
Abstract
Global-scale N-oxide contamination of groundwater within aquifers occurs due to the widespread use of N-bearing fertilizers and chemicals, threatening both human and environmental health. However, the conversion of these pollutants in active nitrogen (N) cycling processes in the subsurface biosphere still remains unclear. This study investigates the global occurrence of anaerobic ammonium oxidation (anammox) in aquifers, where anammox was found to be turned on and off between saturated and unsaturated soil horizons, and contributed 36.8–79.5% to N loss in saturated soil horizons, the remainder being due to denitrification which has traditionally been considered the main pathway for removal of N-pollutants from aquifers. Although anammox activity was undetectable in the unsaturated soil horizons, it could potentially be activated by contact with ascending groundwater. High-throughput pyrosequencing analysis identified Candidatus Brocadia anammoxidans as being the most abundant anammox bacterium in the saturated soils investigated. However, the anammox bacterial abundance was determined by the relative richness of Candidatus Jettenia asiatica. Isotopic pairing experiments revealed that coupling anammox with ammonium oxidation and respiratory ammonification enabled the formation of a revised N cycle in aquifer systems, in which respiratory ammonification acted as an important coordinator. Anammox can therefore contribute substantially to aquifer N cycling and its role in remediation of aquifers contaminated with N-oxides may be of global importance.
Similar content being viewed by others

Introduction
Groundwater is the largest global freshwater resource accessible and plays a central role in drinking water production, agricultural irrigation and global food security [1, 2]. Groundwater is stored in permeable geologic units known as aquifers, with N-oxide contamination of these aquifers presenting significant global problems from both human and environmental health perspectives [3,4,5]. N-oxide pollution results in high nitrate (NO3–) concentrations in aquifers, whereby nitrate is a strong precursor for the formation of carcinogenic nitrosamine. In addition, high nitrate levels in drinking water may cause methaemoglobinaemia via nitrite binding to hem, a form of N-poisoning particularly threatening infants [6]. For decades, it has generally been accepted that heterotrophic denitrification governs nitrogen (N) cycling in aquifers, with this process being the only known pathway for N loss [4, 7], while it still remains unclear what in detail governs active N cycling processes in the terrestrial subsurface associated with aquifers.
Aquifers are generally characterized by long water residence times (global average of 1400 years), with relatively slow water exchange rates and low organic matter [1, 2]. Therefore, aquifers are a potential niche for slow growing autotrophic bacteria. Anaerobic ammonium oxidation (anammox) is mediated by autotrophic bacteria that can oxidize ammonium (NH4+) with nitrite (NO2–) to N2 without the need for an organic electron donor [8,9,10,11]. Research to date has shown that anammox bacteria are widely distributed and contribute significantly to N-loss in marine and terrestrial surface aquatic ecosystems [12,13,14,15,16,17,18]. Anammox bacteria grow slowly with a doubling time of 14–20 days and are therefore, only competitive in environments with relatively minor water exchange rates [8], making aquifers a good potential niche environment for the growth of anammox bacteria.
Previous studies on anammox bacteria in diverse ecotones have indicated that the interface between the two adjacent biomes often contains highly active zones for biogeochemical N cycling, with significant interactions and fluxes between oxidized and reduced N [17, 19,20,21,22,23,24]. The development of new agricultural practices to satisfy the continually growing global demand for food means that the NH4+ contamination of surface waters is increasing worldwide. Furthermore, some groundwater sources are rich in NO2–, which is a major driver in global NO3– flux [1, 4]. In aquifers, NH4+ is partly adsorbed by clay minerals and partly oxidized to NO3−. Oxidized and reduced N can interact at the groundwater table, resulting in transfer between saturated and unsaturated soils. The frequent interaction between surface water and groundwater in aquifer zones provides a unique niche for anammox bacteria [4, 25]. Therefore, it may be inferred that anammox bacteria are likely to globally occur in deep soils, such as in aquifer environments, which serve as active ecotone between surface water and groundwater. Based on previous studies on anammox in aquatic ecosystems [15, 17, 19, 22], the hypothesis of this study is that anammox bacteria can thrive in subsurface aquifer systems and play an important role in N-remediation.
The goal of this investigation was to identify processes that may remove N-oxide pollutants from aquifers. Previous models and practical studies have indicated that anammox bacteria are active in some contaminated groundwater environments (Supplementary Table S1) [26,27,28]. However, the significance of the anammox processes to N-loss and its associated mechanisms remain unknown. Therefore, this study combined isotope tracer, as well as biogeochemical and microbial ecological methods, to investigate the occurrence, distribution, contribution and biogeochemistry of anammox in groundwater aquifers worldwide.
Methods
Study site background
This study initially assessed the hypothesis that anammox occurs in terrestrial subsurface environments on a regional scale (China-Anxin, 24 km transect, 1.5–2.6 m depth). Results were then confirmed using extensive global-scale sampling, from four continents. A total of 1088 samples were collected from different soil depths from sites in the following locations: Australia-Kalgoorlie (21.0 m depth below surface, Gobi yellow brown soil); China-Beijing (28.0 m depth, grassland yellow clunamon soil); China-Wuqing (20.0 m depth, farmland yellow clunamon soil); Switzerland-Zurich (9.9 m depth in summer and 10.8 m depth in winter, grassland cinnamon soil); Mozambique-Nampula (8.0 m depth, desert red soil); China-Antu (2.0 m depth, forest black soil); China-Jilin (1.4 m depth, farmland black soil); Germany-Bremen (1.2 m depth, forest black soil); and Australia-Queensland (1.1 m depth in spring and 1.0 m depth in autumn, farmland brown soil). Temporal evidence gathered from seasonal sampling in Switzerland–Zurich and Australia-Queensland was also examined to assess the hypothesis. This sample suite represents a range of environmental factors, including various soil types, parent substrates, geochemical properties, organic matter content and composition, genetic classification, groundwater table characteristics, nitrogenous compound concentrations and temperature (Supplementary Fig. S1, Table S2).
