunoprecipitation (Co-IP) showing the interaction between ALR1 and BAK1 in protoplasts following control or Al (50 µM) treatments for 10 min. l Inter-phosphorylation of ALR1CD and BAK1CD in response to different concentrations of Al (0, 10, 50 and 100 nM) in vitro. m BAK1CD promotes the phosphorylation of RbohDN by ALR1CD under both control and 100 nM Al treatment conditions. The arrow indicates a phosphorylated RbohDN band detected with pS39 antibodies. The asterisk indicates that pS39 antibodies can also unexpectedly recognize the BAK1 cytoplasmic domain (but cannot recognize other cytoplasmic domains, data not shown). n The effect of PSK (3 µM) on in vivo RbohD phosphorylation. Bars = 1 cm (a, c), 100 µm (g), 20 µm (i). All data were analyzed by unpaired t-test (b, d–f, h, j) (ns non-significance, *P < 0.05, **P < 0.01, ****P < 0.0001).
ROS modify RAE1 to facilitate STOP1 accumulation
Because STOP1 proteolysis is mediated by the RAE1 F-box protein,16 we next determined whether ROS affect this process. By ectopically expressing STOP1-GFP in mesophyll protoplasts (which do not natively express STOP1, even in response to Al), we found that whilst co-expression of RAE1 reduced STOP1-GFP accumulation, this reduction was suppressed by MV, with MV-induced suppression being in turn inhibited by NAC (Supplementary information, Fig. S7a, b). These observations suggest that ROS directly inhibits RAE1-mediated STOP1 proteolysis.
ROS signaling is often mediated via post-translational modification of cysteine (Cys) residues with low pKa.33 Biotin-conjugated iodoacetamide (BIAM) specifically competes with ROS in reacting with target Cys residues, enabling ROS-sensitive Cys residues to be detected by BIAM labeling.34 We found weakly BIAM-modified recombinant STOP1 in vitro (Supplementary information, Fig. S7c). Although subsequent mass spectrum analyses revealed 3 major potential ROS-modified STOP1 Cys residues (Cys27, Cys185 and Cys335), mutational substitution of these Cys residues with Ala residues did not detectably affect either RAE1-mediated STOP1 degradation or ROS-mediated suppression of STOP1 proteolysis (Supplementary information, Fig. S7d), implying that ROS-mediated inhibition of RAE1-dependent STOP1 proteolysis is unlikely to be dependent upon post-translational modification of STOP1.
We next identified strong H2O2-competable modification of recombinant RAE1 in vitro (Fig. 5a), and found that BIAM labeling of in vivo FLAG-RAE1 was suppressed in WT by Al treatment, but not in alr1-1 (Fig. 5b), suggesting that Al promotes the oxidative modification of RAE1 in an ALR1-dependent manner. Mass spectrum analyses further detected 9 BIAM-modified RAE1 Cys residues (Supplementary information, Fig. S7e, f), among which Cys364 is predominantly required for RAE1-promoted STOP1 degradation (Fig. 5c, d; Supplementary information, Fig. S7g, h). Accordingly, STOP1 ubiquitination was substantially reduced by RAE1C364A co-expression (vs WT RAE1 co-expression; Fig. 5e). Intriguingly, the Cys364-to-Ala364 substitution did not detectably affect the in vivo interaction between RAE1 and STOP1 (Supplementary information, Fig. S7i, j), indicating that Cys364, although necessary for RAE1 function, is not necessary for RAE1–STOP1 interaction. Confirming the role of Cys364 in RAE1 function in vivo, we found that transgenic expression of RAE1C364A in a rae1-1 loss-of-function mutant (RAE1C364A/rae1-1) enabled STOP1 accumulation in the absence of Al to a level similar to that in the rae1-1 control, whilst STOP1 accumulation was not detected in RAE1/rae1-1 (Fig. 5f, g; Supplementary information, Fig. S7k). Overall, these results demonstrated that Al-induced ALR1-promoted ROS generation enhances STOP1 accumulation via oxidative modification of RAE1 Cys364.

a in vitro labeling of BIAM for recombinant His-RAE1. b in vivo labeling of BIAM for FLAG-RAE1 immuno-precipitated from seedlings following 20 min control and Al (50 µM) treatments. c STOP1-GFP expression in mesophyll protoplasts with or without co-expressing RAE1 or mutated RAE1 (RAE1C364A). d Abundance of STOP1-GFP in protoplasts detected by α-GFP antibody. e Ubiquitination of STOP1-GFP in protoplasts detected by α-Ub antibody. f, g GFP-STOP1 fluorescence signals in roots (f) and their relative quantification under control condition (g) (n = 20). Bars = 100 µm (c, f). Data were analyzed by unpaired t-test (ns non-significance, ****P < 0.0001).
ALR1 is an Al ion receptor
Because ALR1CD-mediated phosphorylation of RbohD is responsive to Al concentration (Fig. 3a, b), we next determined if ALR1 binds Al ions specifically. Using a microscale thermophoresis (MST) assay to detect binding to a recombinant ALR1CD protein, we found that Al ions display a high binding affinity for ALR1CD (Fig. 6a; dissociation constant ranges from 0.2 µM to 2 µM upon different batches of experiments). This binding was confirmed via isothermal titration calorimetry (Supplementary information, Fig. S8a–e). The binding affinity may partially rely on its kinase activity, as a kinase-dead mutation (ALR1CD-K762R) increases the dissociation constant by 5–10 fold (Supplementary information, Fig. S8f). Moreover, ALR1CD did not detectably bind other metal ions (Fig. 6b), suggesting that ionic binding to ALR1CD is Al-specific. We additionally showed that the BAK1 cytoplasmic domain does not bind Al ions (Fig. 6a), indicating that BAK1 is not directly involved in Al perception. Furthermore, a truncated ALR1 (spanning ALR1 residues 658 to 1008) lacking the extracellular domain is sufficient to restore Al resistance in alr1-1 knockout mutant (ALR1CD/alr1-1; Fig. 6c, d), favoring that the intracellular ALR1 cytoplasmic domain is responsible for Al perception.

a Quantification of the binding affinity of ALR1CD, mutated ALR1CD (ALR1CD4C-A) and BAK1CD for Al ions by MST (n = 4 for ALR1CD, n = 3 for ALR1CD4C-A and BAK1CD). b Quantification of the binding affinity of ALR1CD for metal ions other than Al by MST. c, d Root growth under control and Al treatment (c), and their relative quantification (d) (n = 20). The average length of each genotype was set to 100%, and the relative root length was expressed as a percentage (root length with Al treatment/root length without Al × 100). e Phosphorylation of RbohDN by ALR1CD or ALR1CD4C-A in response to Al. f, g GFP-STOP1 fluorescence signals in roots (f) and their relative quantification under Al treatment (g) (n = 20). Bars = 1 cm (c), 100 µm (f). All data were analyzed by unpaired t-test (d, g) (ns non-significance, ****P < 0.0001).
