
Abstract
The nucleus accumbens (NAc) is considered a hub of reward processing and a growing body of evidence has suggested its crucial role in the pathophysiology of major depressive disorder (MDD). However, inconsistent results have been reported by studies on reward network-focused resting-state functional MRI (rs-fMRI). In this study, we examined functional alterations of the NAc-based reward circuits in patients with MDD via meta- and mega-analysis. First, we performed a coordinated-based meta-analysis with a new SDM-PSI method for all up-to-date rs-fMRI studies that focused on the reward circuits of patients with MDD. Then, we tested the meta-analysis results in the REST-meta-MDD database which provided anonymous rs-fMRI data from 186 recurrent MDDs and 465 healthy controls. Decreased functional connectivity (FC) within the reward system in patients with recurrent MDD was the most robust finding in this study. We also found disrupted NAc FCs in the DMN in patients with recurrent MDD compared with healthy controls. Specifically, the combination of disrupted NAc FCs within the reward network could discriminate patients with recurrent MDD from healthy controls with an optimal accuracy of 74.7%. This study confirmed the critical role of decreased FC in the reward network in the neuropathology of MDD. Disrupted inter-network connectivity between the reward network and DMN may also have contributed to the neural mechanisms of MDD. These abnormalities have potential to serve as brain-based biomarkers for individual diagnosis to differentiate patients with recurrent MDD from healthy controls.
Similar content being viewed by others
Introduction
Major depressive disorder (MDD) with heterogeneous symptoms immensely impairs patients’ health and function, destroys patients’ occupations and life, and mounts a burden borne by the family and society [1]. This disorder is characterized by a high recurrent rate and a chronic deteriorating disease course. As high as 83.3% (at least 33.5%) of patients with MDD may experience recurrence within 6 months [2]. Each recurrence of MDD especially increases the risk of becoming chronic. Up to 70% of patients with MDD may suffer recurrences throughout their lifetime after the second major depressive episode and by 90% after the third or more recurrence [3]. Over the past decades, with the development of precision medicine, more and more studies have focused on searching for objective biomarkers and exploring subtle classifications to guide clinical decision-making and facilitate individualized therapy.
Magnetic resonance imaging (MRI) is one of the noninvasive neuroimaging techniques used extensively to elucidate the neural basis of psychiatric disorders. This technique may be a critical step toward differential diagnoses and treatment prediction. Previous behavioral and MRI studies have implicated altered reward-related processes (e.g., reward anticipation, acceptance, and motivation) in patients with MDD [4,5,6]. Brain areas such as the putamen, ventral tegmental area (VTA), ventral striatum, anterior cingulate cortex (ACC), and medial prefrontal cortex (mPFC) are the components of complicated reward network [7]. Notably, converging evidence has suggested that structural and functional disruptions of the nucleus accumbens (NAc, a part of the ventral striatum) might be a crucial culprit of reward-related abnormalities. The NAc is a key component of dopamine-rich mesocorticolimbic pathways. This region is extensively connected with other brain regions and receives complex signal inputs from various neurochemistry systems including glutamatergic, dopaminergic, serotoninergic, and histaminergic projections [8,9,10]. For instance, the NAc volumes were smaller in patients with life-time MDD than in healthy controls [11]. During a monetary incentive delay task, individuals with MDD displayed significantly weaker activations to gains in the left NAc and bilateral caudate regions [12]. Generally, decreased activation within the ventral or dorsal striatum, coupled with increased or decreased activation in other areas (such as the mPFC and subgeneual ACC) was the most consistent finding in adolescents with MDD in the reward-related task studies using functional MRI (fMRI) [13,14,15]. Similarly, aberrant functional connectivities (FCs) between the NAc and other regions within the reward network, including the ACC, orbital frontal cortex (OFC) and mPFC, have been reported in adult patients with MDD in the resting-state fMRI (rs-fMRI) studies. These disruptions have been associated with depressive severity, cognitive deficit, and treatment response to repetitive transcranial magnetic stimulation [16,17,18,19]. Intriguingly, some brain regions such as the mPFC were also involved in the default mode network (DMN), a network which plays a critical role in self-referential and internal goal-oriented processes [20, 21]. The balanced activity between the reward network and other networks, such as the cognitive control network and DMN, was considered important for self-regulation, emotional regulation and reward motivation, and decision-making [22,23,24]. Gong et al. [25] reported decreased FCs between the NAc and areas within the reward network (bilateral caudate, hippocampus, left pallidum, etc.) and key hubs of the DMN (inferior parietal lobe and dorsal mPFC) and cognitive network (dorsolateral PFC). Furthermore, decreased inter-network FC and diminished controlling of the cognitive network on the reward network were documented in this study, and the imbalance between these networks could predict the severity of anhedonia in patients with MDD.
However, inconsistent results have been reported in these reward network-focused rs-fMRI studies. For example, both increased and decreased FCs have been observed between the NAc and the precuneus and mPFC [16, 18, 25,26,27]. Diverse demographic and clinical profiles of the subjects, analytical methods, and limited statistical power may have contributed to the inconsistencies. Therefore, in the current study, we examined functional alterations in the NAc-based reward circuits in patients with MDD via meta-analysis and mega-analysis. First, we performed a neurofunctional meta-analysis of all up-to-date rs-fMRI studies that focused on the reward circuits of patients with MDD. Then, we tested the meta-analysis results in a large MDD rs-fMRI database, that is, REST-meta-MDD (rfmri.org/REST-meta-MDD). This project contains anonymous rs-fMRI data processed using a standard pipeline built-in Data Processing & Analysis for Brain Imaging/Data Processing Assistant for Resting-State fMRI (DPABI/DPARSF) of 1,300 patients with MDD and 1,128 healthy controls. With the use of these data, project initiators Yan et al. found reduced DMN FC only in patients with recurrent MDD [28]. In the present investigation, we predicted decreased NAc FC in the reward network and compromised relationships between the NAc-based reward network and other networks (e.g., DMN), and mega-analysis would replicate the meta-analysis findings. In addition, we tested whether the altered NAc FC would serve as a potential image feature to distinguish patients with MDD from healthy controls.