Sample collection
Depth-resolved soil cores were collected using an auger (7 cm diameter, Beijing New Landmark Soil Equipment, Beijing, China) from sites with a high groundwater table (<2.0 m depth), using GeoProbe (GeoProbe, USA; 2.0–20.0 m depth) and PowerProbe (AMS, USA; >20.0 m depth) direct push drill rigs, and sectioned at certain intervals based on the drill rig model used. At each sampling site, five parallel quadrats (5 × 5 m; all four corners and the central point) were selected to collect soil samples, within a total plot of approximately 2500 m2 (50 × 50 m). For each quadrat, five sample pits were drilled (all four corners and the central point), and from corner pits continuous soil profile samples were collected and sliced at specific intervals. For each group of equal-depth samples a composite sample was generated via thorough mixing. The central sample was used to measure the dissolved oxygen (DO) concentration in triplicate and to collect groundwater samples. All soil samples were placed in sterile plastic bags and transported to the laboratory on ice for subsequent analysis of molecular and microbial activity, as well as chemical analysis. The incubation media was composed of groundwater samples collected from different depths using a peristaltic pump, stored in sterile vacuum bottles and transported to the laboratory at 4 °C. Upon arrival at the laboratory, the groundwater samples were filtered through 0.22 μm mixed cellulose esters microfiltration membranes (MF-Millipore, Billerica, MA, USA) to remove biological organisms but retain nutrients to serve as the incubation media. Overall, for each sampling site we obtained 1~2 kg of sample per soil depth and 800–1000 mL of groundwater samples. All samples were measured separately and mean values were used to represent the average site conditions.
Potential rate of anammox, denitrification, ammonium oxidation, nitrite oxidation, nitrate reduction (15NO3 −→15NO2 −) and respiratory ammonification with the 15N-tracer technique
Different 15N-amended substrates were added to determine the potential rate of different N-cycling processes, immediately after the fresh samples arrived at the laboratory. The homogenized soil samples (approximately 3.3 g each) were transferred to 12-mL glass vials (Exetainer, Labco, High Wycombe, UK) and combined with the natural water media at ambient temperature to form slurries. Three parallel assays were conducted for each slurry sample. Stock solutions of each isotopic mixture (Na15NO3 [15N at 99.19%]; (15NH4)2SO4 [15N at 99.16%]; Na15NO2 [15N at 98.17%]; (14NH4)2SO4; and Na14NO3) were purchased from Shanghai Research Institute of Chemical Industry, China. Final concentrations were a maximum of 10% of the initial background concentration, minimizing the possibility of overestimating the potential rate [22, 24, 29] (Supplementary Table S3). The dissolved oxygen (DO) level of each resulting slurry was adjusted before incubation by adding ultra-pure He (99.99%) or O2 (99.99%) (Beijing Huayuan Gas, Beijing, China), to establish equivalent in situ DO conditions (±5% deviation).
To determine the potential rates of anammox and denitrification, the resulting slurries were initially preincubated to deplete residual NOx− within soils and incubation media according to refs. [29, 30]. Subsequently, 100 μL of N2-purged stock solution was injected into each treatment, i.e. (1) negative control: 15NH4+, (2) positive control: 15NH4+ + 14NO2–, and (3) 15NO2– (Supplementary Fig. S2, Table S3). The incubations were stopped at defined intervals (0, 4, 10, 24 and 48 h) by adding 200 μL of 7 M ZnCl2 solution.
Potential rates of ammonium oxidation were measured as the rates of 15NO2– production from incubations with 15NH4+ under in situ DO conditions. This was assumed to provide the minimum estimate without inclusion of 15NO3– produced from 15NH4+. First, 100 μL of diluted 15NH4+ isotopic stock solution was added to slurries (Supplementary Table S3) and the mixtures were incubated accordingly, with incubation of the slurries terminated at defined intervals. To assess the N-isotopic ratio of 15NO2–, sulfamic acid was added to form a final concentration of 16.5 mM, to reduce 15NO2– to 29N2 [31, 32]. The sulfamic acid mixtures were incubated for more than 12 h to ensure sufficient conversion and minimize the possibility of further underestimating the potential rates.
Potential rates of nitrite oxidation were detected as the production rates of 15NO3– derived from oxidized 15NO2– under in situ DO conditions. 100 μL of N2-purged 15NO2– isotopic stock solution was first added to incubation. By injecting 100 μL of saturated mercuric chloride solution after defined intervals, about 1.0 g sponge cadmium was added under alkaline (pH = 7.0–8.0) to convert produced 15NO3– to 15NO2–, and then sulfamic acid was added to reduce 15NO2– completely to 29N2 like the above method [31, 32].
To determine the potential rates of nitrate reduction, 100 μL of diluted 15NO3– isotopic stock solution was added to the slurries (Supplementary Table S3) and samples were incubated under in situ DO conditions. One hundred microliters of saturated mercuric chloride was added to terminate the biological activity after defined intervals. Next, sulfamic acid was used to completely reduce 15NO2– to 29N2 comparable to the above method [31, 32].
To assess the potential rates of respiratory ammonification (also referred to as DNRA; Dissimilatory Nitrate/Nitrite reduction to Ammonium), 100 μL of diluted 15NO2– isotopic stock solution was added to the slurries (Supplementary Table S3) under in situ DO conditions. At defined intervals the reaction was stopped and hypobromite was added (at a concentration equal to the concentration of added 15NO2–), with mixtures then incubated for more than 12 h to ensure complete conversion of 15NH4+ to 30N2 [31, 32].
The production of 29N2 or 30N2 was measured by isotope ratio mass spectrometry (IRMS) (MAT253 and Gasbench II, Thermofisher Scientific, Bremen, Germany). Standard curves were established and analyzed together with soil slurries. Demineralized water, equilibrated with atmospheric air at an ambient temperature, was transferred to the exetainer vials, ensuring the vials were completely filled. Subsequently, 8.0 mL of the water was replaced by 99.9% ultra-pure He (Beijing Huayuan Gas, Beijing, China) and 0, 30, 60 or 100 µL air was then injected into individual vails in triplicate. Following this, the glass vials were shaken to obtain equilibrium between the water and headspace. These references are included as values for calibration of total signal versus the amount of N2. Linear regression analysis was conducted between the total signal and the amount of produced N2. According to the raw signal and ratio of 28/29/30N2 analyzed by IRMS, the concentration of N2 produced was calculated and in both reference and soil samples using the average 15NH4+ mole fraction (FA) and 15NOx− mole fraction (FN) according to ref. [33] (Supplementary Table S3, S4). The coefficients of determination (R2) established for linear regression of the labeled 28/29/30N2 concentration shift over time were all >0.90. The precision of the IRMS in this study was established as <0.01‰ and the detection limit was 0.001 nmol N g−1 h−1. Measurements were completed at the Key Laboratory of Tibetan Environment Changes and Land Surface Processes, Institute of Tibetan Plateau Research (ITPR), Chinese Academy of Sciences (CAS).
