
Abstract
Fibrosis is characterized by the excessive extracellular matrix deposition due to dysregulated wound and connective tissue repair response. Multiple organs can develop fibrosis, including the liver, kidney, heart, and lung. Fibrosis such as liver cirrhosis, idiopathic pulmonary fibrosis, and cystic fibrosis caused substantial disease burden. Persistent abnormal activation of myofibroblasts mediated by various signals, such as transforming growth factor, platelet-derived growth factor, and fibroblast growh factor, has been recongized as a major event in the occurrence and progression of fibrosis. Although the mechanisms driving organ-specific fibrosis have not been fully elucidated, drugs targeting these identified aberrant signals have achieved potent anti-fibrotic efficacy in clinical trials. In this review, we briefly introduce the aetiology and epidemiology of several fibrosis diseases, including liver fibrosis, kidney fibrosis, cardiac fibrosis, and pulmonary fibrosis. Then, we summarise the abnormal cells (epithelial cells, endothelial cells, immune cells, and fibroblasts) and their interactions in fibrosis. In addition, we also focus on the aberrant signaling pathways and therapeutic targets that regulate myofibroblast activation, extracellular matrix cross-linking, metabolism, and inflammation in fibrosis. Finally, we discuss the anti-fibrotic drugs based on their targets and clinical trials. This review provides reference for further research on fibrosis mechanism, drug development, and clinical trials.
Similar content being viewed by others
Introduction
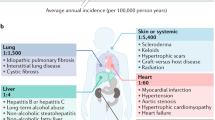
Fibrosis is an important cause of global morbidity and mortality. Common diseases associated with fibrosis include hepatitis virus, nonalcoholic fatty liver disease (NAFLD), chronic kidney diseases, idiopathic pulmonary fibrosis (IPF), pneumonconiosis, and cystic fibrosis. The annual combined incidence of major fibrosis-related diseases is approximately 4968 per 100,000 person-years, causing huge disease burden1. Fibrosis-related diseases accounted for a large proportion of global disability-adjusted life-years (DALYs) in 20192. Therefore, fibrosis is increasingly recognized as a major health challenge.
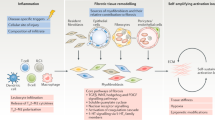
The normal wound healing process and the pathogenesis of fibrotic diseases share many mechanisms in common3. Various factors, such as infectious agents, alcohol, environmental particles, and gene mutation, can cause damage to normal tissue structures, triggering a wound-healing response4. The tissue repair response often starts with inflammation. Activated inflammation contributes to the upregulation of inflammatory mediators and promotes the migration of neutrophils, eosinophils, and macrophages to the injured site to clear debris and necrotic areas. Fibroblasts and other mesenchymel cells are then thansformed to myofibroblasts via the upregulation of fibrotic cytokines such as fibroblast growth factors (FGFs) and platelet-derived growth factor (PDGFs), which secrete extracellular matrix (ECM) components5. In normal wound healing response, activated myofibroblasts would be cleared from wound site via apoptosis after injury repair6,7. However, in fibrotic process, myofibroblasts fail to undergo apoptosis and are continuously activated, eventually leading to excessive ECM deposition8. The progressive accumulation of ECM leads to increased stiffness of injured tissue and hinders oxygen diffusion9, and further promotes cell damage. In addition, dysfunction of other parenchymal cells and dysregulated cell-cell interaction caused by injury are also the important causes of fibrosis, such as vascular proliferation induced by abnormal function of vascular endothelial cells10. The fibrotic process can occur in many organs, with fibrosis of liver, lung, kidney, and heart accounting for a large proportion of all fibrotic diseases1,11. The different characteristics of tissue structure and microenvironment between these organs lead to differences in the fibrotic process (Fig. 1). Despite increasing in-depth research on fibrosis, the mechanisms have not been fully explained, thus hindering the advancement of targeted drug research for fibrosis.

The aetiology of fibrosis in different tissues or organs
In this review, we briefly introduce the aetiology and epidemiology of several fibrosis-related diseases, including liver fibrosis, renal fibrosis, heart fibrosis, lung fibrosis, cystic fibrosis, and myelofibrosis. We then focus on the abnormal cells, aberrant signaling pathways, and anti-fibrotic drugs in fibrosis, providing reference for the mechanism and drugs research of fibrosis.
Aetiology and epidemiology
Liver fibrosis
Liver fibrosis, as a pathophysiological result of chronic liver injury, is the leading cause of mortality from chronic liver diseases (CLDs) worldwide. CLDs mainly include chronic infection with hepatitis virus, NAFLD, alcoholic liver diseases, and autoimmune liver diseases12,13. CLDs could progress to advanced liver fibrosis and eventually to cirrhosis14, which is the 11th cause of global death15. Hepatitis B virus (HBV), hepatitis C virus (HCV), and alcohol are the most common causes of DALYs from cirrhosis16. Alcoholic-related liver cirrhosis and other chronic liver diseases resulted in 332,300 all-age deaths and 9,785,400 years of life lost (YLLs) in 201717. The prevalence of NAFLD is around 25% worldwide18, and its advance can progress to nonalcoholic steatohepatitis (NASH)19. NASH-related cirrhosis caused 118,000 all-age deaths and 3,285,500 YLLs in 201717.
Renal fibrosis
Renal fibrosis is caused by the damage to normal renal tubules, which eventually leads to glomerulosclerosis, tubulointerstitial fibrosis, and angiosclerosis20. Renal fibrosis is not a direct clinical diagnosis but a progressive and irreversible pathological feature of all chronic kidney diseases (CKDs)21,22. In 2017, CKDs caused 35.8 million DALYs, nearly a third of which were diabetic nephropathy23.
Cardiac fibrosis
Cardiac fibrosis manifests as either reactive interstitial fibrosis and replacement fibrosis24. Reactive interstitial fibrosis refers to the expansion of interstitial and perivascular spaces without significant loss of cardiomyocytes and fundamental changes in muscle bundle structure25. Replacement fibrosis replaces dead cardiomyocytes with extracellular matrix tissue and fibroblasts, disrupting the continuous pattern of muscle bundles but maintaining tissue integrity26. Replacement fibrosis mainly occurs in response to ischaemia, ischaemia/reperfusion, inflammation, and toxic injury. Cardiac fibrosis is a common pathophysiological manifestation of most cardiovascular diseases, which are the leading cause of death, morbidity, and disability in most contries27,28.
Lung fibrosis
The causes of chronic respiratory diseases are varied, including allergens, chemicals, radiation, microbial agents, and environmental particles29. Lung fibrosis is the main clinical outcome of most chronic respiratory diseases, such as pneumoconiosis and IPF30. IPF is the most common interstitial lung fibrosis with unknown aetiology31,32. The prevalence of IPF varies widely across regions, ranging from 0.33 to 2.51 in Europe, 0.57 to 4.51 in Asia-Pacific countries, and 2.40 to 2.98 in North America33. IPF mainly occurs in elderly individuals, with high mortality and morbidity34,35. Pneumoconiosis is a major occupational diseases caused by the prolonged inhalation of inorganic particles at work36,37,38. In 2017, all-age deaths of pneumoconiosis was 21,600 and 426,900 YLLs17.
Cystic fibrosis
Cystic fibrosis is an autosomal recessive disorder mainly caused by mutations in the cystic fibrosis transmembrane conductance regulatory protein (CFTR) gene39. Compared with the high incidence rate of cystic fibrosis in Caucasians, cystic fibrosis was much less common in Asia, and the incidence rate varied from 1:10,000 to 1:40,750 among countries40,41,42.
Myelofibrosis
Myelofibrosis, a myeloproliferative tumour with collagen deposition in bone marrow and splenomegaly, has low morbidity and shortened life expectancy43,44,45. Aberrant activity of the Janus kinase (JAK) /signal transducer and activator of transcription (STAT) pathway contributes to myelofibrosis43,46.
Abnormal cells involved in fibrosis
Fibrosis is the result of the interaction between a variety of cells. Cell maps of fibrosis such as IPF, liver fibrosis, renal fibrosis, and systemic sclerosis have been well studied via single-cell sequencing47,48,49,50. These studies confirmed the key role of epithelial cells, endotheliocytes, immunocytes, and fibroblasts in fibrosis, and identified some new cell types involved in the pathological progress. This section will review the major cell types in fibrotic diseases.
Epithelial cells
Epithelial cells, including basal cells, secretory cells, club cells, ciliated cells, and goblet cells, are essential cells to maintain tissue homeostasis in many organs51. In fibrotic process, chronic injury resulted in the apoptosis of epithelial cells, thus destroying the epithelial structure, promoting dysfunctional repair and pathogenic activation of fibroblasts52. Moreover, the epithelial-mesenchymal transition (EMT) is recognized as an important source of myofibroblasts. EMT under pathological conditions can lead to the reduction of normal epithelial cells, destroy the normal structure of the tissue, and promote the production of collagen fibers53.
Studies have showed that epithelial cells, such as alveolar epithelial cells, goblet cells, ciliated cells, and club cells, are crucial for the development of lung fibrosis54,55. Alveolar epithelial cells, including alveolar type 1 epithelial (AT1) and AT2 cells, are one of the main epithelial cells in lung tissue and maintain the integrity of the alveolar wall. When the injury leads to the death of AT1 cells, AT2 cells proliferate and differentiate into AT1 cells, so that the normal structural of the alveoli is maintained56. A new epithelial cell subset Axin2+ AT2 cells with both progenitor and epithelial properties was found in lung and regulate alveolar regeneration57,58. AT2-transdifferentiated plastic keratin 5 basal cells were co-located with pathological transforming growth factor (TGF) -β1hi collagen triple helix repeat containing 1 (CTHRC1)hi fibroblasts and have a synergistic effect in the progress of fibrosis59.
A new group of epithelial cells with high expression of CFTR, named ionocytes, was found in airway epithelium60. One of the most important functions of CFTR is to regulate chloride channels61. Therefore, the mutations of CFTR gene of epithelial cells results in chloride channel defects in airway epithelium, initiating the occurrence of cystic fibrosis62. Moreover, the lack of CFTR in airway increased Na+ channel activity and Na+ hyperabsorption, suggesting that CFTR might be involved in Na+ transport61. The functional change of epithelial cells in the pancreas and liver is also affected by CFTR mutation63. In the normal liver, CFTR cooperates with the chloride channel at the top of cholangiocytes to provide a driving force for bile hydration64. Impaired CFTR function lead to mucosal hyperplasia and obstruction of the bile duct. Subsequent bile salt accumulation contributed to hepatocyte damage, inflammation, and fibrosis in the portal vein64,65.
Endothelial cell
Endothelial cells are main components of blood vessels. Damage to endothelial cells cause abnormal substances exchange between blood and tissues, resulting in metabolic disorders. Furthermore, in fibrotic tissues, abnormal angiogenesis may be induced due to the massive proliferation of fibroblasts requiring more blood nutrients. Studies showed that endothelial cells of different fibrotic tissues may also have specific functions. Two new endothelial cell subtypes, plasmalemma vesicle associated protein (PLVAP)+ endothelial cells and atypical chemokine receptor 1 (ACKR1)+ endothelial cells, were found in liver tissues of patients with liver cirrhosis and could promote the migration of leukocyte48. In lung tissues, five endothelial cell groups were identified by single-cell sequencing, including capillary endothelial cells A and B, venous endothelial cells, and arterial endothelial cells. The fifth kind of endothelial cells recognized by high expression of Collagen 15a1 (COL15A1) gene, located in the bronchioles and fibrous foci, was involved in the production of extracellular matrix47.
Immune cells
Abnormality of immune system might be an early event of fibrosis66. Immunocytes, such as T lymphocytes, macrophages, dendritic cells, granulocytes, and mast cells, are involved in the fibrosis progress49,67,68,69,70. These activated immune cells highly express factors that regulate inflammation and fibrosis, promoting the activation of fibroblasts. T lymphocytes, including CD4+T cells, CD8+T cells, and CD8+effector cells, were increased in IPF patients71. The interferon-γ signal transduction in T lymphocytes in IPF was significantly changed71, while interleukin (IL) -6 signal in T lymphocytes was mainly up-regulated in patients with systemic sclerosis67. In liver tissues, the expression of cytotoxic T cells increased and the inactivation of CD4+ T cells could induce fibrosis72.