Materials and methods
Meta-analysis
Search strategy, selection criteria, and data extraction
The meta-analysis was prepared compliant with Preferred Reporting Items for Systematic Reviews and Meta-analyses guidelines. A literature search of the PubMed and Web of Science database was conducted in September 2021. The following keywords and their combinations were used in [Title/Abstract]: resting-state functional magnetic resonance imaging/rsfMRI, major depressive disorder/MDD/depression/unipolar depression/depressive disorders, and reward network/nucleus accumbens/NAc/ventral striatum/VS. The reference lists of previous review articles were also manually searched.
All original rs-fMRI studies that compared patients with MDD with healthy controls and investigated NAc/VS FC maps were included. Meeting abstracts, study protocols, reviews, and animal studies were excluded. Studies were also excluded if they used different thresholds in different brain areas (e.g., region of interest) or recruited subjects with other major psychiatric or neurological illness.
The following variables were extracted from included studies: sample size, age, sex, education level of patients and controls, depressive severity, type of analysis, statistical thresholds, effect sizes, and if available, the Montreal Neurological Institute space (MNI) or Talairach space coordinates of brain areas with significant group differences.
Statistical analysis
We used a new method, SDM-PSI, to conduct this coordinated-based meta-analysis. This method was developed by Radua et al., and they implemented the Permutation of Subject Images (PSI) algorithm to an existing Anisotropic Effect-Size Seed-based d Mapping (AES-SDM) method [29]. Based on the AES-SDM method [30, 31], a set of reported peak coordinates and t-values were converted into Hedge’s g effect size using standard formulas. This process imputes the slightly lower effect sizes of surrounding voxels than those of the peaks, and until the voxels are far enough from any peak it imputes the effect size as null. However, non-negligible biases would be yielded in the progressive estimation of the effect size of voxels farther from the peaks. To address this limitation, the SDM-PSI method uses AES-SDM Gaussian kernels to calculate the effect-size bounds, maximum-likelihood estimation to estimate parameters, and multiple imputation techniques for each voxel to cover the uncertainty linked to single imputation with a range of effect sizes a voxel may have [29]. Importantly, the MetaNSUE (nonstatistically significant unreported effects [NSUE]) method is adapted to avoid biases introduced by excluding studies with non-significant results and unknown statistics [32, 33].
Age and years of education were included as covariates in the analyses. Given the intra- and inter-study heterogeneities, random-effect models were used and I2 statistic was applied to quantify heterogeneity (I2 ≥ 50% indicates substantial heterogeneity). The threshold was set at p < 0.001 uncorrected and a cluster extent of ten voxels. According to previous studies [29], this particular set of threshold was a conservative recommendation for cases where the t-values were not reported by studies and simultaneously has a remarkable sensitivity and controls the empirical family-wise error rate below 5%. Funnel plots were used to detect whether results might have been biased by small studies. In addition, meta-regression by the mean MDD symptom severity and the percentage of male individuals in the patient group were complemented to the main analysis. All analyses were performed using the SDM-PSI version 6.21 (https://www.sdmproject.com/).
We also conducted a supplementary analysis for the studies only focused on the NAc rather than VS, given possible inconsistency introduced by a broader ROI selection.
Mega-analysis
Subjects
From the data of 2428 subjects provided by the REST-meta-MDD consortium, we selected the data of 186 subjects with recurrent MDDs and 465 matched healthy controls from eight sites across China (the majority of patients included in the meta-analysis were recurrent MDDs. Thus, we focused on recurrent MDDs). The sample were selected as shown in Fig. S1. All individuals aged 18–65 years with complete information (i.e., age, sex, and educational level) were included. Subjects were excluded if they (1) had poor imaging quality or poor spatial normalization (checked by visual inspection), (2) were in remission stage (the score of the 17-item Hamilton depression rating scale (HAMD) ≤7), or (3) had excessive head motion (mean framewise displacement, FD >0.2 mm). Among the 803 remaining patients with MDD, 186 were patients with recurrent MDDs. Data on the duration of illness and 17-item HAMD scores were available for 165 and 143 patients with recurrent MDDs, respectively. Medication information was provided by 126 patients with recurrent MDD, among which 77 patients were taking antidepressants when receiving MRI scans.
Data acquisition, preprocessing, and FC analysis
MRI scans and data preprocessing were performed independently at each site. The scanning parameters of each site and preprocessing details are shown in the Supplementary Materials. As previously described in refs. [28, 34], all preprocessing steps were completed with the DPARSF software, and analysts from each site were able to use it skillfully.
After preprocessing, time series for the Harvard-Oxford Atlas were extracted. We selected bilateral NAc as seed regions and calculated FC (Fisher’s z-transformed Pearson’s correlation coefficients) between the seeds and other brain areas.
Statistical analysis
Bilateral NAc FC maps of patients with recurrent MDD and controls were compared by linear mixed models (LMM) to control for potential study site-related effects, in which the group, age, sex, years of education, and mean FD were included as independent variables: y~1 + group + age + sex + years of education + FD + (1 | site) + (group | site) [28]. Then the yielded t and df values were used to calculate effect size Cohen’s d = (t(n1 + n2)) ⁄ (√df)√n1n2) [35]. To further test the potential effect of the depressive severity on the brain regions with significant group differences, we replaced “group” in the LMM model with HAMD total score or score of core depression subtype (item 1 and 7) [36]. The Pearson’s correlation coefficient r = t / √(n – 2 + t2) [37]. The statistical significance threshold was set at p < 0.05 with a false discovery rate (FDR) correction.