Molecular (q)PCR assay
Soil composite samples were homogenized and freeze-dried at −50 °C to eliminate the effect of different moisture contents (MC) in saturated and unsaturated soils, using a vacuum freeze drier (Alpha 1-2 LD plus, Germany). Genome DNA was extracted from 0.33 g of freeze-dried soil using the FastDNA SPIN Kit for Soil (MP Biomedicals, Solon, OH, USA), with a bead-beating time of 45 s and a speed setting of 5.5 m s−1 using FastPrep-24 (MP Biomedicals, Solon, OH, USA) according to the manufacturer’s instructions. The extracted DNA was quantitatively analyzed using a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific, Schwerte, Germany), with all ratios of absorbance at 260 and 280 nm ranging between 1.8 and 1.9. A nested-PCR assay was performed to detect the anammox hydrazine synthase gene (hzs) gene [20, 21]. Anammox abundance was determined using qPCR with the primers HSBeta396F/HSBeta742R, targeting a subunit of the hzs gene which is specific for anammox [20, 21]. All tests were performed in triplicate with amplification efficiencies of between 90 and 110%. The correlation coefficients (R2) were all >0.98 and the detection limit was 1.00 × 103 copies g−1. The sequences obtained in this study for anammox bacteria are available in the NCBI database, under accession numbers JQ861456-JQ861531 and HQ847736-HQ847832. The detailed primers and corresponding reaction profiles are listed in Supplementary Information and Table S5.
Illumina hzsB amplicon sequencing and molecular ecological network analysis
The hzsB gene (approximately 290 nucleotides) was amplified with the barcoded primer pairs 449F-barcode and 742R. The PCR products were sequenced using a Hiseq 2500 platform (Illumina, CA, USA) with a PE250 strategy (2 × 250 bp). High-quality unique nucleic acid sequences were obtained through quality filtering using QIIME [34]. Operational taxonomic units (OTUs) were clustered at the 97% identity level. Downstream analysis and calculations were performed with QIIME and Mothur software [35]). LefSe (http://huttenhower.sph.harvard.edu/galaxy/) and Circos (http://circos.ca/) analysis were performed to identify significant differences in the relative richness of OTUs and the distribution of anammox species across all global saturated soils, respectively. Molecular ecological networks were constructed using the obtained OTUs [36]. The online pipeline was provided by the Institute of Environmental Genomics, University of Oklahoma. Details are shown in Supplementary Information.
Analytical procedures for soil/sediment environmental variables
Soil samples were dried, homogenized and filtered through a 2.0 mm sieve, prior to physicochemical analysis. Ammonium (NH4+), nitrite (NO2−) and nitrate (NO3–) were extracted from 6 g of sieved soil by shaking for 1 h at room temperature with 30 ml 2 mol L−1 KCl solution and measured using an Auto-Analyzer HR (Seal Analytical, Southampton, UK). The pH value was analyzed using a DELTA 320 pH Analyzer (Mettler Toledo, USA) in a suspension of 5 g sieved soil and 12.5 mL ddH2O (a soil-to-water ratio of 1:2.5). The concentration of total organic matter (TOM) was determined by oxidation with potassium dichromate and sulfuric acid. Total nitrogen (TN), total carbon (TC), and total sulfur (TS) were measured using a Vario EL III Analyzer (Elementar Analyses System GmbH, Germany). The MC was analyzed by oven-drying 2 g of fresh soil/sediment at 108 °C until a constant weight was reached. The DO was detected using a multiprobe optical monitoring sonde (Model 600 XLMV2, YSI, USA) equipped with an optical DO sensor (4.5 cm diameter). According to the standard method of the U.S. Geological Survey Techniques of Water-Resources Investigations [37], the sensor was hermetically sealed into a chamber to avoid aeration and mitigate losses or gains of dissolved gases in the groundwater being sampled. When measuring DO, the groundwater was circulated through the chamber for 20 min to reduce the interference of atmospheric oxygen, until stability was met. To minimize vertical mixing when measuring the temperature (T), the probe was lowered slowly. Each profile required approximately 5 min for data collection and the unit was calibrated before deployment. Triplicates were run for quality assurance/quality control (QA/QC) for all of the above measurements.
Statistical analysis
Basic data management and arrangement was performed in Microsoft Excel 2010. The potential depth-integrated rates were calculated using the potential weight rates multiplied by the soil density and the height of each soil core segment (Supplementary Table S4). Data are expressed as mean values ± standard error (S.E.), while fluctuation level analysis used interquartile range (IQR) data to express a robust range. General statistical analysis was conducted using the statistical product and service solutions software, version 18.0 (SPSS Inc., USA) and software package R. General additive models (GAMs) were used to identify anammox peaks, without considering the effect of seasonality. In addition, mixed effect models were used to investigate the varying effect between microbial abundance/rate and physico-chemical characteristics with site and depth as the effects, in response to differences among groups, i.e. potential rates of anammox and denitrification, peak and nonpeak value, and saturated and unsaturated zones. Prior to analyses, all the predictors and predictand variables were z-score standardized, thus enabling the effects to be compared between sites. The stepwise linear regression (SLR) was used to model the relationships between the anammox abundance/activity and environmental variables. The Wilcoxon signed-rank test was used to identify the significant proportion of the dominant species, among all anammox bacteria. Graphing was achieved using Origin 8.0 software. All statistical differences were set at the α = 0.05 significance level (p ≤ 0.05).
Results
Regional-scale investigation
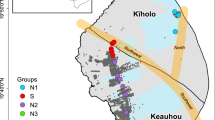
Sampling was conducted along a 24-km long transect in Anxin County, Hebei province, China. A total of 285 soil samples were collected, from six sampling sites along a transect, at depths of 1.5−2.6 m below the ground surface. Ninety samples were collected from unsaturated soils and 195 from saturated soils (Fig. 1, Supplementary Fig. S1 and Table S2). Results show that anammox distribution varied considerably according to depth. Below the groundwater table (0.6 m depth), anammox bacteria Candidatus Brocadia (Ca. Brocadia) and Candidatus Jettenia asiatica (Ca.J. asiatica, similarity of 97.6–98.8%) were detected throughout the saturated soils, at levels of 4.81 × 103–1.98 × 106 gene copies g−1 (Fig. 1a, b, and Supplementary Fig. S3). In contrast, anammox bacterial counts were below the detection limit of <103 copies g−1 in the unsaturated soils.