Since many amino acid residues (e.g., Asp, Glu, Asn, Cys, etc.) can be the candidates for metal ion targets,35 we focused initially on the least distributed Cys residues in the ALR1 cytoplasmic domain, and determined if the 8 Cys residues contribute to Al perception. Mutational analyses of these Cys groups (but not of Cys797, where mutation failed) revealed that substitution of Cys939/944 or Cys985/987 with Ala, affected the Al-binding of ALR1CD (Supplementary information, Fig. S8g–l), and that a simultaneous quadruple Ala substitution (Cys-939/944/985/987-Ala; ALR1CD4C-A) completely abolished the binding (Fig. 6a), indicating that the 4 Cys residues are necessary for ALR1 to recognize Al ions.
The above identified 4 Cys residues (Cys-939/944/985/987) are located in a C-terminal region of the ALR1 cytoplasmic domain (Supplementary information, Fig. S8h). To exclude the possibility that mutation of these Cys residues affects the basal kinase activity or overall structural integrity of ALR1CD, we next found the auto-phosphorylation of recombinant ALR1CD and ALR1CD4C-A to be comparable, as is the phosphorylation of RbohD and BAK1 by ALR1CD and ALR1CD4C-A (Supplementary information, Fig. S9a–c). Additionally, transgenic expression from the native ALR1 promoter of mutant ALR14C-A (encoding a full-length ALR1 with Cys939/944/985/987 all substituted by Ala) in alr1-1 (ALR14C-A/alr1-1) conferred a normal PSK response (as compared with ALR1/alr1-1#1 and alr1-1), whilst the kinase-dead control (ALR1K762R/alr1-1) displayed an abolished PSK response (Supplementary information, Fig. S9d, e). Finally, structural integrity determinations using circular dichroism (CD) revealed the structures of mutant (ALR1CD4C-A) and WT (ALR1CD) proteins to be almost identical (Supplementary information, Fig. S9f, g). These data demonstrate that substitution of these four Cys residues (Cys939/944/985/987) with Ala does not detectably affect the structural integrity or kinase activity of the ALR1 cytoplasmic domain.
We next found that Al-promoted phosphorylation of RbohD was largely abolished by ALR1CD4C-A, and that ALR1CD4C-A kinase activity, unlike that of ALR1CD, was actually increasingly inhibited by increasing Al concentration (Fig. 6e), indicating that these 4 Cys residues are essential for Al-promoted ALR1 kinase activity, and that the disruption of Al binding from their substitution with Ala may cause ALR1CD to be more vulnerable to toxic Al ions. Confirming the in vivo function of these 4 Cys residues, we next found that transgenic expression of the mutated ALR14C-A (ALR14C-A/alr1-1) substantially reduced Al resistance (vs WT and ALR1/alr1-1; Fig. 6c, d). Accordingly, Al-activated root apex NADPH oxidase activity and ROS generation were both markedly decreased in ALR14C-A/alr1-1 lines (vs WT), but restored by ALR1CD in alr1-1 (Supplementary information, Fig. S10a–c). Furthermore, Al-induced STOP1 accumulation was substantially reduced in ALR14C-A/alr1-1 plants (vs WT; Fig. 6f, g; Supplementary information, Fig. S10d). These results collectively demonstrate that ALR1 is an Al ion receptor.
ALR1-mediated Al perception is independent of its PSK sensing function
ALR1 senses PSK peptide through its extracellular domain.28 Our further studies revealed that mutation of the PSK peptide-binding sites in ALR1 (in ALR1F506A/alr1-1, ALR1R300A/alr1-1 transgenic lines) does not obviously affect Al resistance, whilst a kinase-dead mutation (in ALR1K762R/alr1-1) does (Supplementary information, Fig. S11), indicating that ALR1-dependent Al perception is likely independent of its PSK sensing function.
Since binding of PSK or Al ion both enhances the interaction between ALR1 and SERK co-receptors, we indeed found that exogenous application of PSK peptide or overexpression of PSK genes increased Al resistance in an ALR1-dependent manner (Supplementary information, Fig. S12a–d), and that PSK overexpression promotes STOP1-dependent Al signaling (Supplementary information, Fig. S12e–h). Nevertheless, PSK cannot induce Al signaling (e.g., ROS production, STOP1 accumulation, ALMT1 expression) in the absence of Al (Supplementary information, Figs. S12e–g, S13a, b), and PSK promotes root growth independently of RbohD (Supplementary information, Fig. S13c, d). These suggest that although they share same receptor and co-receptors, Al and PSK signaling pathways are separate.
In summary, we demonstrate that ALR1 is an Al ion receptor. Binding of Al ions to the ALR1 cytoplasmic domain promotes ALR1-mediated phosphorylation of RbohD, thus increasing ROS production. The consequent ROS-dependent accumulation of STOP1 stimulates organic acid anion secretion and results in enhanced Al resistance.
Discussion
In this study, we report the unique discovery that the plant ALR1 receptor-like kinase is an Al ion receptor. Our discovery rests on the following evidence: (1) the cytoplasmic domain of ALR1 specifically binds Al ions; (2) the binding of Al to ALR1 recruits BAK1 co-receptor and promotes ALR1-mediated phosphorylation of RbohD, thus stimulating an Al-dependent elevation of ROS generation and signaling; (3) ALR1-dependent RbohD phosphorylation is quantitatively responsive to varying Al concentration and confers commensurate quantitative regulation of STOP1 accumulation; (4) abolition of the ALR1 Al binding function substantially suppresses the predominant STOP1-dependent Al signaling pathway.
Plants constantly sense and respond to a wide diversity of soil ions, including nutrient and toxic ions. Nevertheless, the mechanisms by which plants initially sense these different ions and transduce these signals remain poorly understood. Recent sensor/receptor discoveries have advanced understanding of how plants adaptively sense and respond to a variety of hormonal, chemical and physical cues, thus optimizing growth, survival and reproductive success,21,36,37,38 and the discovery of plant ion receptors will be similarly revealing. The finding that the plant ALR1 kinase is an Al ion receptor is unique because in mammals, ions are mostly sensed by ion channels. For example, mice sense sodium via the sodium ion (Na+) channel ENaC, and lack of ENaC in taste cells blocks mouse neural and behavioral Na+ responses.39 Furthermore, although the nitrate ion is at least partially sensed by the plant nitrate transporter NRT1.1,40 plant ion perception is more generally thought to be achieved via non-channel ion receptors. Accordingly, initial Na+ sensing by the sphingolipid GIPC subsequently activates Arabidopsis Ca2+ signaling,41 the intracellular Fe3+ is potentially sensed by HRZ/BTS RING ubiquitin ligases,42 and Zn2+ is likely to be sensed by plant bZIP transcription factors.43 Recent research has revealed that nitrate is also sensed by NLP7 transcription factor.44 So far as we are aware, our discovery that ALR1 is an Al ion receptor is unique because no kinase-type receptor has previously been shown to sense an inorganic or metal ion in all living organisms. Furthermore, in contrast to other RLKs, which perceive signals via their extracellular domains,45 ALR1 senses intracellular Al ions via its intracellular cytoplasmic domain, likely mainly the kinase domain. The binding of Al ions to the ALR1 cytoplasmic domain promotes ALR1-BAK1 association and inter-phosphorylation (Fig. 4i–l), suggesting a special mechanism for RLK activation. These findings thus provide novel insights into how living organisms perceive and respond to ions.