Macrophages are key cells that mediate inflammation and fibrosis in fibrotic diseases. Seven macrophage subsets were identified in the tissues of patients with liver cirrhosis, including Kupffer cells (resident macrophages in liver) and CD9+ triggering receptor expressed on myeloid cells 2 (TREM2)+ macrophages. Pseudo-time sequence analysis showed that TREM2+CD9+ macrophages were derived from monocytes and increased collagen expression in hepatic stellate cells (HSCs)48. In the lung fibrosis, 18 types of immune cells were found, and the phenotypes of tissue resident macrophages, fibrogenic macrophages and inflammatory macrophages were identified47,54. Resident macrophages in lung are mainly alveolar macrophages (AMs). AMs adheres closely to alveolar epithelium and are exposed to the outside environment73. Inhalable particles and other factors directly led to the death of AMs74. Activated AMs secreted inflammatory mediators to activate the inflammatory response, and elevated pro-fibrotic factors expression to promote lung fibrosis75,76. The sialic acid binding Ig-like lectin F (SiglecF)+ C-X3-C motif chemokine receptor 1 (CX3CR1)+ macrophages were also identified in pulmonary fibrosis mouse model, which were adjacent to fibroblasts and promoted fibrosis by releasing PDGFs to drive the proliferation and activation of fibroblasts77.
Fibroblasts
Differentiation of fibroblasts to myofibroblasts with secretory, contractile, and extracellular matrix-producing properties is a key cellular event in many fibrotic conditions. Single-cell sequencing has demonstrated that myofibroblasts have different gene expression profiles with dynamic changes in fibrosis of different organs78,79. In lung tissue, the differentiation pathways of fibroblasts differ between normal and fibrotic pathological states. Mesenchymal progenitor cells differentiate into lipofibroblasts and COL14A1+ matrix fibroblasts, and the latter then differentiate into myofibroblasts and COL13A1+ matrix fibroblasts. In lung fibrosis, mesenchymal progenitors differentiate into lipofibroblasts, PDGFRβhi subtypes, COL14A1+ matrix fibroblasts, myofibroblasts, and COL13A1+ matrix fibroblasts80. The dominant cell type of fibroblasts in liver are HSCs, which are characterized by their star-like morphology. The differentiation of HSCs may undergo four processes: loss of quiescent properties, promoting inflammation, migration, and ECM production50.
Increasing number and activation of myofibroblasts induced by immune cells, EMT, and endothelial-mesenchymal transition (EndMT) are considered major contributors to the process of fibrogenesis81,82. Inhibiting the proliferation and activation of myofibroblasts has been a critical issue for the treatment of most fibrosis. However, in the fibrotic process, myofibroblast cells could obtain apoptosis resistance during differentiation83, which hinders the implementation of programmed death mechanisms8. Therefore, the therapeutic method for reducing the number of myofibroblasts has limited efficacy. Moreover, the hyper-activation of myofibroblasts is usually a compensatory result of the death of parenchymal cells such as epithelial cells, cardiomyocytes, and endotheliocytes. Therefore, it might be a more effective treatment method to decrease the death or modulate the activity of parenchymal cells and other related cells, so as to indirectly inhibit myofibroblast activation.
In liver fibrosis, the interaction mechanism of HSCs with other cells is complex. Maintenance of liver sinusoidal endothelial cells (LSECs) differentiation leads to HSCs quiescence and fibrosis regression in normal liver84,85. However, in fibrotic process, apoptotic hepatocytes increase the inflammatory response and activate macrophages86. Extracellular events from Kupffer cells (liver-resident macrophages), hepatocytes, B lymphocytes, and T lymphocytes further modulate the activation of HSCs87,88. NK cells could kill activated HSCs via regulating retinoic acid-induced 1/natural killer group 2D (NKG2D) -dependent and TNF-related apoptosis-inducing ligands89,90. Chronic liver injury leads to continuous HSCs activation, which promotes ECM accumulation and tissue structure remodeling, and then results in progressive liver fibrosis91 (Fig. 2).

The activation of HSCs regulated by other cells in liver fibrosis. Extracellular components from injured hepatocytes, Kupffer cells, macrophages, NK cells, T and B lymphocytes modulate HSCs activation via various cytokines. LSECs inhibit or promote the activation of HSCs in different conditions. NK cells kill activated HSCs in IFNγ and TRAIL-dependent ways. TRAIL, TNF-related apoptosis-inducing ligand
In the lung, acute injury of alveolar epithelial cells can cause the reduction of epithelial cells, the destruction of alveolar structure, and the release of pro-inflammatory mediators, thus activating immune cells. These activated inflammatory cells and injured epithelial cells increase the upregulation of cytokines, including TNF-α, IL-1β, IL-6, and TGF-β92,93,94. After the initial inflammatory events, pulmonary fibroblasts are activated into myofibroblasts by upregulating fibrotic cytokines such as PDGFs, FGFs, and vascular endothelial growth factor (VEGFs)95,96,97,98. The transition of epithelial cells by the EMT process could also increase the population of myofibroblasts. Chronic activated myofibroblasts produce ECM components (collagens, fibronectin, proteoglycan), leading to lung fibrosis12,22,99 (Fig. 3).

The interactions among cells involved in lung fibrosis. Injured alveolar epithelial cells activate macrophages, neutrophils, and eosinophils, resulting in the secretion of cytokines, such as TGF-β, IL-1β, and TNF-α. These cytokines mediate the differentiation of fibroblasts into myofibroblasts and the epithelial-mesenchymal transition, which result in the ECM deposition at the injury site
Important signaling pathways in fibrosis
An overwhelming number of mediators have been implicated in fibrosis, regulating myofibroblast activation, metabolism, inflammation, and ECM cross-linking. This part mainly focus on the important signaling pathways involved in fibrotic diseases based on the research intensity and drug efficacy of drug targets in clinical trials.
Growth factors and associated signaling pathways
The growth factors and associated signaling pathways have been reported to promote fibrosis by regulating fibroblasts activation, epithelial cells apoptosis, EMT, and EndMT. Growth factors mainly include TGF-βs, PDGFs, FGFs, and connective tissue growth factor (CTGF). Pathways, such as phosphatidylinositol 3-kinase (PI3K) / protein kinase B (AKT), JAK/STAT, and WNT/β-catenin, are the common downstream signals of these growth factors involved in fibrosis. The interactions between these signaling pathways in fibrosis are depicted in Fig. 4.

Interactions between growth factors-associated signaling pathways and a summary of related target drugs. PDGFs binding to PDGFRs activates the JAK/STAT, PI3K/AKT, and RAS/ERK signals. FGFs binding to FGFRs activates PI3K/AKT and RAS/ERK signals. CTGF binding to FGFR2 (promoting FGF2 and FGF4 binding to FGFR2) activates RAS/ERK signaling, and CTGF binding to LRP6 activates WNT/β-catenin signaling. Drugs targeting these signaling pathways are listed. EMT: epithelial-mesenchymal transition
TGF-β signaling pathway
TGF-β activation
TGF-βs are the key cytokines in most fibrosis. There are three isoforms of TGF-βs, namely, TGF-β1, TGF-β2, and TGF-β3. The pro-TGF-β monomer synthesized in ribosome, folds in the lumen of the endoplasmic reticulum (ER) and dimerizes via a disulfide linkage. Then, the latency-associated peptide (LAP) binds to mature TGF-β and attaches to latent TGF-β binding protein (LTBP)100. This TGF-β/LAP/LTBP complex binds to the ECM in the extracellular space and inactivates TGF-β101. The complex can be cleaved by various proteases to release active TGF-β102. Activated TGF-βs bind to TGFβR2 and TGFβR1100. Upon ligand binding, phosphorylated TGFβR2 then phosphorylates and activates TGFβR1. Factors, such as epidermal growth factor (EGF), IL-1, and TNF-α promote TGF‑β expression in different types of cells103,104. Moreover, the precursors of TGF-β contain an arginine-glycine-aspartate (RGD) motif, which can be recognized by integrin αv/β6105,106, suggesting that the activation of TGF-β gene could be regulated by integrin αv/β6. Partial inhibition of TGF-β with an integrin αv/β6 antibody effectively prevented pulmonary fibrosis in mice without aggravating inflammation107,108.
Canonical and non-canonical signaling
TGF-βs can regulate fibrosis via both canonical and non-canonical signaling pathways. Smad proteins are the canonical intracellular effector of TGF-β/TGFβR. Activated TGFβR1 subsequently induces phosphorylation of Smad2 and Smad3, which interact with Smad4 and enter the nucleus to activate the expression of target genes102. Smad7 is a negative regulator of TGF‑β/Smad signaling109 (Fig. 5). TGF-β could also activate non-canonical (non-Smad) signaling pathways, such as PI3K/AKT, mitogen-activated protein kinase (MAPK) pathways, and JAK/ STAT110. Macrophages, epithelial cells, and fibroblasts were the main sources of TGF-β in fibrosis111,112. TGF-β promotes fibrosis through diverse mechanisms, including activation of resident fibroblasts, promotion of cell apoptosis, and induction of EMT.

Overview of canonical TGF-β/Smad signaling pathway. Various cytokines stimulate the transcipiton of TGF-β, such as PDGFs, TGF-βs, TNF-α, IL-1β, and EGF. Pro-TGF-β is synthesized in the ribosome and endoplasmic reticulum. After dimeration, LAP binds to mature TGF-β and attaches to LTBP, entering the intercellular space through exocytosis. Actived TGF-β is released by proteases, and binds to TGFβR2 and TGFβR1. Phosphorylated TGFβR2 phosphorylates TGFβR1. TGFβR1 subsequently triggers the phosphorylation of Smad2/3, which interact with Smad4 and enter the nucleus to activate the expression of target genes. Smad7 is a negative regulator of TGF-β/Smad signaling. LAP, latency-associated peptide; LTBP, latent TGF-β binding protein
Fibroblast activation induced by TGF-β
Activated TGF-β1/Smad3 signaling pathway promoted the recruitment of fibroblasts to injury sites and mediated fibroblast-to-myofibroblast differentiation, thus stimulating the secretion of ECM components113,114,115. Reactive oxygen species (ROS) has been reported to mediate TGF-β-induced activation of fibroblasts. NADPH oxidase (Nox) enzymes are important mediators of electron transport from NADPH to oxygen to form ROS116. Once produced, ROS could induce the activation of TGF-β1. Nox4 is a member of Nox enzyme family and its expression could be induced by TGF-β in a variety of cells117. TGF-β1 treatment increased the level of Nox4 and alpha-smooth muscle actin (α-SMA), a myofibroblast marker, in primary human cardiac fibroblasts, whereas depletion of Nox4 decreased TGF-β1-stimulated α-SMA expression, indicating that ROS mediated TGF-β1-induced activation of cardiac fibroblasts to myofibroblasts118. Recent studies have suggested that TGF-β1-driven activation of fibroblasts might involve metabolic reprogramming in fibroblasts and enhancement of glycolytic pathways119.
Cell apoptosis induced by TGF-β
TGF-β1-induced apoptosis is important in various fibrosis and the mechanisms might differ between different cell types. ROS plays a key role in endothelial cell apoptosis induced by TGF-β. TGF-β1 caused ROS-dependent p38 activation, while p38 inhibition decrased TGF-β1-induced apoptosis120. TGF-β1 could also induce apoptosis of mesangial cells in kidney via p53 phosphorylation and Bcl-2 Associated protein X (Bax) up-regulation121.
EMT regulated by TGF-β
In fibrosis, the most common type of EMT is the type 2 EMT process. Type 2 EMT, mainly caused by inflammation, is closely related to tissue damage repair response and increases myofibroblasts population122. TGF-β is a crucial mediator in regulating type 2 EMT process in fibrosis and its interaction with various signals regulates the occurrence of EMT. Oxidative stress induced by TGF-β is an important event in the EMT process. TGF-β increased the level of ROS by upregulating the expression of Nox4, and then activated ERK and mTOR signaling molecules to promote EMT and fibrosis123. PI3K/AKT signals also mediated TGF-β-induced EMT124.