Global-scale occurrence, significance and contribution of anammox in aquifer systems. a Global overview of the sampling characteristics of upland soil cores and the contribution of anammox to total N-loss in aquifer systems. Pie charts represent the ratios of anammox or denitrification process contributions to N-loss in the soil cores. b The regional scale (24 km, Anxin county, Hebei province, China) occurrence, activity and contribution of anammox in aquifer systems. a−f represent different soil cores with a pair-wise distance of 7.0, 7.0, 4.0, 4.5 and 1.5 km, successively. c The global-scale occurrence, activity and contribution of anammox in aquifer systems, with three counter-examples. Potential rates are expressed as depth-integrated values with flux units of mmol N m−2 d−1. The saturated and unsaturated soil are indicated in light blue and gray, respectively. The anammox abundance was below the detection limit (<103 copies g−1) with no positive 15N tracing results for the potential rate of anammox in unsaturated soil. All potential rate and abundance data are expressed as the mean value ± s.e. (standard error, n = 5)
The 15N-tracer technique confirmed that anammox activity was mainly present in the saturated soils, using positive and negative controls (Fig. 1a, b, and Supplementary Fig. S2). The potential rates of anammox ranged from 0.03 to 6.69 mmol N m−2 d−1, correlating well with the anammox cell abundances. In all transect cores assessed for all six sampling sites, anammox bacteria were found to be turned on and off between the saturated and unsaturated soils. A higher potential rate of anammox (3.38 ± 1.82 mmol N m−2 d−1) consistently maximized in samples from the upper 15% horizons of saturated soils (Supplementary Fig. S4), where the potential rates of anammox were even higher than those of denitrification (1.15 ± 1.33 mmol N m−2 d−1), accounting for 77.9 ± 15.1% of the total N-loss. Anammox decreased along with the depth, and in the non-peak horizons anammox still contributed 37.6 ± 29.7% to N loss.
Global-scale investigation
The hypothesis that anammox occurs in aquifer soils globally was further investigated in soil samples from 11 cores collected from sites around the world. In total, there were 1070 samples, including 860 from unsaturated soils and 210 from saturated soils (Fig. 1a, c, Supplementary Fig. S1 and Table S2). All of the saturated soils showed the presence of anammox bacteria and associated N-loss due to their activity (Fig. 1a, c). Anammox cell numbers varied between 1.92 × 104 and 8.48 × 107 copies g−1, with Ca. Brocadia and Ca. J. asiatica being the most abundant species (similarity 95.6–97.6%, Supplementary Fig. S3). Only one soil layer from the unsaturated soils (Zurich-Summer; depth of 8.4 m) showed the presence of anammox bacteria ([8.84 ± 1.61] × 104 copies g−1). In the saturated soils, anammox activity was detected in 210 samples, at potential rates ranging from 0.02 to 5.01 mmol N m−2 d−1. The potential rates of anammox did not show any significant correlation with soil composition, depth of the groundwater table, or environmental variables, such as NH4+, NO2−, NO3–, pH and so on (Supplementary Table S6), suggesting that substantial anammox activity is likely to be a characteristic of aquifers in general.
In the integrated saturated soils, the potential rates of anammox (6.68 ± 4.17 mmol N m−2 d−1) were at the same level of denitrification (6.81 ± 6.57 mmol N m−2 d−1) (p > 0.05; Supplementary Table S7), and even higher in most sites (5/8), accounting for 54.0 ± 14.8% of the total N-loss (Fig. 1a, c). At the anammox peak layer, anammox even contributed up to an avg. 67.5 ± 20.2% of the total N-loss. Lower potential rates of anammox were found at greater depths, whereby those of denitrification also were relatively low, and anammox still contributed significantly (42.6 ± 28.8%). At the nonpeak horizons, anammox still accounted for 46.7 ± 29.5% of the total N-loss. Together, these results indicate that anammox may play a significant and previously unrecognized role in aquifer N cycling processes.
Temporal-scale investigation
Analysis conducted on a temporal scale showed that the potential rates of anammox changed in accordance with groundwater table fluctuations (Fig. 1a, c, Supplementary Fig. S1 and Table S2). In the Zurich soil cores, the potential rates of anammox were detectable at depths below 8.4 m in the summer, rising to depth of 4.8 m in the winter, respectively ([0.08 ± 0.02] – [4.54 ± 0.26] mmol N m−2 d−1). In the integrated saturated soils, the contribution to N-loss also increased from about 55.3% in summer to 77.6% in winter. In the Queensland soil cores, the potential rates of anammox were detectable below a depth of 0.8 m in autumn only, rising to a depth of 0.6 m in the spring. In the integrated saturated soils, 36.8–63.8% of total N-loss was to be attributed to anammox. In summary, we conclude that anammox bacteria reveal seasonally fluctuating abundance and activity in subsurface aquifer systems worldwide.
In contrast, all molecular and isotopic results indicated that anammox was absent in the soil cores collected from areas without near-surface aquifers, including cores from Beijing (28.0 m depth), Nampula (7.0 m depth) and Kalgoorlie (21.0 m depth) (Fig. 1). These dry soils were then incubated exclusively with Milli-Q water (1:1 in mass) to assess the potential impact of fluctuating groundwater table levels (Fig. 2a). In the Beijing samples from non-aquifer sites, the anammox bacterial counts were still below the detection limit after the first month of incubation but reached a peak ([5.13 ± 0.87] × 105 copies g−1) in the third month (Fig. 2b). The cell numbers decreased to an “ND” level again by the fifth month, as no substrate was supplied during the experimental period. Similar trends were observed in two additional soil samples, with a time lag observed. These results indicate that a rise in groundwater table could potentially activate anammox bacterial growth and sustain the latter, if groundwater would further provide suitable substrates.

Incubation of anammox bacteria with water to simulate an ascending level of the groundwater table. a Outline of incubation experimental strategy. No anammox was detected in the original in situ soils. Soil samples were then incubated with Milli-Q water at a 1:1 mass ratio. b The temporal variation of anammox bacterial abundance and substrate concentrations in soil samples during the incubation period, along with the in situ original soils from the counter-examples (Beijing-10 m sample in Fig. 1c (j), Mozambique-5 m sample in Fig. 1c (i) and Kalgoorlie-21 m sample in Fig. 1c (k)]. “ND” indicates that the anammox abundance was below the detection limit (<103 copies g−1) with no positive 15N tracing results for anammox rate. All data are expressed as the mean value ± s.e. (standard error, n = 5)
Diversity and biogeography
Anammox diversity was also investigated using high-throughput sequencing analysis targeting an anammox diagnostic gene, hydrazine synthase (hzs). Five major anammox clusters were detected in global aquifers (Fig. 3a, Supplementary Fig. S5, and Table S8). Ca. Brocadia anammoxidans (Ca. B. anammoxidans) dominated the anammox community (90.0–95.3%, p < 0.001, Fig. 3a, Supplementary Fig. S5, Supplementary Tables S8 and S9). Conversely, Ca. Scalindua represented a very minor fraction in anammox bacteria in Australian soil cores only. The diversity indices of anammox community were highly heterogeneous globally, according to α-diversity and LEfSe analyses (97% cut-off; Fig. 3b, Supplementary Table S8). The Shannon and Simpson’s diversity indices were negatively correlated with the dominating species Ca. B. anammoxidans (p < 0.01, respectively; Supplementary Table S10), suggesting that a higher relative abundance of Ca. B. anammoxidans may indicate lower diversity within the anammox community. Both statistical and network analyses revealed that TOM, MC, and NO2− content were the fundamental factors in anammox community structure and diversity (Fig. 3c, Supplementary Table S10), with these factors also significantly positively correlated with anammox abundance (p < 0.05, Supplementary Tables S6, S11).