We have shown that a set of four C-terminal Cys residues (Cys939/944/985/987) are necessary for the Al binding function of ALR1 (Fig. 6). Nevertheless, our structural model suggests that they are unlikely to form an Al ion coordination center in space (Supplementary information, Fig. S8m), and their substitutions with Ala are more likely to indirectly affect Al binding. We propose that the ALR1 Al binding sites likely reside within the region spanned by Cys939 and Cys987. Although we have recently defined the Al binding sites of the ALMT1 protein via structural analysis,46 it is at present unclear if the ALR1 Al binding sites have a similar composition, and biochemical approaches are unlikely to pinpoint it. Future comparisons of the high-resolution structures of ALR1 (perhaps in complex with BAK1 and RbohD) in both Al-bound and unbound states will likely reveal how Al ions bind to ALR1 and how this binding promotes ALR1 kinase activity.
Our results also show that ALR1-mediated Al signaling (via its cytoplasmic domain) is functionally separate from ALR1-mediated PSK signaling (via its extracellular domain) (Fig. 6c, d; Supplementary information, Figs. S11c, d, S13a–d). However, the possibility, that these functionally separate Al and PSK signaling pathways interact, needs to be considered. For example, we here show that Al treatment represses the expression of most PSK-encoding genes (Supplementary information, Fig. S13E), implying that the PSK peptide is unlikely to explain Al resistance in nature. Because PSK signaling promotes root growth,47 the Al-induced repression of PSK expression may be an adaptive response to reduce the exposure of roots to toxic Al ions. Therefore, ALR1 may have dual functions concerning root growth under Al stress conditions, with the extracellular domain controlling normal root growth and the cytoplasmic domain regulating the Al stress response. In contrast, cytoplasmic kinase-domain-mediated Al signaling detoxifies Al and protects root tips from Al-induced cellular damages. Once Al toxicity is relieved, root growth recovers as PSK signaling is restored. ALR1 thus represents a so-far unique RLK that perceives extracellular and intracellular signals via distinct extracellular and intracellular protein domains to coordinate developmental and stress responses. Given the dual function of ALR1, we propose to name the gene PSKR1/ALR1 (Al Receptor 1) for its future identity. Nevertheless, how PSKR1/ALR1 transduces PSK or Al signals to different downstream components remains an open question that needs to be addressed in future studies. Besides, since Al-induced STOP1 accumulation is not completely blocked in the alr1-1 and rbohD-1 mutants, additional ALR1-independent pathways may operate to fine-tune STOP1 expression.
Although ROS production in response to Al was previously thought to be Al toxicity syndrome,1 we give the direct evidence that the RbohD-dependent ROS are key second messengers in Al signaling linking membrane-based intracellular Al perception to nuclear response. we identify Cys364 of RAE1 is both oxidatively modified by ROS and necessary for RAE1 function and Al signaling (Fig. 5). Ala-substitution of Cys364 did not detectably affect the RAE1–STOP1 interaction (Supplementary information, Fig. S7i, j), and Ala-substitution assay of additional Cys residues indicated that they are less important for RAE1 function than Cys364, suggesting that Cys364 may function in the formation of an inter-molecular disulfide bond that facilitates the interaction between RAE1 and unknown components of the proteolytic machinery (e.g., the SCF E3 ubiquitin ligase complex). The finding that ROS directly act on the RAE1 F-box protein, the component of cellular proteolytic machinery, provides a novel ROS-signaling mechanism that has not been uncovered in plants.48 In addition, because MV or H2O2 promotes STOP1 accumulation in the absence of Al (Fig. 2e), any treatment that induces ROS production would likely more or less prevent STOP1 degradation and induce the expression of ALMT1 and MATE genes. However, this would not lead to a large amount of malate and citrate release, as these transporters/channels still require extracellular Al3+ to trigger their activity.11,46
In conclusion, we have discovered ALR1, an LRR-RLK that is an Al ion receptor, and have revealed all subsequent steps in a ROS-mediated Al signaling pathway leading from ALR1 perception of Al ions to the STOP1 central regulator conferring Al resistance (Fig. 7). These discoveries will inform future molecular breeding of Al resistant crops and trees, and thus have huge potential not only for sustaining agricultural production and global food security, but also for forest restoration and improvement of the global ecological environment.

①Al ions entering root cells are perceived by the cytoplasmic domain of the PSKR1/ALR1 receptor. ②The binding of Al ions enables PSKR1/ALR1 to recruit BAK1, the representative of SERKs, as a co-receptor for inter-phosphorylation and activation, and thus ③promotes ALR1-mediated phosphorylation of the NADPH oxidase RbohD at Ser39, hence increasing ROS generation. ④The resultant accumulated ROS in turn inhibits the function of the RAE1 F-box protein via oxidative modification of Cys364 residue, thus suppressing RAE1-mediated degradation of STOP1, the master transcriptional regulator of Al-resistance genes. The consequent accumulation of STOP1 hence activates the expression of downstream genes, including the malate channel-encoding ALMT1 gene, finally promoting organic acid anions secretion to chelate extracellular Al ions, thus conferring Al resistance. OA represents organic acid anions.
Materials and methods
Plant materials and growth conditions
Arabidopsis thaliana lines are in Col-0 genetic background. alr1-1 (SALK_008585), alr1-2 (SALK_071659C), rbohD-1 (SALK_005253C) and stop1 (SALK_114108) were obtained from Arabidopsis Biological Resource Center (ABRC). rbohD-2, RbohDS39A/rbohD-2, rae1-1, serk1, serk2, serk3 (bak1-4), serk1+/– serk2–/– serk3–/–, and STOP1p:GFP-STOP1/stop1 were described previously.16,28,29,32,49 STOP1p:GFP-STOP1 lines in genetic background of alr1-1, bak1-4, alr1-1/pskr2, ALR1ox1, rbohD-1, RbohDS39A/rbohD-2, RbohDS39D/rbohD-1 and rae1-1, were generated by crossing STOP1p:GFP-STOP1/stop1 to alr1-1, bak1-4, alr1-1/pskr2, ALR1ox1, rbohD-1, RbohDS39A/rbohD-2, RbohDS39D/rbohD-1 and rae1-1, respectively. The short-root (shr) and rgi1 rgi2 mutants that are deficient in root growth were used as the negative controls. SHR is a transcription factor that regulates root radial patterning and stem cell niche maintenance,50 and RGI1/2 are receptor-like kinases responsible for RGF peptide perception in regulation of root stem cell niche.51 All the mutants were identified by PCR and confirmed by Sanger sequencing.