Machine learning analysis
We used brain areas which had significantly disrupted FC with NAc as potential features and examined their ability to discriminate patients with recurrent MDDs from healthy controls by using a support vector machine (SVM) in Matlab. SVM could handle data in a high-dimensional space and create a hyperplane that could best classify a new target into predefined categories. A Grid search method was used to obtain optimal values of hyperparameters (such as parameter C and Gamma) of a model. A fivefold cross-validation method was employed to assess the generalizability of classifier models. We trained those models with a Gaussian kernel in 80% of participants and evaluated the models’ performance on the left-out data in the “testing phase” for each fold. This method is good at handling high-dimensional data with optimal boundaries constructed and misclassification error minimized and is less likely to overfitting of the data [38, 39]. Finally, accuracy was calculated based on the results of cross-validation:
where TP and TN represent the number of patients and controls correctly predicted, respectively; FP represents the number of controls classified as patients and FN represents the number of patients classified as controls.
Results
Meta-analysis
Characteristics of eligible studies
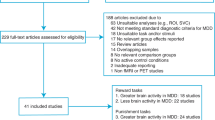
Twelve studies involving 1068 subjects (593 patients and 475 healthy controls) were included in the meta-analysis. Figure S2 shows the flow diagram of the identified, included, and excluded studies. The characteristics of the included studies are shown in the Supplementary Materials (Table S2). The majority of patients recruited in these studies were patients with recurrent MDD and aged 18–60 years old. Two studies have focused on adolescent (12–19 years) and older adults (>60 years) patients with MDD [18, 40]. Nine of the studies selected bilateral NAc as seeds, and the other three selected bilateral inferior VS as seeds. Symptomatic severity was not evaluated by HAMD in two studies, so they were not included in the meta-regression analysis [18, 41].
Group differences and meta-regression analyses
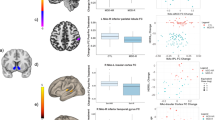
As shown in Fig. 1, compared with healthy controls, patients with recurrent MDD had decreased FC between the NAc and left ventromedial prefrontal area (peak MNI = −28, 36, −14, z = −4.089, p < 0.001, 59 voxels). No increased FCs were reported. Slight but not significant heterogeneity was found in this area (I2 = 0.95%, p = 0.968). No obvious publication bias was observed in the funnel plot (Supplementary Materials and Fig. S3). When we adopted a relatively lenient threshold at p < 0.01, significant results included the left ventromedial prefrontal area (peak MNI = −28, 36, −14, z = −4.089, p < 0.001, 205 voxels) and right ACC (peak MNI = 4, 36, −6, z = −2.519, p = 0.006, 12 voxels).

Compared with healthy controls, patients with recurrent MDD exhibited decreased NAc functional connectivity in the left ventromedial prefrontal cortex.
The meta-regression analysis did not find any significant effects of depressive severity or sex ratio on NAc FC maps.
Our supplementary analysis for studies only focused on the NAc showed no significantly positive results at a threshold of p < 0.001(uncorrected). When threshold was set at p < 0.01, decreased FC between the NAc and left ventromedial prefrontal area (peak MNI = −28, 36, −14, z = −3.219, p < 0.001, 115 voxels) was observed.
Mega-analysis
Group comparisons
The demographic and clinical characteristics of the included subjects are shown in Table 1. No significant differences were reported between patients with recurrent MDD and healthy controls in age and sex ratio. Compared with healthy controls, patients with recurrent MDD exhibited lower education levels.
As shown in Table 2 and Fig. 2, compared with healthy controls, patients with recurrent MDD mainly exhibited generally decreased bilateral NAc FC in the reward network, including the left hippocampus/parahippocampal gyrus (left NAc seed: t = −3.903, p < 0.001, d = −0.340; right NAc seed: t = −3.705, p < 0.001, d = −0.324) and left VTA (right NAc seed: t = −3.342, p = 0.028, d = −0.292). A region within the DMN (right lateral temporal cortex, t = −3.347, p = 0.037, d = −0.292) and a region involved in visual processing (left fusiform gyrus, t = −4.507, p < 0.001, d = −0.393) were also survived multiple comparison correction (The uncorrected results were showed in Supplementary Materials Table S3). No significant associations were found between bilateral NAc FC maps and HAMD scores or subscores in the patients.

A The significant between-group differences in the NAc functional connectivity. B The box figures show the distribution of significantly disrupted bilateral NAc functional connectivity in the group comparison. Fusi fusiform gyrus, HC healthy control, L left, NAc nucleus accumbens, PHa parahippocampal gyrus, R right, Re recurrent major depressive disorder, ITG inferior temporal gyrus, VTA ventral tegmental area.
Distinguishing patients with recurrent MDD from healthy controls
Significantly abnormal bilateral NAc FCs returned by the former step were used as potential features for subsequent SVM analysis. As shown in Fig. 3, the combination of all significant NAc FCs as features could discriminate patients with recurrent MDD from healthy controls with an accuracy of 74.5%; when using significant NAc FCs within the reward network as features, the discriminative accuracy was 74.7%. The specificities of the two strategies were high (100% and 98.7%, respectively), whereas neither of the sensitivities were more than 20%.

A The accuracy, sensitivity, and specificity of the classifications of patients with recurrent MDD versus HCs using all significant NAc FCs as features or using significant NAc FCs within the reward network as features. B Parameter selection result for training set when using (1) all significant NAc FCs as features and (2) using significant NAc FCs within the reward network as features. FC functional connectivity, HC healthy controls, MDD major depressive disorder, NAc nucleus accumbens, SVM support vector machine.