High-throughput sequencing, ecological network analysis and environmental mechanisms of the anammox community in global aquifer systems. a Global overview of the anammox bacterial community structure in saturated soils. Pie charts represent the anammox bacterial diversity and composition. Compositions were determined using high-throughput sequencing analysis (97% cut-off) and were identified via a Neighbor-Joining method using MEGA 6.0, with the hzsB gene sequences of the cultured anammox species. b Different species in the six sites identified by LEfSe with LDA values of 2.0. Colors show the samples with differences in the most abundant OTUs. c Nonlinear RDA ordination plots show the correlations between anammox bacterial community structures and environmental factors. d Molecular ecological networks of the subsurface interface of saturated and unsaturated soils. Nodes represent OTUs, node colors represent different anammox species, and node links show the interactions among different OTUs within the module
The interactive role of anammox species was also found to be highly heterogeneous globally. The anammox abundance was not correlated with most abundant species Ca. B. anammoxidans (p < 0.05) or Ca. B. fulgida (p > 0.05), but significantly positively correlated with Ca. J. asiatica (p < 0.01) (Fig. 3c, Supplementary Table S10). Network analysis also revealed that Ca. J. asiatica, with relative abundance of less than 5%, was the keystone genus linking the anammox community, which may be due to the mixotrophic metabolism of Ca. J. asiatica [38]. In addition, Ca. J. asiatica showed the highest level of positive correlation with MC (p < 0.001) and concentrations of TOM (p < 0.01) and NO2− (p < 0.01) (Supplementary Table S10). These findings indicate that MC was the critical environmental factor for anammox cell material and energy exchange, while TOM and TN supply the basic substrates required for Ca. J. asiatica growth. Therefore, Ca. J. asiatica has the greatest interactive role in the anammox community and abundance within aquifers, even though Ca. B. anammoxidans is the most abundant species.
Interaction of anammox with other N cycling processes
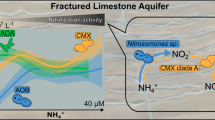
Further studies were performed to determine how the potential rates of anammox compared to those of other N cycle processes in the peak horizon of anammox activity in saturated soil. NO2− is not only an important intermediate in the N cycle, but also a key substrate for anammox reactions. The potential rate of NO2− conversion (anammox, denitrification, NO2– oxidation and respiratory ammonification) and the potential rate of NO2− production (NH4+ oxidation and NO3– reduction) were measured at the anammox peak horizon using the 15N isotopic-pairing technique (Fig. 4). The sources and sinks of NH4+ and NO2– in saturated soils were found to vary significantly, depending on the DO level. Under DO conditions of >0.5 mg L−1, NH4+ oxidation occurred at higher potential rates (53.16 ± 19.50 mmol N m−2 d−1, p < 0.05), providing the main supply of the substrate NO2– (84.6 ± 8.4%, p < 0.01) for both anammox and denitrification processes. The process of respiratory ammonification is usually considered to be significant in peripheral environments only [39, 40]. However, in the present study it was found to occur at high potential rates (10.78 ± 5.13 mmol N m−2 d−1, p < 0.05) when combined with high potential rates of NH4+ oxidation. Consequently, coupling of respiratory ammonification processes with high potential rates of NH4+ oxidation could satisfy a substantial part of the NH4+ requirement for anammox. In addition, respiratory ammonification processes can reduce surplus nitrite to eliminate toxic inhibition. In contrast, under DO conditions of <0.5 mg L−1, NH4+ oxidation occurred at low potential rates (1.01 ± 0.05 mmol N m−2 d−1, p < 0.05) and respiratory ammonification processes also exhibited low potential rates (0.56 ± 0.36 mmol N m−2 d−1, p < 0.01). Therefore, at more anoxic sites, NO2– is mainly supplied by NO3– reduction (over 95.9%).

Biogeochemical mechanisms of anammox in the horizons of highest potential rate revealed by the interaction and estimated flux of different anammox-related N cycle processes in saturated soils, including a China-Anxin, b Australia-Queensland, c China-Antu and d Germany-Bremen, where DO levels were higher than 0.5 mg L−1, e Switzerland-Zurich, and f China-Wuqing, where DO levels were lower than 0.5 mg L−1. Denitrification (black line), anammox (red line), nitrite oxidation (purple line), and respiratory ammonification (green line) processes account for the NO2− sink. Nitrate reduction (blue line) and ammonium oxidation (pink line) were defined as the NO2− source (src). Respiratory ammonification processes compete with anammox and denitrification processes for NO2− and provide anammox with NH4+, acting as an important process coordinator. All potential rates were measured based on 15N isotopic pairing experiments and expressed as the mean value ± s.e. (standard error, n = 5)
Correlation analysis showed that the potential rate of anammox was positively correlated with both the potential rate of total N-loss (r = 0.935, p < 0.01) and the denitrification (r = 0.801, p < 0.05, Supplementary Table S12). In contrast, the potential rate of anammox was negatively correlated with DO concentrations (r = –0.612, p < 0.05, Supplementary Table S12). These findings indicate that the potential rates of anammox in aquifers are significantly affected by DO concentration and that the anammox biogeochemical mechanism and the related N-cycle processes are different, with respiratory ammonification processes acting as an important coordinator.
Discussion
Widespread anammox distribution contributes significantly to N-loss in marine ecosystems and has significantly shaped our view of the global N cycle. In contrast, anammox bacteria are distributed sporadically in terrestrial surface systems and are limited to inland waters and wetland soils [17, 23, 41,42,43], which cover less than 9% of the total global land area [44, 45]. These differences emphasize the need to further investigate the global distribution of anammox in terrestrial ecosystems. This study demonstrates the widespread global occurrence and significant impact of anammox on both N-loss and NO3− removal in aquifers, with anammox processes coupled to nitrification and respiratory ammonification processes. The coupled anammox- respiratory ammonification process has been reported to play a major role in N-loss in marine environments, such as the Eastern Tropical South Pacific [13] and the Arabian Sea [46, 47], where respiratory ammonification supplies NH4+ for anammox in NH4+ limited environments. The NH4+ substrate required for anammox bacteria in aquifer systems is further provided by respiratory ammonification processes, with the latter driven by high NO2− concentrations in turn produced by the higher potential rates of NH4+ oxidation. This mutual coupling of respiratory ammonification and NH4+ oxidation processes promotes the widespread distribution of anammox in global aquifer systems.
