
Abstract
The sensitivity of reef-building coral to elevated temperature is a function of their symbiosis with dinoflagellate algae in the family Symbiodiniaceae. Changes in the composition of the endosymbiont community in response to thermal stress can increase coral thermal tolerance. Consequently, this mechanism is being investigated as a human-assisted intervention for rapid acclimation of coral in the face of climate change. Successful establishment of novel symbioses that increase coral thermal tolerance have been demonstrated in laboratory conditions; however, it is unclear how long these heterologous relationships persist in nature. Here, we test the persistence of a novel symbiosis between Acropora palmata and Durusdinium spp. from Mote Marine Laboratory’s ex situ nursery by outplanting clonal replicates (ramets) of five A. palmata host genotypes to natural reefs in the lower Florida Keys. Amplicon sequencing analysis of ITS2-type profiles revealed that the majority of surviving ramets remained dominated by Durusdinium spp. two years after transplantation. However, 15% of ramets, including representatives of all genotypes, exhibited some degree of symbiont shuffling or switching at six of eight sites, including complete takeover by site-specific strains of the native symbiont, Symbiodinium fitti. The predominant long-term stability of the novel symbiosis supports the potential effectiveness of symbiont modification as a management tool. Although, the finding that 6–7 year-old coral can alter symbiont community composition in the absence of bleaching indicates that Symbiodiniaceae communities are indeed capable of great flexibility under ambient conditions.
Similar content being viewed by others
Increasingly frequent and severe thermal stress events are the predominant threat to reef ecosystems in the Anthropocene. Thermal stress perturbs the photosynthetic mechanism and can result in loss or expulsion of the dinoflagellate algae in the process known as bleaching [1]. While persistent bleaching can result in coral death due to the loss of autotrophic carbon contributed by the symbionts [2], bleaching can also precipitate an acclimatory change in the composition of the endosymbiont community that can subsequently increase coral thermal tolerance [3, 4]. For coral hosting multiple symbiont types, changes in their relative abundance, or “shuffling”, can facilitate survival and recovery [3]. The ability for adult coral to acquire completely new symbiont types following bleaching was also recently confirmed [5]. Although this capacity for flexibility is not universal [6], is influenced by environmental cues, and physiological trade-offs [7], symbiont shuffling and/or switching has been proposed as an important target for human-assisted evolution [8]. Successful establishment of novel symbioses that increase bleaching tolerance have been demonstrated in laboratory conditions [9]; however, it is unclear how long these heterologous relationships persist, especially in natural reef environments. In an essential test of the utility of this approach, we investigate the long-term stability of a heterologous coral symbiosis in the context of reef restoration.
Acropora palmata were once one of the dominant habitat builders of shallow Caribbean reefs [10]. Unprecedented demographic declines since the mid-1970’s have resulted in extensive propagation efforts in both land and ocean-based nursery systems to maintain existing genetic diversity and produce biomass for outplanting efforts aimed at restoring ecosystem structure and function [11]. Naturally occurring populations of A. palmata are highly specific in their symbiosis with Symbiodinium fitti, formerly ITS2-type A3 [12, 13]. Single nucleotide polymorphism genotyping of ex situ nursery-reared lines of A. palmata, and work by Gantt et al. (in press) [14] revealed the dominance of Durusdinium spp. and presence of Cladocopium in Mote A. palmata genotypes (Fig. S1A, Supplementary Methods, GenBank Accessions: ON455821–ON455829) [14]. This provided a unique opportunity to test the maintenance of this heterologous symbiotic relationship after outplanting back to the reef environment.
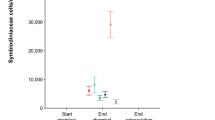
In April 2018, we transplanted three clonal replicates (ramets) of five exclusively ex situ nursery-reared A. palmata genotypes to nine reefs in the lower Florida Keys (n = 27 ramets per genet) to evaluate the long-term stability of the Durusdinium association on natural reefs using periodic phenotypic monitoring and ITS2 amplicon sequencing for symbiont typing using the SymPortal pipeline [15] (Supplementary Methods, Fig. 1). A random sub-sampling of 10–12 ramets per genet confirmed that no Symbiodinium fitti were detectable prior to outplanting in most genotypes (Supplementary Results, Supplementary Table 3 and Supplementary Fig. 2 and Fig. 2A). Four genets were exclusively dominated by a Durusdinium ITS2 type, whereas genet 13-XK showed consistent co-infection with a Cladocopium ITS2 type composed of three co-occurring C1 amplicon sequence variants at T0 (Fig. 2A). Four sequence variants (0.1% of reads) matching to an S. fitti type were identified in one ramet of genet 13-XK at T0 (Supplementary Table 3). An ordination of T0 samples confirmed a significant effect of genotype (p < 0.001) with 13-XK driving divergence (Fig. 2B).

All surviving corals were sampled at time point 24 (T24).