Discussion
Using a meta-analysis and a large MDD sample for mega-analysis, we found decreased FCs within the NAc-based reward system in patients with recurrent MDD compared with healthy controls as predicted. In addition, reduced NAc-DMN FCs were prominent. No significantly increased FCs between the NAc and other brain regions were reported. Specifically, the combinations of the disrupted NAc FCs within the reward network could discriminate patients with recurrent MDD from healthy controls with an optimal accuracy of 74.7%. Though the sensitivity was not satisfactory, the high specificity could ensure the correct identification of healthy individuals.
This study has two main novel aspects. First, we used a new coordinate-based meta-analysis method, namely, SDM-PSI, to statistically summarize up-to-date voxel-based rs-fMRI studies that focused on the NAc and reward network in patients with MDD. This method benefits from the control of family-wise error rate via subject-based permutation test and the control of bias introduced by exclusion of studies with non-significant results and unknown statistics via multiple imputations algorithms-based MetaNSUE method [29]. Second, we used a large rs-fMRI database of MDD from 25 sites in China to cross-validate the findings of the meta-analysis. These imaging data were all preprocessed in the same protocol with the DPARSF software and thus eliminating analytic heterogeneity. Furthermore, we employed SVM to test the potential of the disrupted NAc-based FC map as a biomarker of MDD in this database. The relatively high accuracy of our SVM analyses indicated the important roles of the reward network and its inter-network connectivity with the DMN in the pathophysiology of MDD, and may have promising clinical implications to provide helpful scaffolding in promoting objective diagnostic tools for MDD and developing individualized treatment.
The most robust finding in this study was decreased FC within the reward system, which was observed both in the meta-analysis and mega-analysis. The ACC, mPFC, and hippocampus/parahippocampal gyrus are important hubs in the reward circuit and play critical roles in reward motivation and reappraisal, emotional regulation, and reasoning along with the cognitive control network [24, 42, 43]. Histopathological studies have reported neuron size reduction and glia loss in the ACC and mPFC [44,45,46]. Mounting evidence from the fMRI studies has implicated alterations of these regions in patients with different ages [47,48,49,50]. Chen et al. found altered variability in the dynamic FC between the inferior VS and mPFC in patients with MDD compared with healthy controls and patients with bipolar disorder II, which indicates an MDD-specific alteration [51]. Notably, a negative association between the NAc-subgenual ACC intrinsic FC and anhedonia severity has been reported in adolescent patients with MDD [18]. Anhedonia, which reflects deficits in pleasure feeling and reward processing, is a core symptom of MDD and even persists after treatment [52, 53]. A previous meta-analysis reported that decreased activation in the left ACC was associated with consummatory anhedonia in patients with MDD [54]. Similarly, Rzepa et al. observed blunted neural responses in the ACC and vmPFC during the consummatory phase of rewarding stimuli in young people at risk of depression [55].
It is well established that the hippocampus/parahippocampal gyrus and VTA have close connections with the NAc. The hippocampus receives and sends inputs to the NAc and plays critical and complex roles in the processing of reward valence [56]. The VTA is a heterogeneous brain region, which has dopaminergic and GABAergic projections to the NAc [56]. Previous task fMRI studies found decreased FC between the VTA and striatum in patients with MDD during reward outcome in monetary instrumental learning task compared to healthy individuals, which indicated reward-related deficit in MDD and the underlying disrupted VTA-NAc circuit [57]. Shi et al. reported increased resting-state FCs between the bilateral VTA and ventral striatum in MDD compared to healthy controls using an ROI-to-ROI analysis, but these results were not survived correction for multiple comparisons [58]. Using the same ROI-based method, Wang et al. observed reduced resting-state NAc FCs in the bilateral hippocampus in MDD [25]. Intriguingly, our meta-analysis for all up-to-date NAc-based whole-brain resting-state fMRI studies did not find any significant positive results in the subcortical areas within the reward network, which instead were the main findings in the mega-analysis. Methodology differences may contribute to the inconsistence. In addition to different data preprocessing and analysis operations and diverse multiple comparison correction methods, disparities in NAc definition may be one of the meaningful factors. According to the results of supplementary meta-analysis, the inclusion of ventral striatum in the meta-analysis did not seem to contribute to the heterogeneity, instead, the subregions of NAc are worth noting. The NAc has a core-like and a shell-like subdivision, and the two parts have connections with different regions associated with different reward-related functions [59, 60]. Given the coordinates of bilateral NAc in the Harvard-Oxford Atlas used in the mega-analysis and those used in the meta-analysis, findings from the meta-analysis were more likely to present an abnormal NAc shell-like subdivision-based FC profiles in patients with recurrent MDD. Albeit the disparities, the combination of meta-analysis and mega-analysis in this study highlighted the disrupted NAc-centered reward network in MDD.
Abnormal inter-network connectivity between the DMN and NAc (reward network) in the recurrent MDDs was another prominent result in this study. The ventral mPFC, the significantly positive result in the meta-analysis, was regarded as an important part of the DMN [61], and the DMN was considered to be engaged in reward processing. For instance, Olivo and colleagues reported that connectivity in the DMN (such as lateral temporal regions) was related to reward sensitivity [62]. Altered activation of the DMN was observed in effort avoidance behavior, which suggested that the DMN activity might be associated with reward processing that predicts effort selection [63]. The reward network-DMN FC deficits showed in our study indirectly corroborated previous studies. Using the precuneus/posterior cingulate gyrus as a seed, researchers found decreased DMN-bilateral caudate connectivity in early depression [64]. Hwang et al. [65] found increased FC between the DMN and ventral striatum in two cohorts of subthreshold depression and the value of DMN-ventral striatum FC was positively related to scores of depressive symptomatology (measured by the Center for Epidemiologic Studies Depression Scale). This result was regarded as compensation for the lowered reward function in patients with subthreshold depression. Taken together, our study and previous studies demonstrated disrupted FC between the reward network and DMN in MDD, and such abnormality may change at different stages of depression.