Potential rates can overestimate the actual in situ activity, as NH4+ and NO2− availability may limit in situ processes. Therefore, in the course of our experimental period, samples were incubated with NH4+ or NO2− at a final concentration corresponding to a max. of 10% of the measured in situ concentrations. Furthermore, anammox bacteria can mediate dissimilatory nitrate reduction to ammonium and use the generated NH4+ to supply anammox in NH4+-limited environments [33], which may result in the over- or underestimation of potential rates. In this study, the potential rates of anammox and respiratory ammonification were both determined by incubation treatment of slurries with 15NO2−. The substrate 15NH4+ produced from respiratory ammonification processes was utilized by anammox to generate 30N2 not 29N2, which may result in an underestimation of the anammox process, especially under the significant contribution by respiratory ammonification [33]. Therefore, serious overestimation of anammox in the soil core samples was prevented as far as possible in the present study. Based on the aquifer depth and soil density, an estimated 1.72–246 g-N m−2 yr−1 was lost via anammox processes in the saturated soils. These findings improve our overall understanding of N-cycling in aquifers and provide a feasible and effective way to control NO3− pollution in groundwater aquifers.
At some aquifer depths, anammox contributed higher than denitrification to N loss and NO3− removal. The spatial differences in the contribution of anammox can be attributed to the soil NO3− profile and level of the groundwater table. NO3− is the next preferred terminal electron acceptor after O2. Moreover, NO3− has good leaching capability and concentrations show a decreasing trend from the surface to lower depths in the soil profile, as it has the same electrical properties as soil (negatively charged). Previous studies have shown that water can activate anammox activity in dryland soils, with anammox activity being significantly higher at soil depths with higher NO3− concentrations [48]. Therefore, the trend in NO3− concentrations declined with greater depths in the soil profile and a higher potential rate of anammox was observed at the upper boundary of saturated soils, indicating a significant activity for the anammox process. Furthermore, these peak activity areas change according to the groundwater table, i.e. anammox would always occur and peak in the upper part of the saturated soil.
The varying contributions of anammox and denitrification processes to NO3− removal can be attributed to the difference in metabolic pathways of anammox and denitrifying bacteria. Denitrifiers are mostly facultative heterotrophs and therefore can obtain their energy from the oxidation of organic compounds and use organic carbon as an electron donor [49]. However, aquifers often contain low concentrations of dissolved organic compounds and have low levels of organic carbon, which is needed as an energy source and is usually identified as the major factor limiting denitrification [4]. The anammox process is driven by autotrophic bacteria, with energy supplied by N-compounds and carbon supplied by carbon dioxide (CO2) [50]. The water−land interface has been established as a hot spot for biogeochemical cycling and greenhouse gas CO2 emissions [51,52,53,54]. Furthermore, it has been widely established in the field of anammox bioreactor research that when organic compounds are limited, anammox bacteria are favored [17, 24, 37, 38, 49, 50].
Anammox and respiratory ammonification processes can occur under different oxygenic conditions, though they were usually considered to be significant only in anoxic/anaerobic ecotone environments. Over the last years, several studies reported respiratory ammonification processes under different oxygenic conditions including slurry, intact cores, and water columns with more than 80% of oxygen saturation levels [38, 55, 56]. As for anammox, our previous studies reported that anammox widely occurred in municipal wastewater treatment under aerobic conditions (DO > 5 mg L−1) [57]. In addition, studies also reported that anammox and respiratory ammonification processes could co-occur in the oxic compartment of riparian lake zones and constructed wetlands [58]. The oxygen tolerance of anammox and respiratory ammonification bacteria may both be attributed to their metabolic diversity. Analysis of the respiratory ammonification nrfA gene fragments in respective ecotones indicated that several aerobic microbes potentially performed respiratory ammonification processes [24]. Previous studies reported that Ca. Kuenenia adapt better to high DO (0–6.4 mg L−1) than Ca. B. fulgida (<0.032 mg L−1) and Ca. B. anammoxidans (2.24 mg L−1) [59].
In saturated soils, the potential rates of anammox and abundances were above the 0.5 nM N g−1 h−1 and 105 copies g−1 soil respectively, while in comparison the abundances and potential rates of anammox bacteria were undetectable in the unsaturated soils. Biogeochemical correlation analysis showed that the substrates NH4+ and NO2− had little positive influence on the potential rates of anammox. Therefore, substrate concentrations may not be the key limiting factor for the occurrence of anammox, while water contents showed a highly positive correlation with anammox occurrence. Previous studies have also demonstrated that anammox occurs ubiquitously in freshwater environments [17], with abundant anammox bacteria detected in soils of riparian zones after flooding and soils where no anammox bacteria were detected before flooding [17, 48, 60]. In terrestrial dryland soils, although anammox bacterial genes can be detected, anammox activity has not been reported to date [41]. However, higher abundance (up to 107 copies g−1) and potential rates (0.46 ± 0.08 nmol N g−1 h−1) of anammox have been detected in dryland soils with a high moisture content (>29%) [24]. Water can also reactivate anammox bacteria in dryland soils after long-term dormancy [48] and therefore, it may be inferred that water content is a key factor controlling anammox occurrence. Further investigations are required to establish whether water can be directly used by anammox bacteria or whether it induces the suitable anoxic−anaerobic environment for anammox bacteria.
This hypothesis is based on the special physiological properties of anammox bacteria, which have a very low half-saturation constant (Ks) for the substrates NH4+ (<5 μM) and NO2− (0.2–0.3 μM for Candidatus Kuenenia and <5 μM for Candidatus Brocadia). This may give anammox bacteria a selective advantage during competition for substrates, with either ammonium oxidizing bacteria (Ks of 0.14 mM NH4+ for Nitrospira sp. AV and 1.9 mM NH4+ for Nitrosomonas europaea) or nitrite oxidizing bacteria (Ks of 0.01 mM for Nitrospira and 0.02–0.14 mM for Nitrobacter) [61]. Furthermore, anammox bacteria could mediate dissimilatory nitrate reduction to ammonium and use the generated NH4+ and NO2− to fulfill anammox in NH4+-limited environments [46]. These features of anammox bacteria contribute to their widespread occurrence and abundance in various types of aquatic and terrestrial environments.
The presence of significant anammox activity in global aquifers demonstrates the ecological importance of this process in terrestrial ecosystems. Although the process of groundwater recharge and elevation of groundwater table is not a simple process, it stimulates and reactivates a variety of anammox bacteria and other microorganisms, accelerating biogeochemical processes. The theory that present climate change conditions may potentially initiate dormant bacteria reactivation on a global scale, which is presently not accounted for and its consequences on the nitrogen and carbon cycles, has not yet been explored.