PDGFs/PDGFRs
PDGFs are stimulators of cell division that are required for cell growth and proliferation. They are disulfide-bonded homodimers and heterodimers composed of five different polypeptide chains (subunits), termed AA, AB, BB, CC, and DD125. PDGF ligands bind to PDGFRαα, PDGFRαβ and PDGFRββ126. PDGF-A and -C subunits mainly bind to the α chain, B subunit to both α and β chains, and D subunit to the β chain only127. Upon ligand binding, PDGFRs phosphorylate and activate downstream signals (RAS/MAPK, PI3K/AKT, and JAK/STAT pathways)128.
PDGFs are increased in fibrosis. Macrophages, endothelial cells, and fibroblasts have been identified as the main sources of PDGFs129,130,131,132. Both PDGF-B and PDGF-D were potent factors for HSCs proliferation and migration, therefore potentiating extracellular matrix deposition in liver fibrogenesis133,134, which could be mediated by PGDFRβ135. However, deficiency of PDGF-C failed to inhibit liver fibrosis or functional liver impairment136, but alleviated kidney fibrotic changes in experimental murine kidney fibrosis137. In addition to kidney and liver, studies demonstrated that PDGFs contributed to the formation of heart and lung fibrosis via stimulating activation of fibroblasts138,139,140.
FGFs/FGFRs
There are 18 members of the FGF superfamily, which are divided into 6 groups according to sequence homology and differences in biological properties: aFGF and bFGF; INT2, KGF, FGF10, and FGF22; FGF4, FGF5, and FGF6; FGF8, FGF17, and FGF18; FGF9, FGF16, and FGF20; FGF19, FGF21, and FGF23141. FGF receptors (FGFR1-FGFR4) are mainly composed of a transmembrane domain, a cytoplasmic tyrosine kinase domain, and an extracellular immunoglobulin domain (D1-D3)142. FGFs induce the dimerization, activation, and autophosphorylation of FGFRs and activate the RAS-extracellular signal-regulated kinase (ERK), PI3K-AKT, and JAK/STAT pathways143,144,145. The role of FGFs family in liver fibrosis is not clear. FGF19 deficiency protected mice from liver fibrosis progress in animal models146. However, direct stimulation of FGF19 decreased pro-fibrotic and pro-inflammatory cytokines expression on HSCs147. FGF21 has attracted much attention due to its important role in liver lipid metabolism148,149. FGF21 acts in an endocrine, paracrine, and autocrine-like manner via FGFR1-3/β-Klotho (KLB)150. FGF21-knockout mice decreased β oxidation and increased the level of free fatty acids in mice fed methionine- and choline-deficient (MCD) diets, promoting lipotoxicity and steatosis151. Increasing expression of FGF21 inhibited inflammation in NASH, and synergistically alleviated obesity and insulin resistance151,152. For pulmonary fibrosis, the FGF family is a therapeutic target that promotes fibroblast proliferation and migration but inhibits myofibroblast differentiation153,154,155,156. Inhibition of FGF/FGFR signaling has achieved reduction of pulmonary fibrosis in IPF157.
VEGFs/VEGFRs
The VEGF family has 6 members: VEGF-A, -B, -C, -D, -E, and placental growth factor (PIGF)158. VEGFs, which are similar to PDGF family proteins in structure, regulates vasculogenesis, angiogenesis and immunity159. VEGF-A is widely studied in regulating angiogenesis during homeostasis and disease160. VEGF-A exerts its biological functions by binding to VEGFR1 and VEGFR2160. VEGF-A were decreased in IPF patients, and lung-specific overexpression of VEGF-A attenuated the lung injury and fibrosis in lung fibrosis mouse model161. However, studies have shown the important role of VEGF in promoting pulmonary fibrosis162,163. The selective splicing of exons contributes to the existence of various subtypes of VEGF-A, including VEGF-A121, VEGF-A165, VEGF-A189, and VEGF-A206, among which VEGF-A165 is the most abundant isoform in normal tissues164,165. Most studies on the role of VEGF-A in fibrosis have not clearly identified the subtype of VEGF-A, and the dual role of VEGF-A in fibrosis might be related to its different subtypes166.
CTGF signaling pathway
CTGF is a secreted peptide and has been considered as a novel PDGF-related growth factor regulating the proliferation and chemotaxis of fibroblasts167. CTGF can combine with other molecules to promote their pro-fibrotic effects, thereby promoting fibrosis. The binding of CTGF with FGFR2 enhanced the binding of FGFR to FGF2 and FGF4, thus activating ERK signaling and promoting proliferation168. Additional studies have shown that CTGF could bind to TGF-β1169 and was required for the pro-fibrotic activity of TGF-β1170,171. TGF-β-induced endogenous CTGF leads to transcriptional repression of Smad7 via inducing the transcription factor TIEG-1, and by this mechanism, CTGF blocks the inhibitory effect of Smad7, resulting in persistent activation of TGF-β signaling172.
PI3K/AKT
PI3Ks can be activated by receptor-coupled tyrosine kinase activity, small RAS-related GTPases, and heterotrimeric G proteins173. The common downstream of receptor-mediated PI3K activation is AKT, which can phosphorylate many substrates related to cell proliferation, autophagy, and motility173. Activated PI3K/AKT negatively regulates the activity of mammalian target of rapamycin (mTOR)174. The PI3K/AKT/mTOR is a pivotal signaling involved in cell proliferation and differentiation175, and was activated in fibrotic foci176,177. The activated PI3K/AKT participated in the TGF-β-induced myofibroblasts activation178. PI3K/AKT could also regulate angiogenesis by increasing VEGF/VEGFR signaling179 and enhanced VEGFA/VEGFR2 signaling in liver fibrosis and angiogenesis180,181.
JAK/STAT
The JAKs has four members, JAK1, 2, 3, and TYK2182. Upon ligand binding, JAKs are activated and subsequently phosphorylate downstream signaling molecules, such as STAT, which in turn migrates to the nucleus regulating targeted gene expression183,184. STAT has seven subtypes: STAT1, 2, 3, 4, 5 A, 5B, and 6185,186. JAK signal-mediated transduction depends on the activation of PI3K/AKT/mTOR signaling187,188. Inhibition of PI3K/AKT/mTOR enhanced the effect of JAK2 inhibitors on primary human myeloproliferative neoplasm cells189. JAK/STAT could also be regulated by PDGFs. JAK2 and STAT3 was upregulated in left atrial and left ventricular fibroblasts treated with PDGF-AB190. Inhibition of JAK2 and STAT3 reversed PDGF-AB-induced collagen production in fibroblasts, suggesting that JAK2/STAT3 signaling was involved in PDGF-AB-induced fibrosis190. Furthermore, the activation of JAK/STAT signaling is required for TGF-β-mediated CTGF production in primary mouse HSCs191. JAK/STAT signals together with TGF-β1/Smad signals promote the EMT process in liver fibrosis192.
WNT/β-catenin
β-catenin is a transcription factor and its expression is mainly regulated by WNT proteins193,194. WNT/β-catenin activate and synergize with TGF-β1 to mediate the activation of myofibroblasts in lung fibrosis195,196. WNT/β-catenin signal was upregulated in TGF-β stimulated human lung fibroblasts197,198. Blocking β-catenin induced by TGF-β in vivo and in vitro can alleviate BLM-induced lung fibrosis199. In liver fibrosis, WNT/β-catenin also regulated the vimentin, collagen 1, and fibronectin in HSCs induced by TGF-β200. Apart from TGF-β, WNT/β-catenin can be regulated by CTGF via binding to the WNT coreceptor LDL receptor-related protein 6 (LRP6)201.
Apoptosis signal-regulating kinase 1 (ASK1) signaling pathway
ASK1 is involved in regulating glucose metabolism and maintaining energy homeostasis, which could activate the p38/cJun NH2-terminal kinase (JNK) signaling pathway202. Activation of the JNK signaling cascade suppressed the PPARα and FGF21 pathways203. Inhibition of ASK1 reduced insulin resistance, hepatic steatosis, inflammation, and fibrosis204,205.
Regulation of ECM cross-linking
Lysyl oxidases (LOXs) catalyses the conversion of lysine molecules to highly reactive aldehydes and enhances ECM (primarily collagen and elastin) cross-linking206,207. LOX family includes lysyl oxidase (LOX) and four lysyl oxidase-like proteins (LOXL1-4)208,209,210. The interaction of LOXs with TGF-β mediates the pro-fibrotic effect of LOXs in fibrosis. LOXL1 was required for TGF-β1 induced HSCs activation in liver fibrosis211. LOXL1 deficiency protected against TGF-β1-activated fibrosis and decreased the expression of fibrotic genes in vivo212. Silencing LOXL2 decreased mouse lung fibroblast proliferation and the levels of collagen 1α1 (COL1A1) via inhibition of TGF-β1/Smad2/3213.
Regulation of metabolism and inflammation
Alterations in metabolism can regulate the activation of inflammation-related pathways in epithelial cells, immune cells, and fibroblasts. The interactions between metabolism- and inflammation-related pathways modulate myofibroblasts activation. Signaling molecules that regulate metabolism may provide an interesting avenue for slowing the progression of fibrosis. As most of these signaling pathways regulating metabolism and inflammation are essential for NASH develpoment, the interactions between these signaling pathways in NASH are shown in Fig. 6.

Molecular signaling pathways of NASH and a summary of related target drugs. FFA, free fatty acid; TG, triglycerides
Peroxisome proliferator-activated receptors (PPARs) signaling pathway
PPARs are the nuclear receptors dependent on ligand binding214 and activate targeted genes related to lipid and glucose metabolism and adipogenesis215,216. There are three PPARs: PPARα, PPARγ, and PPARβ (also called δ)217,218. PPARα is most expressed in brown adipose tissue and liver219. The correlation of PPARs with liver fibrosis, especially NASH, is well-elaborated. PPARα is important for fatty acid metabolism220. Increased oxidative stress and hepatocyte apoptosis with higher NASH scores were observed in Pparα-null mice fed a high-fat diet221. Treatment with PPARα ligands attenuated liver fibrosis in rat thioacetamide models of liver cirrhosis222. Fasting-induced PPARα−/− mice showed low levels of FGF21, whereas FGF21 reduced hepatic triglycerides and cholesterol esters only in WT mice, suggesting that the effect of FGF21 on lipid metabolism might be partially dependent on PPARα223.
The function of PPARγ in NASH is more dependent on its role in inflammation. PPARγ activation inhibited inflammatory responses by inactivating nuclear factor-κB (NF-κB) signaling224 and reducing TNF-α and IL-1β expression in monocytes and macrophages225. Dual activation of PPARγ and PPARα has a favourable effect in ameliorating NASH by reducing inflammation, steatosis, and fibrosis226,227. PPAR-α and PPAR-γ activators have achieved efficacy in cardiac fibrosis228, renal fibrosis229 and pulmonary fibrosis230 animal models.
PPARβ/δ is mainly expressed in hepatocytes, Kupffer cells, and HSCs in liver231,232. PPARβ/δ-null mice exhibited aggravated hepatoxicity in carbon tetrachloride (CCl4)-treated mice233. However, the contraditory effects of PPARβ/δ agonists on HSCs proliferation and liver fibrosis hindered PPARβ/δ agonists from entering clinical trials234,235,236, which might be due to discrepancies in the ligands, dosage, and in vivo pharmacological properties of compounds.
Farnesoid X receptor (FXR) signaling pathway
FXR, as a nuclear receptor mainly located in enterohepatic tissues, can be activated by bile acids and regulate lipid and glucose metabolism237,238,239. FXR forms a heterodimer with the 9-cis-retinoic acid receptor and binds to farnesoid X response elements (FXREs), thus regulating target gene expression240. The roles of FXR vary in different organs. FXR expression was upregulated in lung fibrosis, and inhibition of FXR inhibited the bile acid-induced EMT and activation of lung fibroblasts241. However, FXR was reported to exert anti-fibrotic effect on kidney fibrosis and liver fibrosis. Treatment with FXR-activating ligand ameliorated triglyceride accumulation, improved proteinuria, and decreased ECM deposition in kidney disease experimental models242. FXR activation also protected hepatocytes from liver injury by inhibiting the activation of the NACHT, LRR, and PYD domain-containing protein 3 (NLRP3) inflammasome243. The interaction of FXR with other molecules is involved in bile acids circulation and plays an important role in NASH244. PPARα activation was required for the mRNA expression of FXR in the liver of fasted mice245. FXR directly regulated the expression of FGF19, thereby regulating hepatic protein and glycogen metabolism246,247. FXR/FGF19 axis increased FGF21 secretion248,249. FXR might also directly activate the expression of FGF21 by interacting with the FXRE in the 5’-flanking region of the FGF21 gene248.