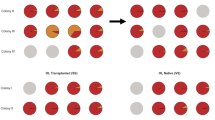
A The relative abundance plot shows the ITS2 symbiont profiles of each individual coral grouped by genotype that was subsampled at T0. Each bar represents an individual and each is labeled with either their ex situ nursery ID or their outplant site ID. Color represents symbiont type. B The relative abundance plot shows the ITS2 symbiont profiles of each individual coral grouped by genotype that was sampled at T24. Each bar represents an individual. Samples are grouped by site, then genotype, and then frag id. Asterisks (*) identify individuals that were also sampled at T0. Color represents symbiont type. C A PCoA plot that shows that symbiont profiles did not significantly change after 2 years of being outplanted. Color represents coral genotype and shape represents site. The ex situ nursery and T0 is included as a site.
No bleaching was observed during quarterly site visits, but site-specific mortality was evident, with 100% ramet mortality at Bahia Honda following 18 months (Supplementary Methods, Table S2). At the remaining eight sites, Durusdinium remained the dominant ITS2 profile after 2 years of outplanting (T24, Fig. 2B, C). Surviving co-infected 13-XK ramets also shuffled to Durusdinium dominance across sites (Fig. 2C) and a PERMANOVA confirmed genotype was no longer a significant factor (p = 0.764). Novel symbiont types were detected at T24, with site-specific patterns (PERMANOVA, p < 0.031, Fig. 2C, Supplementary Table 3 and Supplementary Fig. 3). The homologous symbiont Symbiodinium fitti (A3) was observed at four sites, with ITS2-type profiles indicating potential sub-species or strain-level differences between Dave’s Ledge, Eastern Dry Rocks, Looe Key, and Maryland Shoals (Fig. 2C and Supplementary Table 3). Two ramets of 13-X7 acquired Cladocopium types which were unobserved at T0, including one apparently novel strain (C66) although their relative abundance remained low (Fig. 2C and Supplementary Table 3). A Gerakladium (G) type profile was also detected in ramets of two different genets at two different sites, also at low relative abundance (Fig. 2C and Supplementary Table 3).
The general long-term stability of the heterologous symbiosis between A. palmata and Durusdinium spp. on natural reefs supports the potential effectiveness of symbiont modification as a management intervention, although the ethical implications and long-term ramifications remain to be considered. Infection of coral larvae from mass spawning events with thermally tolerant symbionts and outplanting them to degraded reefs could also be a restoration tool [16]. However, the occasional (re)acquisition of homologous, site-specific strains of Symbiodinium fitti indicates that the temporal dynamics of symbiosis establishment in coral may be more complicated than previously thought. While corals can uptake symbionts in their larval phase, the current consensus is that the majority of symbionts are acquired after settlement. New recruits and juvenile corals can form a variety of novel symbiont associations [17], but this diversity is generally pruned to one numerically dominant taxon per genus over the first few years of life [18]. Consensus suggests that changes to the dominant symbiont taxon in adult coral occur in response to bleaching-level stress with reversion to the dominant symbiont profile in the absence of bleaching stress [3, 5, 19].
The A. palmata used in this study were produced using individual larvae from batch crosses in 2013/14. Although physically small, at the time of outplanting, experimental ramets were 4–5 years old, which under field conditions is the age of sexual maturity for this species [20]. The finding that adult corals can shuffle or switch their symbionts in situ in the absence of bleaching suggests that acquisition and winnowing may be dependent on other factors rather than a simple age-dependent process. It is also possible that Symbiodiniaceae communities are indeed capable of greater flexibility under ambient conditions [21]. However, the stability of the heterologous symbiosis in most individuals suggests that infection with a novel symbiont type as a recruit and maintenance till maturity could produce a stable symbiosis.
Data availability
Sequence Access: NCBI, Accession: PRJNA868513, https://doi.org/10.5281/zenodo.7419654.
References
Hoegh-Guldberg O, Smith JG. The effect of sudden changes in temperature, light, and salinity on the population density and export of zooxanthellae from the reef corals Stylophora pistillata (Esper) and Seriatopora hysterix (Dana). J Exp Mar Biol Ecol. 1989;129:279–303.
Glynn PW. Coral reef bleaching: ecological perspectives. Coral Reefs. 1993;12:1–17.
Berkelmans R, van Oppen MJH. The role of zooxanthellae in the thermal tolerance of corals: a “nugget of hope” for coral reefs in an era of climate change. Proc R Soc B: Biol Sci. 2006;273:2305–12.
Cunning R, Gillette P, Capo T, Galvez K, Baker AC. Growth tradeoffs associated with thermotolerant symbionts in the coral Pocillopora damicornis are lost in warmer oceans. Coral Reefs. 2015;34:155–60.
Scharfenstein HJ, Chan WY, Buerger P, Humphrey C, van Oppen MJH. Evidence for de novo acquisition of microalgal symbionts by bleached adult corals. ISME J. 2022;16:1676–9.