Plants used for treatment or propagation were grown in an environmental controlled growth chamber or room programmed for a 16-h-light/8-h-dark cycle with a daytime temperature of 23 °C and a night temperature of 21°C. For plants used for mesophyll protoplast isolation, the condition of a 10-h-light/14-h-dark cycle was conducted. The half-strength Murashige & Skoog (MS) medium was prepared for normal seedling growth as previously described.52
Constructs and transgenic plants
For ALR1 transgenic complementation test, the coding sequences of full-length ALR1 (3080 bp) and truncated ALR1 (ALR1CD, 1053 bp) driven by ALR1 native promoter (3080 bp) were cloned into pCambia1301 binary vector to generate ALR1p:ALR1 and ALR1p:ALR1CD, respectively. The ALR1p:ALR14C-A, ALR1p:ALR1R300A, ALR1p:ALR1F506A and ALR1p:ALR1K762R constructs were generated by site-directed mutagenesis in ALR1p:ALR1 using the KOD-Plus-Mutagenesis Kit (TOYOBO, SMK-101), respectively. These constructs were transformed into alr1-1 by Agrobacteria strain GV3101-mediated transformation. For generating ALR1ox1 and ox2, the full-length ALR1 coding sequence was cloned into pCAMBIA1301-35S and then transformed into WT. For generating ALR1p:GUS, the same ALR1 native promoter was cloned into pCambia1301-GUS and then transformed into WT. For RbohD transgenic complementation test, a full-length RbohD coding sequence (2766 bp) driven by its native promoter (2448 bp) was cloned into pCambia1301 to generate RbohDp:RbohD. The RbohDp:RbohDS39D construct was generated by site-directed mutagenesis in RbohDp:RbohD. These constructs were then transformed into rbohD-1 background. For RAE1 complementation test, the coding sequences of full-length RAE1 (1998 bp) and mutated RAE1 (RAE1C364A) driven by RAE1 native promoter (1740 bp) were cloned into pCAMBIA1301 to generate RAE1p:RAE1 and RAE1p:RAE1C364A, respectively, and were further transformed into rae1-1/GFP-STOP1 to generate RAE1/rae1-1/GFP-STOP1 and RAE1C364A/rae1-1/GFP-STOP1 transgenic lines, respectively. For generating ALR14C-A/alr1-1/STOP1-GFP lines, the coding sequence of full-length STOP1 (1500 bp) fused with GFP (720 bp) was cloned into pCAMBIA1301-35Sp to generate 35Sp:STOP1-GFP construct, which was then transformed into Col-0 and ALR14C-A/alr1-1, respectively. To generate PSK4ox and PSK5ox, the coding sequence of full-length PSK4 and PSK5 were cloned into pCAMBIA1301-35Sp, respectively, and were further transformed into Col-0. PSK4ox/alr1-1 and PSK5ox/alr1-1 were obtained by crossing PSK4ox or PSK5ox with alr1-1, respectively. To generate the FLAG-ALR1, FLAG-RbohD and FLAG-RAE1 transgenic lines, the full-length coding sequences of ALR1, RbohD and RAE1 were respectively cloned into the pCAMBIA1300-35S-3×FLAG plasmid. The resulting constructs were introduced into WT and alr1-1 plants by agrobacterium-mediated transformation. Primers used are listed in Supplementary Table S2.
Al treatment
For phenotypical analysis under Al treatment, a soaked gel medium was used to evaluate Al sensitivity as previously described with minor modification.16 1/6 MS agar medium plus 0.5% sucrose and 0.8% agar (Sigma-Aldrich, A7002) was soaked with 25 mL of the same nutrient medium with or without 1 mM AlCl3 at pH 3.6. After 2 d of soaking, the solution was removed, and seeds were grown on the agar medium in plates for 7–10 d. For metal ion treatments in hydroponic culture, seeds were sowed on 1/8 MS with or without 2–5 µM AlCl3, 2 µM CuSO4, 2 µM CdCl2, 10 µM LaCl3, 10 µM FeCl3 at pH 5.2. The seedlings were then photographed and the root lengths were measured with ImageJ software. For data presentation, the average length of each genotype under the control condition was set to 100%, and the relative root length was expressed as a percentage (root length with Al treatment/root length without Al × 100), which was used to evaluate Al sensitivity.
A library of RLK T-DNA insertion mutants (including ~300 RLK mutants) was collected in the preliminary study. The Al sensitivity of each mutant was detected on the Al-soaked gel medium as described above, and alr1-1 was isolated.
For other analysis under Al treatment, roots were pretreated with 0.5 mM CaCl2 (pH 4.5) solution for 2 h, and then treated with the same solution with or without AlCl3 for the indicated time.
For soil experiments, the acid soil (pH 4.3) used in this study was a mix of normal soil (pH 6.5) and acid soil (pH 4.0). To minimize the difference in nutrient levels in the two soils, the full-strength nutrient solution (pH 4.3) was used to irrigate the two soils. Furthermore, the almt1 mutant was used as a positive control showing the Al toxicity in acid soils.
Determination of root Al content
Root samples from 1-week-old seedlings following 24 h Al (50 µM) treatment were used for Al content measurement. The Al content in the whole root, root cell sap, or root cell wall was determined as previously described.53
Detection of root organic acid anion exudation
Both 4-week-old plants and 1-week-old seedlings were used for determination of root organic acid anion secretion. For 4-week-old plants, the secretion of organic acid anions was detected as previously described.48 For 1-week-old seedlings, they were transferred to 6-well plate (100 seedlings per well) and pretreated with 0.5 mM CaCl2 solution containing 1% sucrose for 2 h at pH 4.5, and then were treated with 4 mL of the same solution with or without 50 µM AlCl3 for 24 h with constant shaking on a rotary shaker (50 rpm) at room temperature in darkness. Malate and citrate concentrations in exudation media were determined by NAD/NADH cycling-coupled enzymatic method as described previously.54
Confocal microscopic analysis
For observation of Al-induced GFP-STOP1 fluorescence, 1-week-old STOP1p:GFP-STOP1 seedlings in various genetic backgrounds were treated with or without 50 µM AlCl3 in 0.5 mM CaCl2 solution (pH 4.5) for 1.5 h. For chemical treatment, seedlings were treated with 50 µM AlCl3 in 0.5 mM CaCl2 solution (pH 4.5) plus 50 µM DPI, 50 µM AlCl3 plus 10 mM imidazole, 50 µM AlCl3 plus 50 µM NAC, 10 µM MV and 200 µM H2O2 for 1.5 h, respectively. GFP fluorescence was detected and photographed using a confocal laser scanning microscope (Zeiss) with the same parameter setting. For ROS visualization, 1-week-old seedlings were pre-treated with 100 µM NAC in 0.5 mM CaCl2 solution (pH 4.5) for 8 min (to remove background ROS), and were transferred into fresh 0.5 mM CaCl2 solution with or without 15 µM AlCl3 and 10 µM H2DCF-DA for 10 min. The fluorescence was then detected and photographed using a fluorescence microscope (Nikon). The fluorescence intensities of GFP and ROS were analyzed using software ImageJ. The average fluorescence intensity of GFP-STOP1 or ROS in WT under Al treatment was set to 100%, and the relative fluorescence intensity was expressed as a percentage (fluorescence intensity in different genotypes under Al treatment/fluorescence intensity in WT under Al treatment × 100, or fluorescence intensity in WT under other treatments/fluorescence intensity in WT under Al treatment × 100).