Toll-like receptor 4 (TLR4) signaling pathway
TLR4, a member of the TLR family, functions as a crucial regulator in the immune system and inflammatory response. Fibroblast-specific deletion of TLR4 protected from mice lung and skin fibrosis250. In liver fibrosis, HSCs are the main effector cells of TLR4. TLR4 could sensitize HSCs to TGF-β stimulation and promote the activation of Kupffer cells, regulating hepatitis and liver fibrosis251. Activation of the TLR4/NF-κB signaling pathway induced hepatic inflammation252,253. However, TLR4 is an important receptor for AT2 proliferation and deletion of TLR4 in surfactant-protein-C-positive AT2 cells leads to impaired renewal capacity, severe fibrosis and mortality in IPF254.
GIP/GIPR and GLP-1/GLP-1R
Gastric inhibitory polypeptide (GIP) and glucagon-like peptide-1 (GLP-1) are the two major incretin hormones produced by the intestine that regulate insulin and glucagon secretion and food ingestion255. GIP is secreted by K cells in the upper part of the small intestine, while GLP-1 is mainly released by intestinal endocrine cells256,257,258. GIP exerts biological functions via binding to its receptor GIPR259,260 and was related to the activation of macrophages261,262. GLP-1 is expressed in various cells and binds to GLP-1R263. GLP-1 could downregulate collagen expression and TGF-β1 expression via regulating FGF21 in NASH mouse models264,265 and activating AMP-activated protein kinase (AMPK) in diabetic lung fibrosis266. Combined treatment with GLP-1R and GIPR agonists improved NASH steatosis, lobular inflammation, hepatocyte ballooning, and fibrosis267.
Anti-fibrotic drugs and clinical trials
Numerous small molecules or compounds are currently in clinical trials for fibrosis. Published clinical data on these compounds were listed in Table 1, and we categorized these drugs by targets and then ranked each target drug by clinical trial grade (marketed, phase 3, phase 2, and phase 1). Accordingly, antifibrotic drugs that have published clinical data and are in Phase 2, Phase 3 clinical trials or marketed are summarized in this part based on the ranking results.
Anti-fibrotic drugs targeting TGF-β
Most anti-TGF-β therapeutic drugs fall into five groups268,269: (1) nucleic acid drugs that blocking TGF-β synthesis. (2) TGF-β receptor kinases inhibitors, which block ATP binding to TGFβR, thus inhibiting Smad2 and Smad3 activation. (3) monoclonal antibodies preventing TGF-β from binding to its receptors. (4) high-affinity ligand traps prevent TGF-β from binding to its receptor. These inhibitors contain TβRII extracellular domains that could prevent TGF-β1 and TGF-β3 binding to TβRII receptors. (5) Some antibodies or molecules inhibiting the TGF-β activation, for example, drugs targeting αv/β integrins. Anti-fibrotic drugs targeting TGF-β now in clinical trials are mainly used in two diseases, IPF and myelofibrosis. Selected drugs targeting TGF-βs are described in detail.
Pirfenidone
Pirfenidone (PFD) is one of two FDA-approved drugs for IPF270, which inhibits both the synthesis and activition of TGF-βs271. The action mechanism of PFD in IPF has not been fully elaborated. Studies showed that PFD could inhibit the fibroblasts activation, reducing the synthesis of type 1 and type 3 collagen and the deposition of ECM272,273,274,275. Clinical trials demonstrated that PFD reduced lung function decline, decreased mortality, and improved overall survival of IPF patients276,277,278,279,280. Anorexia, rash, and gastrointestinal disorders are reported to be common side effects of PFD281. Based on the effect of PFD on improving inflammation and fibrosis in IPF, clinical studies on PFD for other types of pulmonary fibrosis are in progress. HEC-585 is a pyrimidine compound that is structurally related to PFD. Two phase I clinical trials were carried out to evaluate the safety, tolerability, and pharmacokinetics of HEC-585 in healthy subjects (NCT04512170 and NCT03092102).
Hydronidone
Hydronidone is a derivative of PFD with potential therapeutic efficacy for hepatic fibrosis282. The results of an open-label, randomized, dose-escalating study showed that hydronidone was well tolerated and effectively absorbed in healthy Chinese subjects (ChiCTR-ONC-12002899)282. Currently, a phase III study on the efficacy of hydronidone in HBV-induced liver fibrosis is in progress.
Luspatercept
Luspatercept is a recombinant fusion protein that binds TGF-β ligands to reduce Smad2/3 signaling. Luspatercept has been evaluated in myelofibrosis-associated anemia with 33 patients received concomitant ruxolitinib. Among transfusion-independent patients, 2 patients who did not receive ruxolitinib (10%) and 3 patients who received ruxolitinib (21%) experienced an increase of hemoglobin about 1.5 g/dL over 12 weeks. In the transfusion dependent cohort, 2 patients who did not receive ruxolitinib and 6 patients who received ruxolitinib were transfusion independent for at least 12 weeks283
AVID-200
AVID-200 contains soluble, dimerized, Fc-linked TβRII ectodomains and can be a high-affinity ligand trap preventing TGF-β from binding to its receptor. Treatment of myelofibrosis mononuclear cells with AVID-200 increased numbers of progenitor cells with wild type JAK2 but not mutated JAK2V617F284. Phase 1 clinical study in 12 myelofibrosis patients with ruxolitinib resistant showed that eight patients with grade 3/4 adverse reactions did not have dose-limiting toxicity and had improved platelet counts, with an average increase of 48%283.
Anti-fibrotic drugs targeting RTKs
Nintedanib
Nintedanib is a receptor tyrosine kinase inhibitor (RTKs: FGFRs, VEGFRs, and PDGFRs) that targets growth factor pathways, including FGFRs, VEGFRs, and PDGFRs285. In BLM-treated and silica-induced fibrosis mouse models, nintedanib reduced lung inflammation and fibrosis by decreasing total collagen, inflammatory chemokines, and pro-fibrotic factors both in therapeutic and preventive regimens157,286. Clinical trials have shown that nintedanib decreased the decline in FVC287 and reduced disease progression in IPF patients288,289,290,291. Nintedanib had acceptable safety and tolerability292,293, of which nausea and diarrhoea were the common side effects in the treatment of IPF294. The combination of PFD and nintedanib might produce synergistic effects and provide new prospects for the treatment of IPF277. However, both nintedanib and PFD have some problems such as high liver toxicity, high dosage, and photoallergic reaction, thus their long-term drug tolerance needs to be further determined.
ZSP1603
ZSP1603 (also known as WXFL-152), identified from a series of 4-hydroxyquinoline derivatives, targets VEGFR2, FGFRs, and PDGFRβ295. Our previous study showed the ability of ZSP1603 to reduce pulmonary injury, inflammation, and fibrosis in BLM-treated mice and rats296. ZSP1603 could inhibit the proliferation of primary human pulmonary fibroblasts (pHPFs) by blocking the PDGFRβ/ERK signaling pathway and decrease the differentiation of pHPFs by reducing TGF-β1, tissue inhibitor of metalloproteinase -1, and COL1A1296. The clinical study of ZSP1603 is expected to provide a new choice for IPF therapy.
Anti-fibrotic drugs targeting CTGF
Pamrevlumab
Pamrevlumab is a recombinant antibody that targets CTGF and inactivates its downstream inflammatory signals170. In a phase II, randomized, double-blind, placebo-controlled PRAISE trial involving 7 countries, pamrevlumab decreased the decline in FVC and inhibited the disease progression of IPF (NCT01890265)297. More therapeutic effects of pamrevlumab is expected to be investigated in phase III clinical trials (NCT04419558).
Anti-fibrotic drugs targeting PI3K
PI3K/AKT palys an important role in fibrotic processes and represents a critical target for the development of novel anti-fibrotic strategies. PI3K/AKT inhibitors are currently in clinical evaluation in IPF and myelofibrosis.
Parsaclisib
Parsaclisib is a potent PI3Kδ inhibitor and exerts antitumour effects in models of B-cell malignancy298. Single-dose parsaclisib alone or combination with itraconazole or rifampin achieved safety and toleratility in healthy subjects299. Two clinical trials in phase III studies (NCT04551066 and NCT04551053) were launched to test the efficacy and safety of parsaclisib and ruxolitinib in myelofibrosis.
Omipalisib
Omipalisib (GSK-2126458) is a dual inhibitor of PI3K/mTOR. Omipalisib inhibited the proliferation of pHPFs and decreased collagen accumulation induced by TGF-β1 in pHPFs176. Omipalisib was well absorbed and reached the lung in a randomized, placebo-controlled, double-blind phase I study in subjects with IPF (NCT01725139)300. Diarrhoea was the most commonly reported side effect of omipalisib300.
Anti-fibrotic drugs targeting JAKs
Since JAKs are essential for the occurrence and development of myelofibrosis, JAK inhibitors have achieved improvements in quality of life in patients with myelofibrosis. However, most drugs targeting JAK/STAT did not seem to prevent myelofibrosis patients from progressing to acute myeloid leukemia301.
Ruxolitinib
Ruxolitinib, a JAK1/JAK2 inhibitor, is approved by the FDA for patients with intermediate- and high-risk myelofibrosis. The effect of ruxolitinib in anemic myelofibrosis patients was evaluated in a phase 2 study (NCT02966353), who received ruxolitinib at 10 mg for the first 12 weeks, followed by escalating doses to 25 mg. During the study, palpable spleen length was reduced at least 50% in 70% patients receiving ruxolitinib, but 11.8% of patients needed platelet transfusion. The results also showed that the platelet counts and hemoglobin level of patients receiving increased dose were similar to those of patients who did not receice a dose increase302.
Momelotinib
Momelotinib (also known as CYT387, a JAK1/2 inhibitor) showed favorable therapeutic effects on myelofibrosis in preclinical trials by reducing multiple myeloma proliferation, inducing apoptosis of JAK2-dependent haematopoietic cells, and regulating inflammatory cytokines303. In a phase 3 study (NCT02101268), 156 patients with myeloid fibrosis were assigned to receive momelotinib (104) or standard care (52, 89% of whom received ruxolitinib). Encountered with the standard intervention group (6% of patients), 7% of patients in the momelotinib group had at least a 35% reduction in spleen volume. 11% of patients experienced peripheral neuropathy in the momelotinib group, compared with none in the standard intervention group304. Moreover, compared with ruxolitinib, the blood transfusion requirements and drug dependence of momelotinib were markedly reduced305.
Fedratinib
Fedratinib is a JAK2 inhibitor and has been used in treatment for patients with myeloproliferative neoplasm-associated myelofibrosis306. After 24 weeks, patients in the 400 mg fedratinib group had a 47% spleen volume response rate compared with 1% of patients with myelofibrosis in the placebo group. In this study, the two most common adverse reactions in patients taking fedratinib were anemia and diarrhea307.
Pacritinib
Pacritinib is an inhibitor of JAK2 and FMS-like tyrosine kinase 3. Pacritinib has good tolerance and clinical activity in myelofibrosis308,309. Twice daily pacritinib resulted in a significant reduction in spleen volume and improvements in the total symptom score over the best available therapy for myelofibrosis310.
Itacitinib
Itacitinib (INCB039110), a selective JAK1 inhibitor, has demonstrated favourable safety and anticancer effects311. Itacitinib exerts its anti-inflammatory effects by reducing pro-inflammatory cytokines and regulating the polarization of macrophages312. Administration of itacitinib at 200 mg twice daily and 600 mg once daily reduced the total symptom score in patients with myelofibrosis, and decreased the requirement of red blood cell units transfused in patients who needed transfusions during the 12 weeks prior to itacitinib treatment (NCT01633372)313.