In case of shortage and eutrophication of surface waters, groundwater has been a critically significant alternative source of potable drinking water, as well as for agricultural and industrial uses [51, 52]. However, groundwater resources are more vulnerable and are difficult to remediate once polluted, especially with respect to the current increase in NO3− contamination. This study provides a novel way to control NO3− pollution in groundwater environments. The human-induced environmental changes such as groundwater table control are expected to influence the relative contribution of subsurface N-loss processes, with a shift from denitrification to anammox and a potential reduction in N2O emissions.
References
Aeschbach-Hertig W, Gleeson T. Regional strategies for the accelerating global problem of groundwater depletion. Nat Geosci. 2012;5:853–61.
Taylor RG, Scanlon B, Döll P, Rodell M, van Beek R, Wada Y, et al. Ground water and climate change. Nat Clim Change. 2013;3:322–9.
Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA, Schindler DW, et al. Human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl. 1997;7:737–50.
Rivett MO, Buss SR, Morgan P, Smith JW, Bemment CD. Nitrate attenuation in groundwater: a review of biogeochemical controlling processes. Water Res. 2008;42:4215–32.
Schlesinger WH. On the fate of anthropogenic nitrogen. Proc Natl Acad Sci Usa. 2009;106:203–8.
WHO. Nitrate and nitrite in drinking-water. Background document for development of WHO Guidelines for Drinking-water Quality. Geneva: WHO; 2016.
Burgin AJ, Hamilton SK. Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Front Ecol Environ. 2007;5:89–96.
Strous M, Kuenen JG, Jetten MSM. Key physiology of anaerobic ammonium oxidation. Appl Environ Microbiol. 1999;65:3248–50.
Thamdrup B, Dalsgaard T. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments. Appl Environ Microbiol. 2002;68:1312–8.
Francis CA, Beman JM, Kuypers MM. New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation. ISME J. 2007;1:19–27.
Kartal B, Maalcke WJ, de Almeida NM, Cirpus I, Gloerich J, Geerts W, et al. Molecular mechanism of anaerobic ammonium oxidation. Nature. 2011;479:127–30.
Penton CR, Devol AH, Tiedje JM. Molecular evidence for the broad distribution of anaerobic ammonium-oxidizing bacteria in freshwater and marine sediments. Appl Environ Microbiol. 2006;72:6829–32.
Schubert CJ, Durisch-Kaiser E, Wehrli B, Thamdrup B, Lam P, Kuypers MM. Anaerobic ammonium oxidation in a tropical freshwater system (Lake Tanganyika). Environ Microbiol. 2006;8:1857–63.
Lam P, Gaute L, Marlene J, van de Jack V, Markus S, Dagmar W, et al. Revising the nitrogen cycle in the Peruvian oxygen minimum zone. Proc Natl Acad Sci Usa. 2009;106:4752–7.
Lam P, Kuypers MMM. Microbial nitrogen cycling processes in oxygen minimum zones. Ann Rev Mar Sci. 2011;3:317–45.
Devol AH. Denitrification, anammox, and N2 production in marine sediments. Annu Rev Mar Sci. 2015;7:403–23.
Zhu G, Wang S, Zhou L, Wang Y, Zhao S, Xia C, et al. Ubiquitous anaerobic ammonium oxidation in inland waters of China: an overlooked nitrous oxide mitigation process. Sci Rep. 2015;5:17306.
Lansdown K, McKew BA, Whitby C, Heppell CM, Dumbrell AJ, Binley A, et al. Importance and controls of anaerobic ammonium oxidation influenced by riverbed geology. Nat Geosci. 2016;9:357–60.
Kuypers MMM, Lavik G, Woebken D, Schmid M, Fuchs BM, Amann R, et al. Massive nitrogen loss from the Benguela upwelling system through anaerobic ammonium oxidation. Proc Natl Acad Sci USA. 2005;102:6478–83.
Zhu G, Wang S, Wang Y, Wang C, Risgaard-Petersen N, Jetten MS, et al. Anaerobic ammonia oxidation in a fertilized paddy soils. ISME J. 2011;5:1905–12.
Wang SY, Zhu GB, Peng YZ, Jetten MSM, Yin CQ. Anammox bacterial abundance, activity, and contribution in riparian sediments of the Pearl River estuary. Environ Sci Technol. 2012;46:8834–42.
Zhu G, Wang S, Wang W, Wang Y, Zhou L, Jiang B, et al. Hotspots of anaerobic ammonium oxidation at land-freshwater interfaces. Nat Geosci. 2013;6:103–7.
Nie S, Li H, Yang X, Zhang Z, Weng B, Huang F, et al. Nitrogen loss by anaerobic oxidation of ammonium in rice rhizosphere. ISME J. 2015;9:2059–67.
Zhu G, Wang S, Li Y, Zhuang L, Zhao S, Wang C, et al. Microbial pathways for nitrogen loss in an upland soil. Environ Microbiol. 2018;20:1723–38.
Schmidt SI, Hans JH. What is groundwater and what does this mean to fauna?—An opinion. Limnologica. 2012;42:1–6.
Moore TA, Xing Y, Lazenby B, Lynch MD, Schiff S, Robertson WD, et al. Prevalence of anaerobic ammonium-oxidizing bacteria in contaminated groundwater. Environ Sci Technol. 2011;45:7217–25.
Smith RL, Bohlke JK, Song B, Tobias CR. Role of anaerobic ammonium oxidation (anammox) in nitrogen removal from a freshwater aquifer. Environ Sci Technol. 2015;49:12169–77.
Granger J, Wankel SD. Isotopic overprinting of nitrification on denitrification as a ubiquitous and unifying feature of environmental nitrogen cycling. Proc Natl Acad Sci USA. 2016;113:203–8.
Dalsgaard T, Canfield DE, Petersen J, Thamdrup B, Acuña-González J. N2 production by the anammox reaction in the anoxic water column of Golfo Dulce, Costa Rica. Nature. 2003;422:606–8.
Risgaard-Petersen N, Nielsen LP, Rysgaard S, Dalsgaard T, Meyer RL. Application of the isotope pairing technique in sediments where anammox and denitrification co-exists. Limnol Oceanogr Meth. 2003;1:63–73.
Granger J, Sigman DM. Removal of nitrite with sulfamic acid for nitrate N and O isotope analysis with the denitrifier method. Rapid Commun Mass Sp. 2010;23:3753–62.
Füssel J, Lam P, Lavik G, Jensen MM, Holtappels M, Günter M, et al. Nitrite oxidation in the Namibian oxygen minimum zone. ISME J. 2012;6:1200–9.
Song GD, Liu SM, Kuypers MMM, Lavik G. Application of the isotope pairing technique in sediments where anammox, denitrification, and dissimilatory nitrate reduction to ammonium coexist. Limnol Oceanogr Meth. 2016;14:801–15.
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7:335–6.