Gene expression analysis
For real-time qPCR, total RNAs were extracted from the roots of 1-week-old seedlings with or without 25 µM Al treatment for 6 h, and converted to cDNAs as previously described. RT-qPCR analysis was carried out using the SYBR Green Realtime PCR Master Mix (TOYOBO) on a Roche LightCycler480 real-time qPCR system following the manufacturer’s instructions. Transcript levels of each mRNA were determined and normalized with the level of UBQ10 mRNAs using the ΔCt method.55
RNA-sequencing (RNA-seq) assay
For RNA-seq assay, total RNAs were extracted from the roots of 1-week-old WT, alr1-1 and stop1 seedlings with or without 200 µM Al treatment for 6 h in 0.5 mM CaCl2 solution at pH 5.2. Three biological replicates were done. The RNA-seq analysis was performed by Novogene Science and Technology Co., Ltd. (Tianjin, China) using the Illumina NovaSeq 6000 Sequencing System (150 bp paired-end reads; 6 G). The differentially expressed genes were identified using edge R (| log2Fold Change | > = 0.6, FDR < 0.05).
Yeast two-hybrid assay
A split-ubiquitin membrane yeast two-hybrid system was used as previously described.56 pPR3-N-RbohD (AD-RbohD) and pPR3-N-STOP1 (AD-STOP1) constructs were either individually transformed or co-transformed with pBT3-STE-ALR1 (BD-ALR1) into NMY51 yeast competent cells according to the manufacturer’s instructions (Clontech). Transformed yeast cells were grown in SD (–Leu/Trp) liquid media to an OD600 of 0.1 and diluted in a 10× dilution series with 0.9% NaCl, and were further spotted on SD double dropout (–Leu/Trp) and SD quadruple dropout (–Ade/His/Leu/Trp) media plates.
Split luciferase (LUC) complementation assay
The 35 S:cLUC-RbohA/C/D/E/F and 35 S:ALR1-nLUC constructs were transferred into the Agrobacterium GV3101 lines individually, and the GV3101 lines were co-infiltrated into the N. benthamiana leaves. Luciferase imaging was performed 48 h post infiltration using a NightShade LB 985 in vivo Plant Imaging System with a CCD camera.
Transient expression in protoplasts
Mesophyll cell protoplasts were isolated mainly as previously described.57 The protoplasts that were transformed with the same constructs and were further exposed to different treatments were from the same pool; otherwise, they were from independent pools. For independent transformation, we used a 35 S:LUC plasmid as an internal control to monitor the transformation efficiency.
To assess RAE1 and RAE1 mutants-mediated STOP1 protein degradation, protoplasts were transfected with HBT-35S:STOP1-GFP and pUC35S-Flag-RAE1or pUC35S-Flag-RAE1m (including RAE1C160A, RAE1C218A, RAE1C262A, RAE1C288A, RAE1C339A, RAE1C365A, RAE1C391A, RAE1C507A, RAE1C364A, RAE19C-A and RAE18C-A). After 12 h incubation at room temperature, the protoplasts were harvested and the STOP1-GFP fluorescence signal was detected using a fluorescence microscope (Nikon). For MV treatment, 10 µM MV was added to the protoplasts after transfection for 6 h.
For immunoblot assay, 1 mL of protoplasts were transfected with 50 µg HBT-35S:STOP1-GFP and 50 µg pUC35S-Flag-RAE1 or pUC35S-Flag-RAE1C364A or pUC35S-Flag-empty. The protoplasts were harvested and lysed in 300 µL of lysis buffer (50 mM Tris-HCl, pH 7.4, 5 mM EDTA, 150 mM NaCl, 1% Trition X-100, 0.1% SDS, 0.5% sodium deoxycholate, 1× protease inhibitor cocktail). After centrifugation at 13,000× g for 10 min, 20 µL of supernatants were separated on 10% SDS-polyacrylamide gels, and the proteins were then transferred to PVDF membrane by wet electroblotting and were further detected by α-GFP (ABclonal) and α-actin (ABclonal) antibodies with dilution of 1:5000, respectively.
To assess RAE1 and RAE1C364A-mediated ubiquitination of STOP1, 1 mL of protoplasts were transfected with 50 µg HBT-35S:STOP1-GFP and 50 µg pUC35S-Flag-RAE1 or pUC35S-Flag-RAE1C364A or pUC35S-Flag-empty. The protoplasts were harvested and lysed in 300 µL of lysis buffer. After centrifugation at 13,000× g for 10 min, 50 µL of the supernatants were kept as input, while the rest was incubated with 20 µL of GFP-Trap (ChromoTek) for 2 h. After the beads were washed three times with TBS buffer, the ubiquitinated conjugates were detected by immunoblot with α-GFP (1:5000; ABclonal) and α-Ub (1:5000; Ubbiotech) antibodies. The input was detected with α-GFP (1:5000; ABclonal) and α-FLAG (1:5000; ABclonal) antibodies.
BiFC assay
To generate constructs for BiFC assay, the full-length ALR1, chimeric ALR1 (comprised of ALR1 extracellular and transmembrane domains (1–680 aa) and the PSY1R cytoplasmic domain (740–1095 aa)), RbohD, the full-length BAK1 and the truncated BAK1 (BAK1ED, comprised of BAK1 extracellular and transmembrane domains (1–249 aa)) fragments were cloned into pUC35S-nYFP or pUC35S-cYFP (BIOGLE GeneTech) vectors to obtain ALR1-nYFP, ALR1ED-TM-PSYR1CD-nYFP, RbohD-cYFP, BAK1-cYFP, and BAK1ED-TM-cYFP, respectively. These plasmids were selectively co-transferred into the Arabidopsis mesophyll protoplasts. Cells were incubated in W5 solution for 10–12 h, and the expression of YFP was detected using a confocal laser scanning microscope (Zeiss). For AlCl3 or LaCl3 treatment, cells from the same pool were treated with 50 µM AlCl3 or 50 µM LaCl3 for 10 min before fluorescence detection. The nYFP or cYFP empty plasmids were used as negative controls.
NADPH oxidase activity assay
For NADPH oxidase activity assay, 1-week-old Arabidopsis WT, alr1-1, rbohD-2, RbohDS39A/rbohD-2, ALR1CD/alr1-1 and ALR14C-A/alr1-1 grown on 1/2 MS media were treated with or without 50 µM AlCl3 in 0.5 mM CaCl2 solution (pH 4.5) for 1.5 h. The seedlings were harvested and lysed in 1 mL of RIPA buffer (50 mM Tris-HCl pH 7.4, 5 mM EDTA, 150 mM NaCl, 1% Triton X-100, 0.1% SDS, 0.5% sodium deoxycholate, 1× protease inhibitor cocktail) and incubated for 1.5 h at 4 °C. Then the lysed cells were clarified by centrifugation at 13,000× g for 10 min, and the supernatants were used to detect NADPH oxidase activity as previously described.