Anti-fibrotic drugs targeting β-catenin
PRI-724
PRI-724 (also known as ICG-001) is a small molecule drug that modulate β-catenin/CBP transcription314,315. Preclinical studies demonstrated the efficacy of PRI-724 in decreasing ECM deposition and hepatic inflammation in a mouse model of CCl4-induced acute liver injury315 and a mouse model of HCV-infection316. In a dose escalation phase I trial, PRI-724 was well-tolerated in patients with HCV-induced cirrhosis at the dose of 10 or 40 mg/m(2) daily for 12 weeks317. However, PRI-724 did not effectively reduce liver fibrosis in patients with HCV- and HBV-induced cirrhosis, either by sequential scoring or by measuring proportional area of collagen for 12 weeks, but significantly improved liver stiffness (NCT03620474)318.
Anti-fibrotic drugs targeting ASK-1
Selonsertib
Selonsertib (GS-4997), a small molecule inhibitor of ASK1, showed efficacy in reducing collagen deposition, fibrosis stage, steatosis, and inflammation in a phase 2 study319. However, the phase III clinical trial (NCT03053050) of selonsertib was terminated in NASH patients with bridging fibrosis or compensated cirrhosis because its effect in alleviating fibrosis was not obvious320.
Anti-fibrotic drugs targeting PPARs
Since PPARs are involved in glucose and lipid metabolism, PPARs ligands are expected to be promising therapeutic agents for NAFLD/NASH. However, PPARα ligands (Clofibrate and Fenofibrate) showed no effect in inflammation and fibrosis in NASH231. PPARβ/δ agonist (GW501516) reduced inflammatory cells migration, insulin resistance and lipid levels, and increased ALT concentration in NASH experimental model321, but GW501516 has been terminated due to safety concerns. PPARγ agonists alleviated steatosis and inflammation yet with little effect fibrosis, and long time of administration is a major concern231. The effect of dual or pan agonists of PPARs in NASH are summarized below.
Elafibranor
The targets of elafibranor (GFT505) are PPARα and PPARδ322. Our previous results showed that GFT505 could inhibit steatosis, inflammation, and fibrosis in a NASH mouse model, and reduce the expression of lipid metabolism-, inflammation-, and fibrosis-related signaling molecules323. Treatment with 120 mg/d elafibranor for 1 year reduced NASH progression and liver fibrosis stage324. However, a phase III study of elafibranor in NASH patients was terminated because it did not achieve the predicted efficacy without safety issues (NCT02704403).
Saroglitazar
Saroglitazar is a novel dual PPARα/γ agonist that regulates glucose metabolism and improve insulin resistance. NAFLD/NASH patients were given placebo or 1 mg, 2 mg, or 4 mg saroglitazar. After the week 16, the ALT changes in the group taking 1 mg, 2 mg and 4 mg saroglitazine were -25.5%, 27.7%, and -45.8%, respectively, while the ALT changes in the group taking placebo were 3.4%. Administration of saroglitazar 4 mg decreased adiponectin, insulin resistance, and triglycerides, and the avarage body weight in patients taking 4 mg saroglitazar increased by 1.5 kg compared with 0.3 kg in placebo group325.
Lanifibranor
Lanifibranor (IVA337) is a PPAR α/γ/δ triple activator that can reduce immune cells infiltration and decreased steatosis in NASH experimental models326. In a phase 2b study, NASH patients without cirrhosis received placebo or 800 mg or 1200 mg lanifibranor daily for 24 weeks (NCT03008070). Results showed that most biomarkers of lipid, inflammation, and fibrosis were improved in both dose groups of lanifibranor. However, compared with patients receiving 800 mg lanifibranor, those receiving 1200-mg dose of lanifibranor had greater decrease in the SAF (the steatosis, activity, fibrosis) score327.
Pemafibrate
Pemafibrate targeting PPARα modulator regulates lipid and glucose metabolism. Preclinical studies have shown that pemafibrate could improve insulin resistance, inhibit hepatocyte ballooning degeneration, decrease the NAFLD score, and reduce myeloid cell recruitment328,329. Liver stiffness and ALT level were reduced in patients with high-risk NAFLD who received 0.2 mg pemafibrate twice daily for 72 weeks in a phase 2 trial (NCT03350165)330.
Anti-fibrotic drugs targeting FXR
FXR has emerged as a promising therapeutic target for NAFLD/NASH due to its diverse functions that modulate bile acid metabolism, inflammation, and immune responses. FXR agonists could be divided into steroidal and nonsterodial, and pruritus is the most common side effect of these targeted drugs.
Obeticholic acid
Obeticholic acid, a steroidal FXR agonist, has been shown to improve NASH symptoms. In a phase 3 trial (NCT02548351), NASH patients were given placebo, or 10 mg or 25 mg of obeticholic acid daily. Improvement in fibrosis was achieved in 23% of patients in the obeticholic acid 25 mg group compared with 18% of patients in the 10-mg obeticholic acid group and 12% of patients in the placebo group. However, there was no difference of NASH resolution endpoint between the three groups (P = 0.13)331. Patients taking obeticholic acid usually stop or reduce their dosage because of severe pruritus.
Cilofexor
Cilofexor (GS-9674) is a potent and selective FXR nonsteroidal agonist which activates FXR in the intestine and does not experience enterohepatic circulation. Twenty-four weeks of cilofexor improved serum bile acids metabolism and decreased hepatic steatosis in patients with NASH, but there was no significant change in fibrosis (NCT02854605)332.
EDP-305
EDP-305 is an effective FXR agonist showing little cross reaction with other nuclear receptors. EDP-305 inhibited HSCs activation in vitro and reduced MCD-induced steatohepatitis and liver fibrosis333. Liver fat and ALT level were reduced in NASH patients receiving 2.5 mg of EDP-305 compared with placebo group334. Pruritus was also one of the most common adverse events of EDP-305334.
Tropifexor
Tropifexor is a non-steroidal FXR agonist and significantly reduced steatohepatitis and fibrosis in NASH preclinical model335. Tropifexor was well tolerated up to 3000 µg and 100 µg in the single- and multiple-ascending doses (SAD/MAD) studies, respectively336, and is currently in phase 2 development for NASH.
Anti-fibrotic drug targeting TLR4
JKB-121
JKB-121 is a nonselective opioid TLR4 antagonist that has been proved to reduce LPS-induced liver inflammation in a MCD-induced model of NAFLD and inhibit the activation of HSCs337.
Anti-fibrotic drugs targeting GIP and GLP-1
FXR mainly negatively regulates liver gluconeogenesis, lipogenesis, and steatosis, while GIP and GLP-1 regulates glucose and lipid metabolism by reducing appetite, regulating liver fat content and inflammation. The dual receptor agonist of GIP and GLP-1 has been considered as an important therapeutic target for NASH.
Tirzepatide
Tirzepatide (LY3298176), a dual GIP and GLP-1 receptor agonist, has been used to explore its efficacy in clinical trials for the treatment of NASH, obesity, and type 2 diabetes mellitus (T2DM)338,339. Treatment with 10 mg of tirzepatide reduced NASH-related biomarkers, such as serum ALT and aspartate aminotransferase (AST), in patients with T2DM (NCT03131687)338. A phase III trial investigating tirzepatide in NASH patients is currently in progress (NCT04166773).
Semaglutide
Semaglutide is a GLP-1 receptor agonist and has been approved for T2DM therapy. In a 72-week phase 2 trial, NASH patients with liver fibrosis of stage F1, F2, or F3 received placebo, or semaglutide at 0.1 mg, 0.2 mg, or 0.4 mg. The percentage of patients who achieved NASH improvement without worsening fibrosis was 40%, 36% and 59% in the 0.1 mg semaglutide group, 0.2 mg semaglutide group and 0.4 mg semaglutide group, respectively, and 17% in the placebo group. However, the changes in fibrosis was not statistically significant in the 0.4 mg semaglutide group (43% of the patients) and in the placebo group (33% of the patients, P = 0.48)340.
Cotadutide
Cotadutide (MEDI0382) is a dual receptor agonist of GIP and GLP-1 and has shown safety and tolerability341. Cotadutide reduced hepatic lipid content, inflammation, steatosis, and NAS score in a mouse model of NASH342.
Anti-fibrotic drugs targeting CFTR
Drugs that improve the structure and function of CFTR have good therapeutic prospects in cystic fibrosis. At present, two kinds of drugs with different action mechanisms but complementary therapeutic effects have been developed, namely, CFTR potentiators and CFTR correctors343. CFTR potentiators enhance the gating of CFTR at the cell surface to mediate ion transport and are very effective in treating gated mutations344. CFTR correctors modify the processing and transportation of CFTR protein in cells, thus increasing the number of functional CFTR on the cell surface345.
Ivacaftor and Tezacaftor
Ivacaftor (VX-770) is the first CFTR potentiators approved by the FDA for cystic fibrosis patients with the gated mutation. Tezacaftor is a CFTR corrector approved by the FDA to be utilized in combination with ivacaftor. In a phase 2 clinical study, daily intake of 100 mg tezacaftor and 150 mg ivacaftor every 12 hours was effective in reducing chloride ion concentration in the sweat of cystic fibrosis patients, while increasing the percent predicted FEV1 (ppFEV1) value by 3.75% (NCT01531673)346.
Lumacaftor
Lumacaftor (VX-809), a CFTR corrector, is usually used in combination with ivacaftor for the treatment of cystic fibrosis. Lumacaftor increased the trafficking of CFTR protein to the extracellular membrane, while ivacaftor enabled the opening of dysfunctional chloride channels347. In 6- to 11-year-old patients with cystic fibrosis, sweat chloride concentration and CFQ-R RD score were improved after lumacaftor/ivacaftor combination therapy, but the FEV1 parameter was not changed (NCT02514473)348. However, FEV1 increased in patients with cystic fibrosis aged 12 years or older in a combination therapy with lumacaftor and ivacaft (NCT01807949)349.
ABBV-2222
ABBV-2222 (GLPG2222) is a novel and potent CFTR corrector350. Oral administration of ABBV-2222 once daily for 29 days in patients with homozygous or heterozygous of F508del CFTR and a gating mutation reduced sweat chloride concentrations in a dose-dependent manner without ppFEV1 improvements (NCT03119649 and NCT03045523)351.
Eluforsen
Eluforsen is an antisense oligonucleotide targeting the F508del mutation mRNA region to restore CFTR function352. Inhalation of eluforsen by single or multiple doses (up to 50 mg) demonstrated safety and tolerability353. In a phase 1b study, cystic fibrosis patients with a FEV1 > 70% in four single ascending dose cohorts and four MAD cohorts received eluforsen three times weekly for 4 weeks. CFQ-R Respiratory Symptom Score was improved in subjects of three groups in the MAD study353.
Conclusions
The high mortality and complex pathogenesis of fibrotic diseases pose great challenges in clinical therapy. Various cells and signaling pathways are involved in the progression of fibrosis. Drugs targeting these abnormal pathways are constantly being developed, and most of them demonstrate good anti-fibrotic properties in clinical trials. However, the side effects of these drugs often lead to drug discontinuation. Therefore, reducing adverse effects is also a great challenge for drug development. In addition, due to the complicated interaction of these signaling pathways in fibrosis, multitarget drug regimens would be beneficial for fibrosis therapy. In conclusion, this review provides reference for further mechanism and drug study of fibrosis.
References
Zhao, X. et al. Targeting metabolic dysregulation for fibrosis therapy. Nat. Rev. Drug Disco. 19, 57–75 (2020).
Global burden of 369 diseases and injuries in 204 countries and territories, 1990-2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet. 396, 1204–1222 (2020).
Shu, D. Y. & Lovicu, F. J. Myofibroblast transdifferentiation: The dark force in ocular wound healing and fibrosis. Prog. Retin Eye Res 60, 44–65 (2017).
Rockey, D. C., Bell, P. D. & Hill, J. A. Fibrosis-a common pathway to organ injury and failure. N. Engl. J. Med 372, 1138–1149 (2015).
Tomasek, J. J. et al. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 3, 349–363 (2002).
Desmoulière, A., Redard, M., Darby, I. & Gabbiani, G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am. J. Pathol. 146, 56–66 (1995).