Unexpectedly, we found altered FC between the NAc and fusiform gyrus (a region adjoins hippocampus/parahippocampal gyrus and involves in visual pathways of recognition [66]) in the mega-analysis. On the one hand, some studies have indicated a possible role of the visual system in patients with MDD. Altered blood-oxygenation-level-dependent signals in the occipital lobes were found during the facial expression tasks [67], working memory tasks [68], and reward tasks [69]. Reduced nodal centralities in the occipital regions were documented in a study that investigated the topological organization of brain networks during the resting state in patients with MDD [70]. Processing emotion- or reward-related visual stimuli in the task and eyes-open status at rest may provide explanations for these findings [69, 71,72,73]. In addition, mixed medication treatments and different antidepressant responses in patient with recurrent MDD may affect the activation of the occipital cortex. For example, compared with non-responders, remitters had a more obvious decrease in activation in the occipital cortex following escitalopram treatment [74]. On the other hand, a significantly positive result seen in the fusiform gyrus in our mega-analysis may be a result of the extended effect of the hippocampus/parahippocampal gyrus. Thus, it is still unclear whether inter-network connectivities between the reward network and visual system play roles in MDD due to limited evidence.
Several limitations of this study should be acknowledged. First, most studies included in the meta-analysis and all patients recruited in the mega-analysis were from China. Thus, the results should be taken with caution when generalizing to other populations. Data from other ethnicity/culture (such as the UK biobank) should be analyzed and compared with our results. Similarly, combined data from other international data-sharing consortiums will help explore MDD subtypes (clinical-symptom based or neurophysiological based). Second, medication details are not available in our study and thus may confound the results. As previously mentioned, antidepressants and response disparities may have different effects on the brain function [74, 75]. Third, permutation tests used in the SDM-PSI analysis were not entirely free from bias. Images recreated from peak t values could not fully duplicate raw images [29]. Finally, results of associations between disrupted NAc-based reward network FC and HAMD scores were mixed in the meta-analysis and mega-analysis. Whether illness severity contributes to the abnormalities remains unknown. Future studies need to focus on specific symptoms (such as anhedonia) instead of using general rating scales.
In summary, by combining a meta-analysis and a large MDD sample for mega-analysis, we confirmed the critical role of NAc-based reward system in MDD. We identified decreased FC in the NAc-based reward network in patients with recurrent MDD. Disrupted inter-network connectivity between the reward network and DMN may also have contributed to the pathophysiological mechanisms of MDD. Moreover, a combination of abnormal NAc FCs in the reward network can serve as potential brain-based biomarkers for individual diagnosis to differentiate patients with recurrent MDD from healthy controls.
References
Smith K. Mental health: a world of depression. Nature. 2014. https://doi.org/10.1038/515180a.
Rush AJ, Trivedi MH, Wisniewski SR, Nierenberg AA, Stewart JW, Warden D, et al. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: a STAR*D report. Am J Psychiatry. 2006. https://doi.org/10.1176/ajp.2006.163.11.1905.
Monroe SM, Harkness KL. Recurrence in major depression: a conceptual analysis. Psychol Rev. 2011. https://doi.org/10.1037/a0025190.
Pizzagalli DA, Iosifescu D, Hallett LA, Ratner KG, Fava M. Reduced hedonic capacity in major depressive disorder: evidence from a probabilistic reward task. J Psychiatr Res. 2008. https://doi.org/10.1016/j.jpsychires.2008.03.001.
Rutledge RB, Moutoussis M, Smittenaar P, Zeidman P, Taylor T, Hrynkiewicz L, et al. Association of neural and emotional impacts of reward prediction errors with major depression. JAMA Psychiatry. 2017. https://doi.org/10.1001/jamapsychiatry.2017.1713.
Misaki M, Suzuki H, Savitz J, Drevets WC, Bodurka J. Individual variations in nucleus accumbens responses associated with major depressive disorder symptoms. Sci Rep. 2016. https://doi.org/10.1038/srep21227.
Haber SN, Knutson B. The reward circuit: linking primate anatomy and human imaging. Neuropsychopharmacology. 2010. https://doi.org/10.1038/npp.2009.129.
Abdallah CG, Jackowski A, Salas R, Gupta S, Sato JR, Mao X. et al. The nucleus accumbens and ketamine treatment in major depressive disorder. Neuropsychopharmacology. 2017. https://doi.org/10.1038/npp.2017.49.
Gipson CD, Kupchik YM, Kalivas PW. Rapid, transient synaptic plasticity in addiction. Neuropharmacology. 2014. https://doi.org/10.1016/j.neuropharm.2013.04.032.
Yager LM, Garcia AF, Wunsch AM, Ferguson SM. The ins and outs of the striatum: role in drug addiction. Neuroscience. 2015. https://doi.org/10.1016/j.neuroscience.2015.06.033.
Ancelin ML, Carriere I, Artero S, Maller J, Meslin C, Ritchie K. et al. Lifetime major depression and grey-matter volume. J Psychiatry Neurosci. 2019. https://doi.org/10.1503/jpn.180026.
Pizzagalli DA, Holmes AJ, Dillon DG, Goetz EL, Birk JL, Bogdan R, et al. Reduced caudate and nucleus accumbens response to rewards in unmedicated individuals with major depressive disorder. Am J Psychiatry. 2009. https://doi.org/10.1176/appi.ajp.2008.08081201.