Recombinant protein expression and purification
For recombinant protein expression in E.coli, ALR1CD (2041–3027 bp), RbohDN (1–1130 bp), and their mutant variants (ALR1CD-K762R, ALR1CD-C742A, ALR1CD-C768A, ALR1CD-C852A, ALR1CD-C939/944A, ALR1CD-C985/987A, ALR1CD944A, ALR1CD3C-A, ALR1CD4C-A, TF-RbohDN-S39A and TF-RbohDN-S152A) were amplified and respectively cloned into pCold-TF (TAKAR) vector containing a His-TF-Tag sequence (encoding a 48 kDa chaperone helping decrease protein misfolding). The PCR fragment of BAK1CD (cytoplasmic domain, 76–1971 bp) was cloned into both pCold-TF and GST-tagged vectors. These plasmids were transformed into E. coli BL21 CondonPlus (DE3) strains. Cultures were grown at 37 °C until OD600 = 0.4–0.6, and protein expression was induced by 1 mM IPTG at 16 °C for 16 h. After induction, the bacteria were collected by centrifugation at 5000× g and stored at –80 °C until use.
The His-tagged recombinant proteins were purified using Ni-NTA superflow column (QIAGEN, 30622) according to the manufacturer’s instructions. The purified proteins were finally eluted by 3 mL elution buffer (50 mM Tris-HCl, 300 mM NaCl, 250 mM imidazole, 2 mM DTT, 1× protease inhibitor cocktail). The protein after dialysis was aliquoted and stored at –80 °C until use.
Phosphorylation assay
ATP-γ-S-dependent in vitro phosphorylation assay was performed as described previously.58 The phosphorylation was detected by α-Thiophosphate ester antibody (1:5000; Abcam, ab92570). For pS39 antibodies-dependent in vitro phosphorylation assay, the recombinant proteins were incubated in kinase reaction buffer (20 mM HEPES pH 7.4, 10 mM MgCl2, 1 mM DTT, and 20 mM ATP) with or without AlCl3 (0, 1, 10, 50 and 100 nM) for 30 min at 37 °C and stopped by adding 5× SDS loading buffer. The samples were then separated by 10% SDS-PAGE, and the phosphorylation of His-RbohDN was detected by immunoblot with pS39 antibodies (1:5000). The pS39 antibodies were generated as described previously.32
For in vivo phosphorylation assay, 1-week-old FLAG-RbohD/WT and FLAG-RbohD/alr1-1 transgenic lines were treated with or without 50 µM AlCl3 in 0.5 mM CaCl2 solution (pH 4.5) for 2, 5, 10 and 30 min, respectively. The seedlings were harvested and lysed in 1 mL of RIPA buffer. The FLAG-RbohD proteins were immune-precipitated by anti-Flag magnetic beads (Med Chem Express, HY-K0207), and the phosphorylation was detected by immunoblot with pS39 antibodies (1:5000). The total proteins were detected with α-Flag antibody (1:5000; ABclonal).
BIAM labeling assay
For in vitro BIAM labeling, 30 µL of recombinant proteins were treated with 0, 100 and 500 µM H2O2 and incubated in labeling buffer (50 mM MES, 100 mM NaCl, 1% Triton X-100, 100 µM BIAM, pH 6.5) at room temperature for 30 min. The proteins were precipitated, re-dissolved, and separated by SDS-PAGE as previously described.34 BIAM-labeled proteins were detected by immunoblot with α-biotin HRP-Linked antibody (1:5000; Cell signaling, 7075 S). The total His-STOP1 and His-RAE1 proteins were detected with α-His antibody (1:5000; ABclonal).
For in vivo BIAM labeling, 1-week-old transgenic Arabidopsis FLAG-RAE1/WT- and FLAG-RAE1/alr1-1 grown on 1/2 MS media were treated with or without 75 µM AlCl3 in 0.5 mM CaCl2 solution (pH 4.5) for 0.5 and 1 h, respectively. The seedlings were harvested and lysed in 1 mL of RIPA buffer. The FLAG-RAE1 proteins were immune-precipitated by anti-Flag magnetic beads at 4 °C. The magnetic beads were washed four times, and then incubated with BIAM-labeling buffer at room temperature in the dark for 30 min with constant shaking. BIAM-labeled proteins were finally detected by immunoblot with α-biotin HRP-Linked antibody (1:5000; Cell signaling, 7075 S). The total proteins were detected with α-Flag antibody (1:5000; ABclonal).
Mass spectrometric analysis for BIAM modification
The purified His-RAE1 protein labeled with BIAM was digested in gel by trypsin, and then analyzed by LC-MS/MS analysis as previously described.59 The molecular weight for BIAM modification on Cys is ~326.14 kD.
Co-IP assay
For Co-IP assays, 3 mL of protoplasts from FLAG-ALR1/WT were transfected with 200 µg pUC35S-YFP-BAK1 or pUC35S-YFP-empty. After 12 h incubation at room temperature, 50 µM of AlCl3 was added to the protoplasts for 10 min. The protoplasts were then harvested and lysed in 500 µL IP buffer (20 mM Tris-HCl, pH 7.4, 1 mM EDTA, 100 mM NaCl, 1% NP-40, 0.25% sodium deoxycholate, 1× protease inhibitor cocktail). After centrifugation at 13,000× g for 10 min, 40 µL of supernatants were kept as input controls, whilst the rest was incubated with 25 µL of GFP-Trap (ChromoTek) for 2 h. After the beads were washed five times with TBS buffer, the proteins were eluted by adding 60 µL 2× SDS sampling buffer at 95 °C for 5 min. Then the eluted proteins and input controls were further detected with α-GFP (1:5000; ABclonal) and α-FLAG (1:5000; ABclonal) antibodies.
MST assay
The MST assay was performed as previously described with minor modification.60 The affinity of the purified ALR1CD or its mutant (ALR1CD4C-A) with AlCl3 or other metal ions (LaCl3, CdCl2, CeCl3, InCl3, GaCl3, FeCl3, MnCl2, and CaCl2) was measured using the Monolith NT.115 (Nanotemper Technologies). Proteins were first fluorescently labeled using the Monolith Protein Labeling Kit RED-NHS 2nd Generation (Nanotemper Technologies, MO-L011) according to the manufacturer’s protocol, and the labeled protein used for each assay was about 100 nM. A solution of unlabeled metal ions was diluted for appropriate serial concentration gradient. The samples were loaded into MST standard capillaries (Nanotemper Technologies, MO-K022). Measurements were performed in buffer containing 50 mM HEPES, pH 7.4, 150 mM NaCl, 1 mM MgCl2, 2 mM DTT, and 0.05% Tween-20, by using medium MST power and 20% LED power. Data were fitted in Kd model using MO.Affinity Analysis v2.2.4, and were finally displayed in ΔFnorm normalization.