Mascharak, S. et al. Multi-omic analysis reveals divergent molecular events in scarring and regenerative wound healing. Cell Stem Cell 29, 315–327.e316 (2022).
Hinz, B. & Lagares, D. Evasion of apoptosis by myofibroblasts: a hallmark of fibrotic diseases. Nat. Rev. Rheumatol. 16, 11–31 (2020).
Beyer, C. et al. Hypoxia. Hypoxia in the pathogenesis of systemic sclerosis. Arthritis Res Ther. 11, 220 (2009).
Tsou, P. S. et al. Inhibition of EZH2 prevents fibrosis and restores normal angiogenesis in scleroderma. Proc. Natl Acad. Sci. USA 116, 3695–3702 (2019).
Nanthakumar, C. B. et al. Dissecting fibrosis: therapeutic insights from the small-molecule toolbox. Nat. Rev. Drug Disco. 14, 693–720 (2015).
Parola, M. & Pinzani, M. Liver fibrosis: Pathophysiology, pathogenetic targets and clinical issues. Mol. Asp. Med 65, 37–55 (2019).
Dhar, D., Baglieri, J., Kisseleva, T. & Brenner, D. A. Mechanisms of liver fibrosis and its role in liver cancer. Exp. Biol. Med (Maywood) 245, 96–108 (2020).
Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 392, 1789–1858 (2018).
Asrani, S. K., Devarbhavi, H., Eaton, J. & Kamath, P. S. Burden of liver diseases in the world. J. Hepatol. 70, 151–171 (2019).
Murray, C. J. et al. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990-2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380, 2197–2223 (2012).
Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 392, 1736–1788 (2018).
Younossi, Z. et al. Global burden of NAFLD and NASH: trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 15, 11–20 (2018).
Byrne, C. D. & Targher, G. NAFLD: a multisystem disease. J. Hepatol. 62, S47–S64 (2015).
Yu, S. M. & Bonventre, J. V. Acute kidney injury and maladaptive tubular repair leading to renal fibrosis. Curr. Opin. Nephrol. Hypertens. 29, 310–318 (2020).
Pei, G., Yao, Y. & Yang, Q. Lymphangiogenesis in kidney and lymph node mediates renal inflammation and fibrosis. Sci. Adv. 5, eaaw5075 (2019).
Majo, J., Klinkhammer, B. M., Boor, P. & Tiniakos, D. Pathology and natural history of organ fibrosis. Curr. Opin. Pharm. 49, 82–89 (2019).
Global, regional, and national burden of chronic kidney disease, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 395, 709–733 (2020).
Krenning, G., Zeisberg, E. M. & Kalluri, R. The origin of fibroblasts and mechanism of cardiac fibrosis. J. Cell Physiol. 225, 631–637 (2010).
Mewton, N. et al. Assessment of myocardial fibrosis with cardiovascular magnetic resonance. J. Am. Coll. Cardiol. 57, 891–903 (2011).
Nattel, S. Molecular and Cellular Mechanisms of Atrial Fibrosis in Atrial Fibrillation. JACC Clin. Electrophysiol. 3, 425–435 (2017).
Timmis, A. et al. European Society of Cardiology: Cardiovascular Disease Statistics 2017. Eur. Heart J. 39, 508–579 (2018).
Zhou, M. et al. Cause-specific mortality for 240 causes in China during 1990-2013: a systematic subnational analysis for the Global Burden of Disease Study 2013. Lancet 387, 251–272 (2016).
Wilson, M. S. & Wynn, T. A. Pulmonary fibrosis: pathogenesis, etiology and regulation. Mucosal Immunol. 2, 103–121 (2009).
Lederer, D. J. & Martinez, F. J. Idiopathic Pulmonary Fibrosis. N. Engl. J. Med 378, 1811–1823 (2018).
Martinez, F. J. et al. Idiopathic pulmonary fibrosis. Nat. Rev. Dis. Prim. 3, 17074 (2017).
O’Dwyer, D. N., Ashley, S. L. & Moore, B. B. Influences of innate immunity, autophagy, and fibroblast activation in the pathogenesis of lung fibrosis. Am. J. Physiol. Lung Cell Mol. Physiol. 311, L590–L601 (2016).
Maher, T. M. et al. Global incidence and prevalence of idiopathic pulmonary fibrosis. Respir. Res 22, 197 (2021).
Borchers, A. T., Chang, C., Keen, C. L. & Gershwin, M. E. Idiopathic pulmonary fibrosis-an epidemiological and pathological review. Clin. Rev. Allergy Immunol. 40, 117–134 (2011).
Esposito, D. B. et al. Idiopathic Pulmonary Fibrosis in United States Automated Claims. Incidence, Prevalence, and Algorithm Validation. Am. J. Respir. Crit. Care Med 192, 1200–1207 (2015).
Mossman, B. T. & Churg, A. Mechanisms in the pathogenesis of asbestosis and silicosis. Am. J. Respir. Crit. Care Med 157, 1666–1680 (1998).
Mutsaers, S. E. et al. Fibroblast mitogens in bronchoalveolar lavage (BAL) fluid from asbestos-exposed subjects with and without clinical evidence of asbestosis: no evidence for the role of PDGF, TNF-alpha, IGF-1, or IL-1 beta. J. Pathol. 185, 199–203 (1998).
Cullinan, P. & Reid, P. Pneumoconiosis. Prim. Care Respir. J. 22, 249–252 (2013).
Naehrig, S., Chao, C. M. & Naehrlich, L. Cystic Fibrosis. Dtsch Arztebl Int 114, 564–574 (2017).
Powers, C. A., Potter, E. M., Wessel, H. U. & Lloyd-Still, J. D. Cystic fibrosis in Asian Indians. Arch. Pediatr. Adolesc. Med 150, 554–555 (1996).
Yamashiro, Y. et al. The estimated incidence of cystic fibrosis in Japan. J. Pediatr. Gastroenterol. Nutr. 24, 544–547 (1997).
Guo, X. et al. Clinical and genetic characteristics of cystic fibrosis in CHINESE patients: a systemic review of reported cases. Orphanet J. Rare Dis. 13, 224 (2018).
Schieber, M., Crispino, J. D. & Stein, B. Myelofibrosis in 2019: moving beyond JAK2 inhibition. Blood Cancer J. 9, 74 (2019).
Tefferi, A. Primary myelofibrosis: 2014 update on diagnosis, risk-stratification, and management. Am. J. Hematol. 89, 915–925 (2014).
Hultcrantz, M. et al. Patterns of survival among patients with myeloproliferative neoplasms diagnosed in Sweden from 1973 to 2008: a population-based study. J. Clin. Oncol. 30, 2995–3001 (2012).
Zahr, A. A. et al. Bone marrow fibrosis in myelofibrosis: pathogenesis, prognosis and targeted strategies. Haematologica 101, 660–671 (2016).
Adams, T. S. et al. Single-cell RNA-seq reveals ectopic and aberrant lung-resident cell populations in idiopathic pulmonary fibrosis. Sci. Adv. 6, eaba1983 (2020).
Ramachandran, P. et al. Resolving the fibrotic niche of human liver cirrhosis at single-cell level. Nature 575, 512–518 (2019).
Xue, D. et al. Expansion of Fcγ Receptor IIIa-Positive Macrophages, Ficolin 1-Positive Monocyte-Derived Dendritic Cells, and Plasmacytoid Dendritic Cells Associated With Severe Skin Disease in Systemic Sclerosis. Arthritis Rheumatol. 74, 329–341 (2022).
Yang, W. et al. Single-Cell Transcriptomic Analysis Reveals a Hepatic Stellate Cell-Activation Roadmap and Myofibroblast Origin During Liver Fibrosis in Mice. Hepatology 74, 2774–2790 (2021).
Carraro, G. et al. Transcriptional analysis of cystic fibrosis airways at single-cell resolution reveals altered epithelial cell states and composition. Nat. Med 27, 806–814 (2021).
Parimon, T. et al. Alveolar Epithelial Type II Cells as Drivers of Lung Fibrosis in Idiopathic Pulmonary Fibrosis. Int. J. Mol. Sci. 21, 2269 (2020).
Yao, L. et al. Paracrine signalling during ZEB1-mediated epithelial-mesenchymal transition augments local myofibroblast differentiation in lung fibrosis. Cell Death Differ. 26, 943–957 (2019).
Reyfman, P. A. et al. Single-Cell Transcriptomic Analysis of Human Lung Provides Insights into the Pathobiology of Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med 199, 1517–1536 (2019).
Xu, Y. et al. Single-cell RNA sequencing identifies diverse roles of epithelial cells in idiopathic pulmonary fibrosis. JCI Insight 1, e90558 (2016).
Barkauskas, C. E. et al. Type 2 alveolar cells are stem cells in adult lung. J. Clin. Invest 123, 3025–3036 (2013).
Zacharias, W. J. et al. Regeneration of the lung alveolus by an evolutionarily conserved epithelial progenitor. Nature 555, 251–255 (2018).
Zepp, J. A. et al. Distinct Mesenchymal Lineages and Niches Promote Epithelial Self-Renewal and Myofibrogenesis in the Lung. Cell 170, 1134–1148.e1110 (2017).
Kathiriya, J. J. et al. Human alveolar type 2 epithelium transdifferentiates into metaplastic KRT5(+) basal cells. Nat. Cell Biol. 24, 10–23 (2022).
Montoro, D. T. et al. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 560, 319–324 (2018).
Stutts, M. J. et al. CFTR as a cAMP-dependent regulator of sodium channels. Science 269, 847–850 (1995).
Ratjen, F. & Döring, G. Cystic fibrosis. Lancet 361, 681–689 (2003).
Olivier, A. K., Gibson-Corley, K. N. & Meyerholz, D. K. Animal models of gastrointestinal and liver diseases. Animal models of cystic fibrosis: gastrointestinal, pancreatic, and hepatobiliary disease and pathophysiology. Am. J. Physiol. Gastrointest. Liver Physiol. 308, G459–G471 (2015).
Ledder, O. et al. Cystic fibrosis: an update for clinicians. Part 2: hepatobiliary and pancreatic manifestations. J. Gastroenterol. Hepatol. 29, 1954–1962 (2014).
Feranchak, A. P. & Sokol, R. J. Cholangiocyte biology and cystic fibrosis liver disease. Semin Liver Dis. 21, 471–488 (2001).
Frantz, C. et al. IL-2-related regulatory CD4 T-cell deficiency leads to the development of lung fibrosis and vascular remodeling. Arthritis Rheumatol (2022).
Valenzi, E. et al. Disparate Interferon Signaling and Shared Aberrant Basaloid Cells in Single-Cell Profiling of Idiopathic Pulmonary Fibrosis and Systemic Sclerosis-Associated Interstitial Lung Disease. Front Immunol. 12, 595811 (2021).
Morimoto, Y. et al. Amphiregulin-Producing Pathogenic Memory T Helper 2 Cells Instruct Eosinophils to Secrete Osteopontin and Facilitate Airway Fibrosis. Immunity 49, 134–150.e136 (2018).
Takemura, N. et al. Eosinophil depletion suppresses radiation-induced small intestinal fibrosis. Sci Transl Med. 10, eaan0333 (2018).
Meadows, V. et al. Mast Cells Regulate Ductular Reaction and Intestinal Inflammation in Cholestasis Through Farnesoid X Receptor Signaling. Hepatology 74, 2684–2698 (2021).
Serezani, A. P. et al. Multi-Platform Single-Cell Analysis Identifies Immune Cell Types Enhanced in Pulmonary Fibrosis. Am. J. Respir. Cell. Mol. Biol. 10.1165/rcmb.2021-0418OC (2022).
Liang, Q. et al. The T Cell Receptor Immune Repertoire Protects the Liver From Reconsitution. Front Immunol. 11, 584979 (2020).
Byrne, A. J., Mathie, S. A., Gregory, L. G. & Lloyd, C. M. Pulmonary macrophages: key players in the innate defence of the airways. Thorax 70, 1189–1196 (2015).
Wang, X. et al. MCPIP1 Regulates Alveolar Macrophage Apoptosis and Pulmonary Fibroblast Activation After in vitro Exposure to Silica. Toxicol. Sci. 151, 126–138 (2016).