Forbes EE, Hariri AR, Martin SL, Silk JS, Moyles DL, Fisher PM et al. Altered striatal activation predicting real-world positive affect in adolescent major depressive disorder. Am J Psychiatry. 2009. https://doi.org/10.1176/appi.ajp.2008.07081336.
Keedwell PA, Andrew C, Williams SC, Brammer MJ, Phillips ML The neural correlates of anhedonia in major depressive disorder. Biol Psychiatry. 2005 https://doi.org/10.1016/j.biopsych.2005.05.019.
Gaffrey MS, Luby JL, Repovs G, Belden AC, Botteron KN, Luking KR, et al. Subgenual cingulate connectivity in children with a history of preschool-depression. Neuroreport. 2010. https://doi.org/10.1097/WNR.0b013e32834127eb.
Gong L, Yin Y, He C, Ye Q, Bai F, Yuan Y et al. Disrupted reward circuits is associated with cognitive deficits and depression severity in major depressive disorder. J Psychiatr Res. 2017. https://doi.org/10.1016/j.jpsychires.2016.09.016.
Downar J, Geraci J, Salomons TV, Dunlop K, Wheeler S, McAndrews MP, et al. Anhedonia and reward-circuit connectivity distinguish nonresponders from responders to dorsomedial prefrontal repetitive transcranial magnetic stimulation in major depression. Biol Psychiatry. 2014. https://doi.org/10.1016/j.biopsych.2013.10.026.
Gabbay V, Ely BA, Li Q, Bangaru SD, Panzer AM, Alonso CM, et al. Striatum-based circuitry of adolescent depression and anhedonia. J Am Acad Child Adolesc Psychiatry. 2013. https://doi.org/10.1016/j.jaac.2013.04.003.
Janes AC, Zegel M, Ohashi K, Betts J, Molokotos E, Olson D, et al. Nicotine normalizes cortico-striatal connectivity in non-smoking individuals with major depressive disorder. Neuropsychopharmacology. 2018. https://doi.org/10.1038/s41386-018-0069-x.
Andrews-Hanna, JR. The brain’s default network and its adaptive role in internal mentation. Neuroscientist. 2012. https://doi.org/10.1177/1073858411403316.
Spreng, RN. The fallacy of a “task-negative” network. Front Psychol. 2012. https://doi.org/10.3389/fpsyg.2012.00145.
Heatherton TF, Wagner DD. Cognitive neuroscience of self-regulation failure. Trends Cogn Sci. 2011. https://doi.org/10.1016/j.tics.2010.12.005.
Heatherton, TF. Neuroscience of self and self-regulation. Annu Rev Psychol. 2011. https://doi.org/10.1146/annurev.psych.121208.131616.
Hu Y, Salmeron BJ, Gu H, Stein EA, Yang Y. Impaired functional connectivity within and between frontostriatal circuits and its association with compulsive drug use and trait impulsivity in cocaine addiction. JAMA Psychiatry. 2015. https://doi.org/10.1001/jamapsychiatry.2015.1.
Gong L, He C, Zhang H, Zhang H, Zhang Z, Xie, C. Disrupted reward and cognitive control networks contribute to anhedonia in depression. J Psychiatr Res. 2018. https://doi.org/10.1016/j.jpsychires.2018.05.010.
Meng Y, Li H, Wang J, Xu Y Wang B. Cognitive behavioral therapy for patients with mild to moderate depression: treatment effects and neural mechanisms. J Psychiatr Res. 2021. https://doi.org/10.1016/j.jpsychires.2021.02.001.
Hou Z, Gong L, Zhi M, Yin Y, Zhang Y, Xie C. et al. Distinctive pretreatment features of bilateral nucleus accumbens networks predict early response to antidepressants in major depressive disorder. Brain Imaging Behav. 2018. https://doi.org/10.1007/s11682-017-9773-0.
Yan CG, Chen X, Li L, Castellanos FX, Bai TJ, Bo QJ, et al. Reduced default mode network functional connectivity in patients with recurrent major depressive disorder. Proc Natl Acad Sci USA. 2019. https://doi.org/10.1073/pnas.1900390116.
Albajes-Eizagirre A, Solanes A, Vieta E, Radua J. Voxel-based meta-analysis via permutation of subject images (PSI): theory and implementation for SDM. Neuroimage. 2019 https://doi.org/10.1016/j.neuroimage.2018.10.077.
Radua J, Mataix-Cols D, Phillips ML, El-Hage W, Kronhaus DM, Cardoner N, et al. A new meta-analytic method for neuroimaging studies that combines reported peak coordinates and statistical parametric maps. Eur Psychiatry. 2012. https://doi.org/10.1016/j.eurpsy.2011.04.001.
Radua J, Rubia K, Canales-Rodriguez EJ, Pomarol-Clotet E, Fusar-Poli P, Mataix-Cols, D. Anisotropic kernels for coordinate-based meta-analyses of neuroimaging studies. Front Psychiatry. 2014. https://doi.org/10.3389/fpsyt.2014.00013.
Radua J, Schmidt A, Borgwardt S, Heinz, A, Schlagenhauf, F, McGuire, P et al. Ventral Striatal Activation During Reward Processing in Psychosis: A Neurofunctional Meta-Analysis. JAMA Psychiatry (2015). https://doi.org/10.1001/jamapsychiatry.2015.2196.
Albajes-Eizagirre, A, Solanes, A,Radua, J. Meta-analysis of non-statistically significant unreported effects. Stat Methods Med Res. 2019. https://doi.org/10.1177/0962280218811349.