CD spectrum
The CD spectrum was obtained using a J-1500 spectropolarimeter (JASCO). The purified protein ALR1CD and ALR1CD4C-A were dissolved in phosphate buffer (pH 7.6) containing 10 mM Na2HPO4 and 1.8 mM KH2PO4. Protein concentration was adjusted to 0.2 mg/mL. Measurements were performed at wavelengths ranging from 190 to 260 nm. The cell length was 1 mm, bandwidth was 1 nm, scanning speed was 100 nm/min, response time was 1 s and data pitch was 1 nm.
Statistical analysis
Independent experiments were performed at least three times, unless indicated otherwise. All data were statistically analyzed using GraphPad Prism 8. The unpaired t-test was conducted when two samples were compared. For comparison of multi samples, data were analyzed by two-way ANOVA when two factors were introduced.
Data availability
The RNA-seq data can be accessed at NCBI with accession number PRJNA913658. All other data needed to evaluate the conclusions in the manuscript are present in the manuscript or the Supplementary information.
References
McBride,, M. B. Environmental Chemistry of Soils. (Oxford University Press, New York, 1994).
Kochian, L. V. Cellular mechanisms of aluminum toxicity and resistance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 46, 237–260 (1995).
Ma, J. F., Ryan, P. R. & Delhaize, E. Aluminum tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 6, 273–278 (2001).
von Uexküll, H. R. & Mutert, E. Global extent, development and economic impact of acid soils. In: Date, R. A., Grundon, N. J., Raymet, G. E. & Probert, M. E., (eds.) Plant–soil Interactions at low pH: principles and management. 5–19 (Dordrecht, The Netherlands: Kluwer Academic Publishers, 1995).
Kochian, L. V., Piñeros, M. A., Liu, J. P. & Magalhaes, J. V. Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 66, 571–598 (2015).
Godbold, D. L., Fritz, E. & Hüttermann, A. Aluminum toxicity and forest decline. Proc. Natl. Acad. Sci. USA 85, 3888–3892 (1988).
Ma, J. F., Zheng, S. J., Hiradate, S. & Matsumoto, H. Detoxifying aluminum with buckwheat. Nature 390, 569–570 (1997).
Magalhaes, J. V. et al. A gene in the multidrug and toxic compound extrusion (MATE) family confers aluminum tolerance in sorghum. Nat. Genet. 39, 1156–1161 (2007).
de la Fuente, J. M., Ramírez-Rodríguez, V., Cabrera-Ponce, J. L. & Herrera-Estrella, L. Aluminum tolerance in transgenic plants by alteration of citrate synthesis. Science 276, 1566–1568 (1997).
Delhaize, E. et al. Engineering high-level aluminum tolerance in barley with the ALMT1 gene. Proc. Natl. Acad. Sci. USA 101, 15249–15254 (2004).
Hoekenga, O. A. et al. AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 103, 9738–9743 (2006).
Liu, J. P., Magalhaes, J. V. & Kochian, L. V. Aluminum-activated citrate and malate transporters from the MATE and ALMT families function independently to confer Arabidopsis aluminum tolerance. Plant J. 57, 389–399 (2009).
Iuchi, S. et al. Zinc finger protein, STOP1, is critical for proton tolerance in Arabidopsis and co-regulates a key gene in aluminum tolerance. Proc. Natl. Acad. Sci. USA 104, 9900–9905 (2007).
Yamaji, N. et al. A zinc finger transcription factor ART1 regulates multiple genes implicated in aluminum tolerance in rice. Plant Cell 21, 3339–3349 (2009).
Ohyama, Y. et al. Characterization of AtSTOP1 orthologous genes in tobacco and other plant species. Plant Physiol. 162, 1937–1946 (2013).
Zhang, Y. et al. F-box protein RAE1 regulates the stability of the aluminum-resistance transcription factor STOP1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 116, 319–327 (2019).
Fang, Q. et al. Regulation of aluminum-resistance in Arabidopsis involves the SUMOylation of the zinc finger transcription factor STOP1. Plant Cell 32, 3921–3938 (2020).
Zhou, F. et al. The MEKK1-MKK1/2-MPK4 cascade phosphorylates and stabilizes STOP1 to confer aluminum resistance in Arabidopsis. Mol. Plant 16, 337–353 (2023).
Kinoshita, T. et al. Binding of brassinosteroids to the extracellular domain of plant receptor kinase BRI1. Nature 433, 167–171 (2005).
Laohavisit, A. et al. Quinone perception in plants via leucine-rich-repeat receptor-like kinases. Nature 587, 92–97 (2020).
Wu, F. et al. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis. Nature 578, 577–581 (2020).
Matsuzaki, Y., Ogawa-Ohnishi, M., Mori, A. & Matsubayashi, Y. Secreted peptide signals required for maintenance of root stem cell niche in Arabidopsis. Science 329, 1065–1067 (2010).
Takahashi, F. et al. A small peptide modulates stomatal control via abscisic acid in long-distance signaling. Nature 556, 235–238 (2018).
Shiu, S. H. & Bleecker, A. B. Plant receptor-like kinase gene family: diversity, function, and signaling. Sci. STKE 113, re22 (2001).
Matsubayashi, Y., Ogawa, M., Kihara, H., Niwa, M. & Sakagami, Y. Disruption and overexpression of Arabidopsis phytosulfokine receptor gene affects cellular longevity and potential for growth. Plant Physiol. 142, 45–53 (2006).
Mosher, S. et al. The tyrosine-sulfated peptide receptors PSKR1 and PSY1R modify the immunity of Arabidopsis to biotrophic and necrotrophic pathogens in an antagonistic manner. Plant J. 73, 469–482 (2013).
Ladwig, F. et al. Phytosulfokine regulates growth in Arabidopsis through a response module at the plasma membrane that includes CYCLIC NUCLEOTIDE-GATED CHANNEL17, H+-ATPase, and BAK1. Plant Cell 27, 1718–1729 (2015).
Wang, J. et al. Allosteric receptor activation by the plant peptide hormone phytosulfokine. Nature 525, 265–268 (2015).
Balzergue, C. et al. Low phosphate activates STOP1-ALMT1 to rapidly inhibit root cell elongation. Nat. Commun. 8, 15300 (2017).
Amano, Y., Tsubouchi, H., Shinohara, H., Ogawa, M. & Matsubayashi, Y. Tyrosine-sulfated glycopeptide involved in cellular proliferation and expansion in Arabidopsis. Proc. Natl. Acad. Sci. USA 104, 18333–18338 (2007).
Torres, M. A., Dangl, J. L. & Jones, J. D. G. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 99, 523–528 (2002).
Li, L. et al. The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity. Cell Host Microbe 15, 329–338 (2014).
Møller, I. M., Jensen, P. E. & Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 58, 459–481 (2017).
Kim, J. R. et al. Identification of proteins containing cysteine residues that are sensitive to oxidation by hydrogen peroxide at neutral pH. Anal. Biochem. 283, 214–221 (2000).