Prasse, A. et al. A vicious circle of alveolar macrophages and fibroblasts perpetuates pulmonary fibrosis via CCL18. Am. J. Respir. Crit. Care Med 173, 781–792 (2006).
Zhang, W. et al. S100a4 Is Secreted by Alternatively Activated Alveolar Macrophages and Promotes Activation of Lung Fibroblasts in Pulmonary Fibrosis. Front Immunol. 9, 1216 (2018).
Aran, D. et al. Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage. Nat. Immunol. 20, 163–172 (2019).
Farbehi, N. et al. Single-cell expression profiling reveals dynamic flux of cardiac stromal, vascular and immune cells in health and injury. Elife. 8, e43882 (2019).
Tabib, T. et al. SFRP2/DPP4 and FMO1/LSP1 Define Major Fibroblast Populations in Human Skin. J. Invest Dermatol 138, 802–810 (2018).
Xie, T. et al. Single-Cell Deconvolution of Fibroblast Heterogeneity in Mouse Pulmonary Fibrosis. Cell Rep. 22, 3625–3640 (2018).
Yuan, Q., Tan, R. J. & Liu, Y. Myofibroblast in Kidney Fibrosis: Origin, Activation, and Regulation. Adv. Exp. Med Biol. 1165, 253–283 (2019).
Zeisberg, E. M. et al. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat. Med 13, 952–961 (2007).
Kato, K. et al. Impaired Myofibroblast Dedifferentiation Contributes to Nonresolving Fibrosis in Aging. Am. J. Respir. Cell Mol. Biol. 62, 633–644 (2020).
Xie, G. et al. Role of differentiation of liver sinusoidal endothelial cells in progression and regression of hepatic fibrosis in rats. Gastroenterology 142, 918–927.e916 (2012).
Deleve, L. D., Wang, X. & Guo, Y. Sinusoidal endothelial cells prevent rat stellate cell activation and promote reversion to quiescence. Hepatology 48, 920–930 (2008).
Seki, E. & Schwabe, R. F. Hepatic inflammation and fibrosis: functional links and key pathways. Hepatology 61, 1066–1079 (2015).
Tsuchida, T. & Friedman, S. L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 14, 397–411 (2017).
Mendez-Sanchez, N. et al. The cellular pathways of liver fibrosis in non-alcoholic steatohepatitis. Ann. Transl. Med 8, 400 (2020).
Radaeva, S. et al. Natural killer cells ameliorate liver fibrosis by killing activated stellate cells in NKG2D-dependent and tumor necrosis factor-related apoptosis-inducing ligand-dependent manners. Gastroenterology 130, 435–452 (2006).
Jeong, W. I. et al. Suppression of innate immunity (natural killer cell/interferon-γ) in the advanced stages of liver fibrosis in mice. Hepatology 53, 1342–1351 (2011).
Rosselli, M. et al. The metabolic syndrome and chronic liver disease. Curr. Pharm. Des. 20, 5010–5024 (2014).
Ramachandran, P., Iredale, J. P. & Fallowfield, J. A. Resolution of liver fibrosis: basic mechanisms and clinical relevance. Semin Liver Dis. 35, 119–131 (2015).
Zhang, Y. et al. Enhanced IL-1 beta and tumor necrosis factor-alpha release and messenger RNA expression in macrophages from idiopathic pulmonary fibrosis or after asbestos exposure. J. Immunol. 150, 4188–4196 (1993).
Chen, G. et al. IL-1β dominates the promucin secretory cytokine profile in cystic fibrosis. J. Clin. Invest 129, 4433–4450 (2019).
Frangogiannis, N. G. The inflammatory response in myocardial injury, repair, and remodelling. Nat. Rev. Cardiol. 11, 255–265 (2014).
Fan, D., Takawale, A., Lee, J. & Kassiri, Z. Cardiac fibroblasts, fibrosis and extracellular matrix remodeling in heart disease. Fibrogenes. Tissue Repair 5, 15 (2012).
Kolb, M. et al. Transient expression of IL-1beta induces acute lung injury and chronic repair leading to pulmonary fibrosis. J. Clin. Invest 107, 1529–1536 (2001).
Barron, L., Gharib, S. A. & Duffield, J. S. Lung Pericytes and Resident Fibroblasts: Busy Multitaskers. Am. J. Pathol. 186, 2519–2531 (2016).
Distler, J. H. W. et al. Shared and distinct mechanisms of fibrosis. Nat. Rev. Rheumatol. 15, 705–730 (2019).
Tzavlaki, K. & Moustakas, A. TGF-beta Signaling. Biomolecules. 10, 487 (2020).
Robertson, I. B. & Rifkin, D. B. Regulation of the Bioavailability of TGF-β and TGF-β-Related Proteins. Cold Spring Harb Perspect Biol. 8, a021907 (2016).
Meng, X. M., Nikolic-Paterson, D. J. & Lan, H. Y. TGF-beta: the master regulator of fibrosis. Nat. Rev. Nephrol. 12, 325–338 (2016).
Jakowlew, S. B. et al. Differential regulation of the expression of transforming growth factor-beta mRNAs by growth factors and retinoic acid in chicken embryo chondrocytes, myocytes, and fibroblasts. J. Cell Physiol. 150, 377–385 (1992).
Villiger, P. M. & Lotz, M. Differential expression of TGF beta isoforms by human articular chondrocytes in response to growth factors. J. Cell Physiol. 151, 318–325 (1992).
Shi, M. et al. Latent TGF-beta structure and activation. Nature 474, 343–349 (2011).
Aluwihare, P. et al. Mice that lack activity of alphavbeta6- and alphavbeta8-integrins reproduce the abnormalities of Tgfb1- and Tgfb3-null mice. J. Cell Sci. 122, 227–232 (2009).
Horan, G. S. et al. Partial inhibition of integrin alpha(v)beta6 prevents pulmonary fibrosis without exacerbating inflammation. Am. J. Respir. Crit. Care Med 177, 56–65 (2008).
Koivisto, L., Bi, J., Häkkinen, L. & Larjava, H. Integrin αvβ6: Structure, function and role in health and disease. Int J. Biochem Cell Biol. 99, 186–196 (2018).
Yan, X. & Chen, Y. G. Smad7: not only a regulator, but also a cross-talk mediator of TGF-beta signalling. Biochem J. 434, 1–10 (2011).
Finnson, K. W., Almadani, Y. & Philip, A. Non-canonical (non-SMAD2/3) TGF-β signaling in fibrosis: Mechanisms and targets. Semin Cell Dev. Biol. 101, 115–122 (2020).
Coker, R. K. et al. Localisation of transforming growth factor beta1 and beta3 mRNA transcripts in normal and fibrotic human lung. Thorax 56, 549–556 (2001).
Cai, X. et al. CXCL6-EGFR-induced Kupffer cells secrete TGF-β1 promoting hepatic stellate cell activation via the SMAD2/BRD4/C-MYC/EZH2 pathway in liver fibrosis. J. Cell Mol. Med 22, 5050–5061 (2018).
Thannickal, V. J. et al. Myofibroblast differentiation by transforming growth factor-beta1 is dependent on cell adhesion and integrin signaling via focal adhesion kinase. J. Biol. Chem. 278, 12384–12389 (2003).
Chanda, D. et al. Developmental pathways in the pathogenesis of lung fibrosis. Mol. Asp. Med 65, 56–69 (2019).
Cheng, Q. et al. Methyl ferulic acid attenuates liver fibrosis and hepatic stellate cell activation through the TGF-β1/Smad and NOX4/ROS pathways. Chem. Biol. Interact. 299, 131–139 (2019).
Magnani, F. & Mattevi, A. Structure and mechanisms of ROS generation by NADPH oxidases. Curr. Opin. Struct. Biol. 59, 91–97 (2019).
Jiménez-Uribe, A. P. et al. Backstage players of fibrosis: NOX4, mTOR, HDAC, and S1P; companions of TGF-β. Cell Signal 87, 110123 (2021).
Cucoranu, I. et al. NAD(P)H oxidase 4 mediates transforming growth factor-beta1-induced differentiation of cardiac fibroblasts into myofibroblasts. Circ. Res 97, 900–907 (2005).
Si, M. et al. Inhibition of hyperglycolysis in mesothelial cells prevents peritoneal fibrosis. Sci. Transl. Med. 11, eaav5341 (2019).
Yan, F. et al. Nox4 and redox signaling mediate TGF-β-induced endothelial cell apoptosis and phenotypic switch. Cell Death Dis. 5, e1010 (2014).
Negulescu, O. et al. Estradiol reverses TGF-beta1-induced mesangial cell apoptosis by a casein kinase 2-dependent mechanism. Kidney Int 62, 1989–1998 (2002).
Marconi, G. D. et al. Epithelial-Mesenchymal Transition (EMT): The Type-2 EMT in Wound Healing, Tissue Regeneration and Organ Fibrosis. Cells. 10, 1587 (2021).
Kim, S. J. et al. Activation of ERK1/2-mTORC1-NOX4 mediates TGF-β1-induced epithelial-mesenchymal transition and fibrosis in retinal pigment epithelial cells. Biochem Biophys. Res Commun. 529, 747–752 (2020).
Zhou, T. et al. Runt-Related Transcription Factor 1 (RUNX1) Promotes TGF-β-Induced Renal Tubular Epithelial-to-Mesenchymal Transition (EMT) and Renal Fibrosis through the PI3K Subunit p110δ. EBioMedicine 31, 217–225 (2018).
Papadopoulos, N. & Lennartsson, J. The PDGF/PDGFR pathway as a drug target. Mol. Asp. Med 62, 75–88 (2018).
Iwayama, T. & Olson, L. E. Involvement of PDGF in fibrosis and scleroderma: recent insights from animal models and potential therapeutic opportunities. Curr. Rheumatol. Rep. 15, 304 (2013).
Kazlauskas, A. PDGFs and their receptors. Gene 614, 1–7 (2017).
Borkham-Kamphorst, E. & Weiskirchen, R. The PDGF system and its antagonists in liver fibrosis. Cytokine Growth Factor Rev. 28, 53–61 (2016).
Martinet, Y. et al. Exaggerated spontaneous release of platelet-derived growth factor by alveolar macrophages from patients with idiopathic pulmonary fibrosis. N. Engl. J. Med 317, 202–209 (1987).
Vanhee, D. et al. Mechanisms of fibrosis in coal workers’ pneumoconiosis. Increased production of platelet-derived growth factor, insulin-like growth factor type I, and transforming growth factor beta and relationship to disease severity. Am. J. Respir. Crit. Care Med 150, 1049–1055 (1994).
Ying, H. Z. et al. PDGF signaling pathway in hepatic fibrosis pathogenesis and therapeutics (Review). Mol. Med Rep. 16, 7879–7889 (2017).
Heldin, P. et al. Differential expression of platelet-derived growth factor alpha- and beta- receptors on fat-storing cells and endothelial cells of rat liver. Exp. Cell Res 193, 364–369 (1991).
Borkham-Kamphorst, E. et al. PDGF-D signaling in portal myofibroblasts and hepatic stellate cells proves identical to PDGF-B via both PDGF receptor type alpha and beta. Cell Signal 27, 1305–1314 (2015).
Wang, X. et al. Targeting the PDGF-B/PDGFR-beta Interface with Destruxin A5 to Selectively Block PDGF-BB/PDGFR-betabeta Signaling and Attenuate Liver Fibrosis. EBioMedicine 7, 146–156 (2016).
Jiang, Y., Zhao, Y., He, F. & Wang, H. Artificial MicroRNA-Mediated Tgfbr2 and Pdgfrb Co-Silencing Ameliorates Carbon Tetrachloride-Induced Hepatic Fibrosis in Mice. Hum. Gene Ther. 30, 179–196 (2019).
Martin, I. V. et al. Platelet-derived growth factor (PDGF)-C neutralization reveals differential roles of PDGF receptors in liver and kidney fibrosis. Am. J. Pathol. 182, 107–117 (2013).
Eitner, F. et al. PDGF-C is a proinflammatory cytokine that mediates renal interstitial fibrosis. J. Am. Soc. Nephrol. 19, 281–289 (2008).