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol. 2009;75:7537–41.
Deng Y, Jiang YH, Yang Y, He Z, Luo F, Zhou J. Molecular ecological network analyses. BMC Bioinf. 2012;13:113.
Rounds SA, Wilde FD, Ritz GF. Dissolved oxygen (ver. 3.0): U.S. Geological Survey Techniques of Water-Resources Investigations; Book 9, Chap. A6, Sec. 6.2, Virginia: U.S. Geological Survey, Reston, 2013.
Ali M, Oshiki M, Awata T, Isobe K, Kimura Z, Yoshikawa H, et al. Physiological characterization of anaerobic ammonium oxidizing bacterium “Candidatus Jettenia caeni”. Environ Microbiol. 2014;17:2172–89.
Roberts KL, Kessler AJ, Grace MR, Cook PL. Increased activity of dissimilatory nitrate reduction to ammonium (DNRA) under oxic conditions in a periodically hypoxic estuary. Geochim Cosmochim Acta. 2014;133:313–24.
van den Berg EM, van Dongen U, Abbas B, van Loosdrecht MC. Enrichment of DNRA bacteria in a continuous culture. ISME J. 2015;9:2153–61.
Shan J, Zhao X, Sheng R, Xia Y, Ti C, Quan X, et al. Dissimilatory nitrate reduction processes in typical Chinese paddy soils: rates, relative contributions and influencing factors. Environ Sci Technol. 2016;50:9972–80.
Humbert S, Tarnawski S, Fromin N, Mallet MP, Aragno M, Zopfi J. Molecular detection of anammox bacteria in terrestrial ecosystems: distribution and diversity. ISME J. 2010;4:450–4.
Yang XR, Li H, Nie SA, Su JQ, Weng BS, Zhu GB, et al. Potential contribution of anammox to nitrogen loss from paddy soils in Southern China. Appl Environ Microbiol. 2015;81:938–47.
Zedler JB, Kercher S. Wetland resources: status, trends, ecosystem services, and restorability. Environ Resour. 2005;15:39–74.
Mitsch WJ. Applying science to conservation and restoration of the world’s wetlands. Water Sci Technol. 2005;51:13–26.
Jensen MM, Lam P, Revsbech NP, Nagel B, Gaye B, Jetten MSM, et al. Intensive nitrogen loss over the omani shelf due to anammox coupled with dissimilatory nitrite reduction to ammonium. ISME J. 2011;5:1660–70.
Kartal B, Kuypers MMM, Lavik G, Schalk J, Op den Camp HJM, Jetten MSM, et al. Anammox bacteria disguised as denitrifiers: nitrate reduction to dinitrogen gas via nitrite and ammonium. Environ Microbiol. 2007;9:635–42.
Zhu G, Wang S, Wang C, Zhou L, Zhao S, Li Y, et al. Resuscitation of anammox bacteria after >10,000 years of dormancy. ISME J. 2019;13:1098–109.
Zhu G, Peng Y, Li B, Guo J, Wang S. Biological removal of nitrogen from wastewater. Rev Environ Contam Toxicol. 2008;192:159–95.
Jetten MSM. The microbial nitrogen cycle. Environ Microbiol. 2008;10:2903–9.
Naiman RJ, Décamps H. The ecology of interfaces: Riparian zones. Annu Rev Ecol Syst. 1997;28:621–58.
McClain ME, Boyer EW, Dent CL, Gergel SE, Grimm NB, Groffman PM, et al. Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems. 2003;6:301–12.
Verhoeven JTA, Arheimer B, Yin C, Hefting MM. Regional and global concerns over wetlands and water quality. Geochim Cosmochim Acta. 2006;21:0–103.
Wang H, Lu J, Wang W, Yang L, Yin C. Methane fluxes from the littoral zone of hypereutrophic Taihu lake, china. Geochim Cosmochim Acta. 2006;111:1–8.
Song B, Lisa JA, Tobias CR. Linking DNRA community structure and activity in a shallow lagoonal estuarine system. Front Microbiol. 2014;5:1–10.
Bonaglia S, Klawonn I, De Brabandere L, Deutsch B, Thamdrup B, Brüchert V. Denitrification and DNRA at the Baltic Sea oxic–anoxic interface: substrate spectrum and kinetics. Limnol Oceanogr. 2016;61:1900–15.
Wang SY, Peng YZ, Ma B, Wang SY, Zhu GB. Anaerobic ammonium oxidation in traditional municipal wastewater treatment plants with low-strength ammonium loading: widespread but overlooked. Water Res. 2015;84:66–75.
Wang SY, Wang WD, Liu L, Zhuang LJ, Zhao SY, Su Y, et al. Microbial nitrogen cycle hotspots in the plant-bed/ditch system of a constructed wetland with N2O mitigation. Environ Sci Technol. 2018;52:6226–36.
Oshiki M, Satoh H, Okabe S. Ecology and physiology of anaerobic ammonium oxidizing bacteria. Environ Microbiol. 2016;18:2784–96.
Hu BL, Shen LD, Liu S, Cai C, Chen TT, Kartal B, et al. Enrichment of an anammox bacterial community from a flooded paddy soil. Environ Microbiol Rep. 2013;5:483–9.
Strous M, Fuerst JA, Kramer EH, Logemann S, Muyzer G, van de Pas-Schoonen KT, et al. Missing lithotroph identified as new planctomycete. Nature. 1999;400:446–9.
Acknowledgements
The authors thank Mr. Dachun Zhu for sampling the soil cores in Jilin and Antu (farmland and forest), Dr. Shuangbing Huang for providing the soil core samples in Wuqing, and Dr. Shuci Liu for his help on model analysis. This research was financially supported by the National Natural Science Foundation of China (Nos. 41322012, 41671471, and 91851204), Strategic Priority Research Program of the Chinese Academy of Sciences (XDB15020303), Local Innovative and Research Teams Project of Guangdong Pearl River Talents Program (2017BT01Z176), National Key R&D Program (2016YFA0602303), and a special fund from the Key Laboratory of Drinking Water Science and Technology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences (15Z07KLDWST and 16Z03KLDWST) as well as the Max Planck Society (MPG). GZ gratefully acknowledges the Program of the Youth Innovation Promotion Association (CAS), and the support of a Humboldt Research Fellowship (1152633). MSMJ was supported by erc ag 232937 anammox, erc ag ecomom 339880 and siam 024002002. LS acknowledges support by the German Research Foundation (Schw554/25).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
About this article

Cite this article
Wang, S., Zhu, G., Zhuang, L. et al. Anaerobic ammonium oxidation is a major N-sink in aquifer systems around the world. ISME J 14, 151–163 (2020). https://doi.org/10.1038/s41396-019-0513-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41396-019-0513-x