Goulet TL. Most corals may not change their symbionts. Mar Ecol Prog Ser. 2006;321:1–7.
Jones A, Berkelmans R. Potential costs of acclimatization to a warmer climate: growth of a reef coral with heat tolerant vs. sensitive symbiont types. PLoS ONE. 2010;5:e10437.
van Oppen MJH, Oliver JK, Putnam HM, Gates RD. Building coral reef resilience through assisted evolution. Proc R Soc B: Biol Sci. 2015;112:2307–13.
Buerger P, Alvarez C, Coppin CW, Pearce SL, Chakravarti LJ, Oakeshott JG, et al. Heat-evolved microalgal symbionts increase coral bleaching tolerance. Sci Adv. 2020;6:eaba2498.
Kuffner IB, Toth LT. A geological perspective on the degradation and conservation of western Atlantic coral reefs. Conserv Biol: J Soc Conserv Biol. 2016;30:706–15.
Young CN, Schopmeyer SA, Lirman D. A review of reef restoration and Coral propagation using the threatened genus Acropora in the Caribbean and western Atlantic. Bull Mar Sci. 2012;88:1075–98.
Reich HG, Kitchen SA, Stankiewicz KH, Devlin-Durante M, Fogarty ND, Baums IB. Genomic variation of an endosymbiotic dinoflagellate (Symbiodinium fitti) among closely related coral hosts. Mol Ecol. 2021;30:3500–14.
Baums IB, Devlin-Durante MK, Lajeunesse TC. New insights into the dynamics between reef corals and their associated dinoflagellate endosymbionts from population genetic studies. Mol Ecol. 2014;23:4203–15.
Gantt SE, Keister E, Manfroy A, Merck D, Fitt W, Muller E, et al. Wild and nursery-raised corals: comparative physiology of two framework coral species. Coral Reefs. (In Press).
Hume BCC, Smith EG, Ziegler M, Hugh J, Warrington M, Burt J, et al. SymPortal: a novel analytical framework and platform for coral algal symbiont next-generation sequencing ITS2 profiling. Mol Ecol Resour. 2019;19:1063–80.
Randall CJ, Negri AP, Quigley KM, Foster T, Ricardo GF, Webster NS, et al. Sexual production of corals for reef restoration in the Anthropocene. Mar Ecol Prog Ser. 2020;635:203–32.
Bay LK, Cumbo VR, Abrego D, Kool JT, Ainsworth TD, Willis BL. Infection dynamics vary between Symbiodinium types and cell surface treatments during establishment of endosymbiosis with coral larvae. Diversity. 2011;3:356–74.
Abrego D, van Oppen MJH, Willis BL. Highly infectious symbiont dominates initial uptake in coral juveniles. Mol Ecol. 2009;18:3518–31.
Cunning R, Silverstein RN, Baker AC. Investigating the causes and consequences of symbiont shuffling in a multi-partner reef coral symbiosis under environmental change. Proc R Soc B: Biol Sci. 2015;282:20141725.
Chamberland VF, Petersen D, Latijnhouwers KRW, Snowden S, Mueller B, Vermeij MJA. Four-year-old Caribbean Acropora colonies reared from field-collected gametes are sexually mature. Bull Mar Sci. 2016;92:263–4.
Silverstein RN, Correa AMS, Baker AC. Specificity is rarely absolute in coral–algal symbiosis: implications for coral response to climate change. Proc R Soc B: Biol Sci. 2012;279:2609–18.
Acknowledgements
We thank the staff of Mote Marine Laboratory for their assistance in producing and maintaining the coral genotypes used in this study. We also thank all graduate students of the Cnidarian Evolutionary Ecology Lab and Mote Marine Laboratory staff who participated in monitoring of these corals during the outplant period. The transplant of coral colonies for the purpose of coral restoration was permitted under FKNMS-2015-163 and FKNMS-2015-163-A3 to EB. Tissue sampling of transplants was permitted under FKNMS-2018-035 to CDK.
Funding
This research was supported by National Oceanic and Atmospheric Administration Coral Reef Conservation Program grant NA17NOS4820084, and private funding from the Alfred P. Sloan, and Rose Hills Foundations.
Author information
Authors and Affiliations
Contributions
CDK, EMM, CJK, WCM, and HE conceived the experiments and analysis. WCM, EB, CJK, and CDK conducted experiments. EMM and EB provided advice and logistical support through the experiment. HE conducted genetic and statistical analysis and wrote the manuscript. CDK and EMM provided advice and support for analysis. CDK, EMM, and CJK helped write the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Elder, H., Million, W.C., Bartels, E. et al. Long-term maintenance of a heterologous symbiont association in Acropora palmata on natural reefs. ISME J 17, 486–489 (2023). https://doi.org/10.1038/s41396-022-01349-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41396-022-01349-x