Tainer, J. A., Roberts, V. A. & Getzoff, E. D. Metal binding sites in proteins. Curr. Opin. Biotech. 2, 582–591 (1991).
Ding, Y. et al. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 173, 1454–1467 (2018).
Jung, J. H. et al. A prion-like domain in ELF3 functions as a thermosensor in Arabidopsis. Nature 585, 256–260 (2020).
Dorone, Y. et al. A prion-like protein regulator of seed germination undergoes hydration-dependent phase separation. Cell 184, 4284–4298 (2021).
Chandrashekar, J. et al. The cells and peripheral representation of sodium taste in mice. Nature 464, 297–301 (2010).
Ho, C. H., Lin, S. H., Hu, H. C. & Tsay, Y. F. CHL1 functions as nitrate sensor in plants. Cell 138, 1184–1194 (2009).
Jiang, Z. et al. Plant cell-surface GIPC sphingolipids sense salt to trigger Ca2+ influx. Nature 572, 341–346 (2019).
Kobayashi, T. et al. Iron-binding haemerythrin RING ubiquitin ligases regulate plant iron responses and accumulation. Nat. Commun. 4, 2792 (2013).
Lilay, G. H. et al. Arabidopsis bZIP19 and bZIP23 act as zinc sensors to control plant zinc status. Nat. Plants 7, 137–143 (2021).
Liu, K. H. et al. NIN-like protein 7 transcription factor is a plant nitrate sensor. Science 377, 1419–1425 (2022).
Hohmann, U., Lau, K. & Hothorn, M. The structural basis of ligand perception and signal activation by receptor kinases. Annu. Rev. Plant Biol. 68, 109–137 (2017).
Wang, J. et al. Structural basis of ALMT1-mediated aluminum resistance in Arabidopsis. Cell Res. 32, 89–98 (2022).
Matsubayashi, Y., Ogawa, M., Morita, A.& Sakagami, Y. An LRR receptor kinase involved in perception of a peptide plant hormone, phytosulfokine. Science 296, 1470–1472 (2002).
Mittler, R., Zandalinas, S. I., Fichman, Y. & Breusegem, F. V. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 23, 663–679 (2022).
Helariutta, Y. et al. The SHORT-ROOT-gene controls radial patterning of the Arabidopsis root through radial signaling. Cell 101, 555–567 (2000).
Ou, Y. et al. RGF1 INSENSITIVE 1 to 5, a group of LRR receptor-like kinases, are essential for the perception of root meristem growth factor 1 in Arabidopsis thaliana. Cell Res. 26, 686–698 (2016).
Sagi, M. et al. Superoxide Production by Plant Homologues of the gp91phox NADPH Oxidase. Modulation of Activity by Calcium and by Tobacco Mosaic Virus Infection. Plant Physiol. 126, 1281–1290 (2001).
Ding, Z. J., Yan, J. Y., Xu, X. Y., Li, G. X. & Zheng, S. J. WRKY46 functions as a transcriptional repressor of ALMT1, regulating aluminum-induced malate secretion in Arabidopsis. Plant J. 76, 823–835 (2013).
Li, C. X. et al. A WRKY transcription factor confers aluminum tolerance via regulation of cell wall modifying genes. J. Integr. Plant Biol. 62, 1176–1192 (2020).
Delhaize, E., Ryan, P. R. & Randall, P. J. Aluminum tolerance in wheat (Triticum aestivum L.) (II. Aluminum-stimulated excretion of malic acid from root apices). Plant Physiol. 103, 695–702 (1993).
Schmittgen, T. D. & Livak, K. J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 3, 1101–1108 (2008).
Stagljar, I., Korostensky, C., Johnsson, N. & te Heesen, S. A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo. Proc. Natl. Acad. Sci. USA 95, 5187–5192 (1998).
Yoo, S. D., Cho, Y. H. & Sheen, J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc. 2, 1565–1672 (2007).
Allen, J. J. et al. A semisynthetic epitope for kinase substrates. Nat. Methods 4, 511–516 (2007).
Tian, Y. et al. Hydrogen peroxide positively regulates brassinosteroid signaling through oxidation of the BRASSINAZOLE-RESISTANT1 transcription factor. Nat. Commun. 9, 1063 (2018).
Jerabek-Willemsen, M. et al. MicroScale thermophoresis: interaction analysis and beyond. J. Mol. Struc. 1077, 101–113 (2014).
Acknowledgements
We thank Prof. Heven Sze and Prof. William J. Snell (University of Maryland), Dr. Peter Ryan (CSIRO, Australia) for critical reading and commenting the manuscript, Prof. Thierry Desnos (CEA Cadarache, France) for providing the STOP1p:GFP-STOP1/stop1 line, Prof. Weicai Yang (Chinese Academy of Sciences, Beijing) and Prof. Jia Li (Guangzhou University, Guangzhou) for providing serk1, serk2, serk3 (bak1-4) and serk1+/– serk2–/– serk3–/– mutants, Prof. Chaofeng Huang (Shanghai Center for Plant Stress Biology, Shanghai) for providing the rae1-1 loss-of-function mutant, Prof. Dong Liu (Peking University, Beijing) for the split-LUC vectors, Prof. Yan Xiong and Dr. Yanlin Liu (Fujian Agriculture and Forestry University) for technical assistance with mesophyll protoplast experiments, and Dr. Shelong Zhang (College of Life Sciences, ZJU) for their technical assistance with MST and confocal microscopic analyses. This work was supported by the National Natural Science Foundation of China (31730006), the Ministry of Science and Technology of China (2015CB942903), the National Key R&D Program of China (2022YFA1303402), the National Natural Science Foundation of China (31970272), Guangdong Laboratory for Lingnang Modern Agriculture (NT2021010), the “111” project from the Ministry of Education Bureau of Foreign Experts of China (B14027), ZJU Tang Scholar Foundation, and the Fundamental Research Funds for the Central Universities.
Author information
Authors and Affiliations
Contributions
S.J.Z. proposed the research; Z.J.D. and S.J.Z. designed the research; Z.J.D. and C.X. performed most experiments; J.Y.Y., Y.X.W., M.Q.C., J.J.Y., Y.N.W., G.X.L. and C.X.L. performed additional experiments; Y.Z.S. and J.M.X. conducted elemental analysis; J.X.W. and Y.R.W. were involved in antibody analysis and plant material generation; Z.J.D., C.X. and S.J.Z. analyzed data; M.B. assisted transcriptomic analysis; G.X.L., N.P.H., C.Z.M., J.T.G., and J.M.Z. discussed part of the experimental design; Z.J.D, S.J.Z, C.X., N.P.H., and J.M.Z. contributed to writing the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ding, Z.J., Xu, C., Yan, J.Y. et al. The LRR receptor-like kinase ALR1 is a plant aluminum ion sensor. Cell Res 34, 281–294 (2024). https://doi.org/10.1038/s41422-023-00915-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41422-023-00915-y
This article is cited by
-
Sensing the toxic aluminum cations in acidic soils
Cell Research (2024)