Ding YD, Yang R, Yan CG, Chen X, Bai TJ, Bo QJ, et al. Disrupted hemispheric connectivity specialization in patients with major depressive disorder: evidence from the REST-meta-MDD Project. J Affect Disord. 2021. https://doi.org/10.1016/j.jad.2021.02.030.
Rosenthal R,RL, R. Essentials of Behavioral Research: Methods and Data Analysis. 3rd edn, (McGraw-Hill, New York, 1991).
Ahmed AT, Frye MA, Rush AJ, Biernacka JM, Craighead WE, McDonald WM. et al. Mapping depression rating scale phenotypes onto research domain criteria (RDoC) to inform biological research in mood disorders. J Affect Disord. 2018. https://doi.org/10.1016/j.jad.2018.05.005.
Rahman NA. A course in theoretical statistics. (Charles Griffin and Company, New York, 1968).
Pereira F, Mitchell T, Botvinick M. Machine learning classifiers and fMRI: a tutorial overview. Neuroimage. 2009. https://doi.org/10.1016/j.neuroimage.2008.11.007.
Mourao-Miranda J, Reinders AA, Rocha-Rego V, Lappin J, Rondina J, Morgan C, et al. Individualized prediction of illness course at the first psychotic episode: a support vector machine MRI study. Psychol Med. 2012. https://doi.org/10.1017/S0033291711002005.
Tadayonnejad R, Yang S, Kumar A, Ajilore O. Multimodal brain connectivity analysis in unmedicated late-life depression. PLoS ONE. 2014. https://doi.org/10.1371/journal.pone.0096033.
Kerestes R, Harrison BJ, Dandash O, Stephanou K, Whittle S, Pujol J, et al. Specific functional connectivity alterations of the dorsal striatum in young people with depression. Neuroimage Clin. 2015. https://doi.org/10.1016/j.nicl.2014.12.017.
Ochsner KN, Gross JJ. The cognitive control of emotion. Trends Cogn Sci. 2005. https://doi.org/10.1016/j.tics.2005.03.010.
Levesque J, Eugene F, Joanette Y, Paquette V, Mensour B, Beaudoin G, et al. Neural circuitry underlying voluntary suppression of sadness. Biol Psychiatry. 2003. https://doi.org/10.1016/s0006-3223(02)01817-6.
Hercher C, Turecki G, Mechawar N. Through the looking glass: examining neuroanatomical evidence for cellular alterations in major depression. J Psychiatr Res. 2009. https://doi.org/10.1016/j.jpsychires.2009.01.006.
Rajkowska G. Postmortem studies in mood disorders indicate altered numbers of neurons and glial cells. Biol Psychiatry. 2000. https://doi.org/10.1016/s0006-3223(00)00950-1.
Rajkowska G, O’Dwyer G, Teleki Z, Stockmeier CA, Miguel-Hidalgo JJ. GABAergic neurons immunoreactive for calcium binding proteins are reduced in the prefrontal cortex in major depression. Neuropsychopharmacology. 2007. https://doi.org/10.1038/sj.npp.1301234.
Jin C, Gao C, Chen C, Ma S, Netra R, Wang Y, et al. A preliminary study of the dysregulation of the resting networks in first-episode medication-naive adolescent depression. Neurosci Lett. 2011. https://doi.org/10.1016/j.neulet.2011.08.017.
Grimm S, Ernst J, Boesiger P, Schuepbach D, Hell D, Boeker H, et al. Increased self-focus in major depressive disorder is related to neural abnormalities in subcortical-cortical midline structures. Hum Brain Mapp. 2009. https://doi.org/10.1002/hbm.20693.
Tao H, Guo S, Ge T, Kendrick KM, Xue Z, Liu Z. et al. Depression uncouples brain hate circuit. Mol Psychiatry. 2013. https://doi.org/10.1038/mp.2011.127.
Xie C, Goveas J, Wu Z, Li W, Chen G, Franczak M, et al. Neural basis of the association between depressive symptoms and memory deficits in nondemented subjects: resting-state fMRI study. Hum Brain Mapp. 2012. https://doi.org/10.1002/hbm.21291.
Chen G, Chen P, Gong J, Jia, Y, Zhong, S, Chen, F. et al. Shared and specific patterns of dynamic functional connectivity variability of striato-cortical circuitry in unmedicated bipolar and major depressive disorders. Psychol Med. 2020. https://doi.org/10.1017/S0033291720002378.
Pizzagalli DA, Jahn AL, O’Shea JP. Toward an objective characterization of an anhedonic phenotype: a signal-detection approach. Biol Psychiatry. 2005. https://doi.org/10.1016/j.biopsych.2004.11.026.
Pechtel P, Dutra SJ, Goetz EL, Pizzagalli DA. Blunted reward responsiveness in remitted depression. J Psychiatr Res. 2013. https://doi.org/10.1016/j.jpsychires.2013.08.011.
Zhang B, Lin P, Shi H, Ongur D, Auerbach RP, Wang X et al. Mapping anhedonia-specific dysfunction in a transdiagnostic approach: an ALE meta-analysis. Brain Imaging Behav. 2016. https://doi.org/10.1007/s11682-015-9457-6.
Rzepa E, Fisk J, McCabe, C. Blunted neural response to anticipation, effort and consummation of reward and aversion in adolescents with depression symptomatology. J Psychopharmacol. 2017. https://doi.org/10.1177/0269881116681416.
Heshmati M, Russo, SJ. Anhedonia and the brain reward circuitry in depression. Curr Behav Neurosci Rep. 2015. https://doi.org/10.1007/s40473-015-0044-3.
Kumar P, Goer F, Murray L, Dillon DG, Beltzer ML, Cohen AL. et al. Impaired reward prediction error encoding and striatal-midbrain connectivity in depression. Neuropsychopharmacology. 2018. https://doi.org/10.1038/s41386-018-0032-x.
Shi J, Geng J, Yan R, Liu X, Chen Y, Zhu R, et al. Differentiation of transformed bipolar disorder from unipolar depression by resting-state functional connectivity within reward circuit. Front Psychol. 2018. https://doi.org/10.3389/fpsyg.2018.02586.
Xia X, Fan L, Cheng C, Eickhoff SB, Chen J, Li H, et al. Multimodal connectivity-based parcellation reveals a shell-core dichotomy of the human nucleus accumbens. Hum Brain Mapp. 2017. https://doi.org/10.1002/hbm.23636.
Liu R, Wang Y, Chen X, Zhang Z, Xiao L, Zhou Y. Anhedonia correlates with functional connectivity of the nucleus accumbens subregions in patients with major depressive disorder. Neuroimage Clin. 2021. https://doi.org/10.1016/j.nicl.2021.102599.
Smallwood J, Bernhardt BC, Leech R, Bzdok D, Jefferies E, Margulies DS. The default mode network in cognition: a topographical perspective. Nat Rev Neurosci. 2021. https://doi.org/10.1038/s41583-021-00474-4.
Olivo G, Wiemerslage L, Nilsson EK, Solstrand Dahlberg L, Larsen AL, Olaya Bucaro M, et al. Resting-state brain and the FTO obesity risk allele: default mode, sensorimotor, and salience network connectivity underlying different somatosensory integration and reward processing between genotypes. Front Hum Neurosci. 2016. https://doi.org/10.3389/fnhum.2016.00052.
Sayali C, Badre D. Neural systems of cognitive demand avoidance. Neuropsychologia. 2019. https://doi.org/10.1016/j.neuropsychologia.2018.06.016.
Bluhm R, Williamson P, Lanius R, Theberge J, Densmore M, Bartha R, et al. Resting state default-mode network connectivity in early depression using a seed region-of-interest analysis: decreased connectivity with caudate nucleus. Psychiatry Clin Neurosci. 2009. https://doi.org/10.1111/j.1440-1819.2009.02030.x.
Hwang JW, Xin SC, Ou YM, Zhang WY, Liang YL, Chen J, et al. Enhanced default mode network connectivity with ventral striatum in subthreshold depression individuals. J Psychiatr Res. 2016. https://doi.org/10.1016/j.jpsychires.2016.02.005.
Sigurdardottir HM, Gauthier I. In: Toga AW, editor. Brain mapping. Academic Press, Cambridge, Massachusetts, 2015. p. 523–7.
Tang Y, Li Y, Wang N, Li H, Li H, Wang J. The altered cortical connectivity during spatial search for facial expressions in major depressive disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2011. https://doi.org/10.1016/j.pnpbp.2011.08.006.
Garrett A, Kelly R, Gomez R, Keller J, Schatzberg AF,Reiss AL. Aberrant brain activation during a working memory task in psychotic major depression. Am J Psychiatry. 2011. https://doi.org/10.1176/appi.ajp.2010.09121718.
McClure SM, Ericson KM, Laibson DI, Loewenstein G, Cohen JD. Time discounting for primary rewards. J Neurosci. 2007. https://doi.org/10.1523/JNEUROSCI.4246-06.2007.
Zhang J, Wang J, Wu Q, Kuang W, Huang X, He Y, et al. Disrupted brain connectivity networks in drug-naive, first-episode major depressive disorder. Biol Psychiatry. 2011. https://doi.org/10.1016/j.biopsych.2011.05.018.
Lang PJ, Bradley MM, Fitzsimmons JR, Cuthbert BN, Scott JD, Moulder B, et al. Emotional arousal and activation of the visual cortex: an fMRI analysis. Psychophysiology. 1998. https://doi.org/10.1111/1469-8986.3520199.
Zou Q, Long X, Zuo X, Yan C, Zhu C, Yang Y, et al. Functional connectivity between the thalamus and visual cortex under eyes closed and eyes open conditions: a resting-state fMRI study. Hum Brain Mapp. 2009. https://doi.org/10.1002/hbm.20728.
Yan C, Liu D, He Y, Zou Q, Zhu C, Zuo X, et al. Spontaneous brain activity in the default mode network is sensitive to different resting-state conditions with limited cognitive load. PLoS ONE. 2009. https://doi.org/10.1371/journal.pone.0005743.
Cheng Y, Xu J, Arnone D, Nie B, Yu H, Jiang H, et al. Resting-state brain alteration after a single dose of SSRI administration predicts 8-week remission of patients with major depressive disorder. Psychol Med. 2017. https://doi.org/10.1017/S0033291716002440.
Ang YS, Kaiser R, Deckersbach T, Almeida J, Phillips ML, Chase HW, et al. Pretreatment reward sensitivity and frontostriatal resting-state functional connectivity are associated with response to bupropion after sertraline nonresponse. Biol Psychiatry. 2020. https://doi.org/10.1016/j.biopsych.2020.04.009.
Acknowledgements
This study was supported by grants from the National Natural Science Foundation of China (Grant No. 82171508), Natural Science Foundation of Hunan (Grant No. 2020JJ4784), Science and Technology Program of Hunan Province (Grant No. 2020SK53413), and Key-Area Research and Development Program of Guangdong Province (2018B030334001).
Author information
Authors and Affiliations
Contributions
Y-DD and W-BG designed the research, analyze the data and wrote the paper. W-BG and the other authors collected data and were contributors to REST-meta-MDD project.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ding, YD., Chen, X., Chen, ZB. et al. Reduced nucleus accumbens functional connectivity in reward network and default mode network in patients with recurrent major depressive disorder. Transl Psychiatry 12, 236 (2022). https://doi.org/10.1038/s41398-022-01995-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-022-01995-x