Nishioka, Y., Azuma, M., Kishi, M. & Aono, Y. Targeting platelet-derived growth factor as a therapeutic approach in pulmonary fibrosis. J. Med Invest 60, 175–183 (2013).
Tuuminen, R. et al. PDGF-A, -C, and -D but not PDGF-B increase TGF-beta1 and chronic rejection in rat cardiac allografts. Arterioscler Thromb. Vasc. Biol. 29, 691–698 (2009).
Gallini, R. et al. PDGF-A and PDGF-B induces cardiac fibrosis in transgenic mice. Exp. Cell Res 349, 282–290 (2016).
Mohammadi, M., Olsen, S. K. & Ibrahimi, O. A. Structural basis for fibroblast growth factor receptor activation. Cytokine Growth Factor Rev. 16, 107–137 (2005).
Ghedini, G. C., Ronca, R., Presta, M. & Giacomini, A. Future applications of FGF/FGFR inhibitors in cancer. Expert Rev. Anticancer Ther. 18, 861–872 (2018).
Bae, J. H. & Schlessinger, J. Asymmetric tyrosine kinase arrangements in activation or autophosphorylation of receptor tyrosine kinases. Mol. Cells 29, 443–448 (2010).
Katoh, M. Therapeutics Targeting FGF Signaling Network in Human Diseases. Trends Pharm. Sci. 37, 1081–1096 (2016).
Ornitz, D. M. & Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 4, 215–266 (2015).
Uriarte, I. et al. Ileal FGF15 contributes to fibrosis-associated hepatocellular carcinoma development. Int J. Cancer 136, 2469–2475 (2015).
Schumacher, J. D. et al. Direct and Indirect Effects of Fibroblast Growth Factor (FGF) 15 and FGF19 on Liver Fibrosis Development. Hepatology 71, 670–685 (2020).
Geng, L., Lam, K. S. L. & Xu, A. The therapeutic potential of FGF21 in metabolic diseases: from bench to clinic. Nat. Rev. Endocrinol. 16, 654–667 (2020).
Lewis, J. E., Ebling, F. J. P., Samms, R. J. & Tsintzas, K. Going Back to the Biology of FGF21: New Insights. Trends Endocrinol. Metab. 30, 491–504 (2019).
Sonoda, J., Chen, M. Z. & Baruch, A. FGF21-receptor agonists: an emerging therapeutic class for obesity-related diseases. Horm Mol Biol Clin Investig. 30 (2017).
Fisher, F. M. et al. Fibroblast growth factor 21 limits lipotoxicity by promoting hepatic fatty acid activation in mice on methionine and choline-deficient diets. Gastroenterology 147, 1073–1083.e1076 (2014).
Yin, J. et al. Enhanced expression and distinctive characterization of a long-acting FGF21 and its potential to alleviate nonalcoholic steatohepatitis. Biochimie 151, 166–175 (2018).
Joannes, A. et al. FGF9 and FGF18 in idiopathic pulmonary fibrosis promote survival and migration and inhibit myofibroblast differentiation of human lung fibroblasts in vitro. Am. J. Physiol. Lung Cell Mol. Physiol. 310, L615–L629 (2016).
Shimbori, C. et al. Fibroblast growth factor-1 attenuates TGF-β1-induced lung fibrosis. J. Pathol. 240, 197–210 (2016).
Koo, H. Y. et al. Fibroblast growth factor 2 decreases bleomycin-induced pulmonary fibrosis and inhibits fibroblast collagen production and myofibroblast differentiation. J. Pathol. 246, 54–66 (2018).
Yang, L. et al. FGF/FGFR signaling: From lung development to respiratory diseases. Cytokine Growth Factor Rev. 62, 94–104 (2021).
Wollin, L. et al. Mode of action of nintedanib in the treatment of idiopathic pulmonary fibrosis. Eur. Respir. J. 45, 1434–1445 (2015).
Hu, K. & Olsen, B. R. Vascular endothelial growth factor control mechanisms in skeletal growth and repair. Dev. Dyn. 246, 227–234 (2017).
Shibuya, M. VEGF-VEGFR Signals in Health and Disease. Biomol. Ther. (Seoul.) 22, 1–9 (2014).
Apte, R. S., Chen, D. S. & Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 176, 1248–1264 (2019).
Murray, L. A. et al. Antifibrotic role of vascular endothelial growth factor in pulmonary fibrosis. JCI Insight. 2, e92192 (2017).
Chaudhary, N. I. et al. Inhibition of PDGF, VEGF and FGF signalling attenuates fibrosis. Eur. Respir. J. 29, 976–985 (2007).
Farkas, L., Gauldie, J., Voelkel, N. F. & Kolb, M. Pulmonary hypertension and idiopathic pulmonary fibrosis: a tale of angiogenesis, apoptosis, and growth factors. Am. J. Respir. Cell Mol. Biol. 45, 1–15 (2011).
Ferrara, N. & Adamis, A. P. Ten years of anti-vascular endothelial growth factor therapy. Nat. Rev. Drug Disco. 15, 385–403 (2016).
Barratt, S. L., Flower, V. A., Pauling, J. D. & Millar, A. B. VEGF (Vascular Endothelial Growth Factor) and Fibrotic Lung Disease. Int. J. Mol. Sci. 19, 1269 (2018).
Barratt, S. L. et al. Effects of hypoxia and hyperoxia on the differential expression of VEGF-A isoforms and receptors in Idiopathic Pulmonary Fibrosis (IPF). Respir. Res 19, 9 (2018).
Lau, L. F. Cell surface receptors for CCN proteins. J. Cell Commun. Signal 10, 121–127 (2016).
Aoyama, E., Kubota, S. & Takigawa, M. CCN2/CTGF binds to fibroblast growth factor receptor 2 and modulates its signaling. FEBS Lett. 586, 4270–4275 (2012).
Toda, N., Mukoyama, M., Yanagita, M. & Yokoi, H. CTGF in kidney fibrosis and glomerulonephritis. Inflamm. Regen. 38, 14 (2018).
Lipson, K. E., Wong, C., Teng, Y. & Spong, S. CTGF is a central mediator of tissue remodeling and fibrosis and its inhibition can reverse the process of fibrosis. Fibrogenes. Tissue Repair 5, S24 (2012).
Mao, L. et al. MKL1 mediates TGF-β-induced CTGF transcription to promote renal fibrosis. J. Cell Physiol. 235, 4790–4803 (2020).
Wahab, N. A., Weston, B. S. & Mason, R. M. Modulation of the TGFbeta/Smad signaling pathway in mesangial cells by CTGF/CCN2. Exp. Cell Res 307, 305–314 (2005).
Fruman, D. A. et al. The PI3K Pathway in Human Disease. Cell 170, 605–635 (2017).
Janku, F., Yap, T. A. & Meric-Bernstam, F. Targeting the PI3K pathway in cancer: are we making headway? Nat. Rev. Clin. Oncol. 15, 273–291 (2018).
Lawrence, J. & Nho, R. The Role of the Mammalian Target of Rapamycin (mTOR) in Pulmonary Fibrosis. Int J Mol Sci. 19, E778 (2018).
Mercer, P. F. et al. Exploration of a potent PI3 kinase/mTOR inhibitor as a novel anti-fibrotic agent in IPF. Thorax 71, 701–711 (2016).
Wu, W. et al. Yu Jin Pulvis inhibits carbon tetrachloride-induced liver fibrosis by blocking the MAPK and PI3K/Akt signaling pathways. Am. J. Transl. Res 11, 5998–6006 (2019).
Conte, E. et al. Inhibition of PI3K prevents the proliferation and differentiation of human lung fibroblasts into myofibroblasts: the role of class I P110 isoforms. PLoS One 6, e24663 (2011).
Chetty, C., Lakka, S. S., Bhoopathi, P. & Rao, J. S. MMP-2 alters VEGF expression via alphaVbeta3 integrin-mediated PI3K/AKT signaling in A549 lung cancer cells. Int J. Cancer 127, 1081–1095 (2010).
Wang, L. et al. Neuropilin-1 aggravates liver cirrhosis by promoting angiogenesis via VEGFR2-dependent PI3K/Akt pathway in hepatic sinusoidal endothelial cells. EBioMedicine 43, 525–536 (2019).
Yan, Z. et al. CD147 promotes liver fibrosis progression via VEGF-A/VEGFR2 signalling-mediated cross-talk between hepatocytes and sinusoidal endothelial cells. Clin. Sci. (Lond.) 129, 699–710 (2015).
Gadina, M. et al. Janus kinases to jakinibs: from basic insights to clinical practice. Rheumatol. (Oxf.) 58, i4–i16 (2019).
Maruotti, N. & Corrado, A. Janus kinase inhibitors role in bone remodeling. J. Cell Physiol. 235, 1915–1920 (2020).
Leonard, W. J. & O’Shea, J. J. Jaks and STATs: biological implications. Annu Rev. Immunol. 16, 293–322 (1998).
Villarino, A. V., Kanno, Y. & O’Shea, J. J. Mechanisms and consequences of Jak-STAT signaling in the immune system. Nat. Immunol. 18, 374–384 (2017).
Wang, Y. & Levy, D. E. Comparative evolutionary genomics of the STAT family of transcription factors. Jakstat 1, 23–33 (2012).
Bartalucci, N., Guglielmelli, P. & Vannucchi, A. M. Rationale for targeting the PI3K/Akt/mTOR pathway in myeloproliferative neoplasms. Clin. Lymphoma Myeloma Leuk. 13, S307–S309 (2013).
Meyer, S. C. & Levine, R. L. Molecular pathways: molecular basis for sensitivity and resistance to JAK kinase inhibitors. Clin. Cancer Res 20, 2051–2059 (2014).
Fiskus, W. et al. Dual PI3K/AKT/mTOR inhibitor BEZ235 synergistically enhances the activity of JAK2 inhibitor against cultured and primary human myeloproliferative neoplasm cells. Mol. Cancer Ther. 12, 577–588 (2013).
Chen, Y. et al. JAK-STAT signalling and the atrial fibrillation promoting fibrotic substrate. Cardiovasc Res 113, 310–320 (2017).
Liu, Y. et al. Transforming growth factor-β (TGF-β)-mediated connective tissue growth factor (CTGF) expression in hepatic stellate cells requires Stat3 signaling activation. J. Biol. Chem. 288, 30708–30719 (2013).
Yang, Y. Z. et al. Magnesium isoglycyrrhizinate ameliorates high fructose-induced liver fibrosis in rat by increasing miR-375-3p to suppress JAK2/STAT3 pathway and TGF-β1/Smad signaling. Acta Pharm. Sin. 40, 879–894 (2019).
Xu, W. & Kimelman, D. Mechanistic insights from structural studies of beta-catenin and its binding partners. J. Cell Sci. 120, 3337–3344 (2007).
Murillo-Garzón, V. & Kypta, R. WNT signalling in prostate cancer. Nat. Rev. Urol. 14, 683–696 (2017).
Zhou, B. et al. Interactions between β-catenin and transforming growth factor-β signaling pathways mediate epithelial-mesenchymal transition and are dependent on the transcriptional co-activator cAMP-response element-binding protein (CREB)-binding protein (CBP). J. Biol. Chem. 287, 7026–7038 (2012).
Newman, D. R. et al. Expression of WNT5A in Idiopathic Pulmonary Fibrosis and Its Control by TGF-beta and WNT7B in Human Lung Fibroblasts. J. Histochem Cytochem 64, 99–111 (2016).
Ng-Blichfeldt, J. P. et al. TGF-β activation impairs fibroblast ability to support adult lung epithelial progenitor cell organoid formation. Am. J. Physiol. Lung Cell Mol. Physiol. 317, L14–l28 (2019).
Newman, D. R. et al. Expression of WNT5A in Idiopathic Pulmonary Fibrosis and Its Control by TGF-β and WNT7B in Human Lung Fibroblasts. J. Histochem Cytochem 64, 99–111 (2016).
Mackinnon, A. C. et al. Regulation of transforming growth factor-β1-driven lung fibrosis by galectin-3. Am. J. Respir. Crit. Care Med 185, 537–546 (2012).
Beljaars, L. et al. WNT-5A regulates TGF-β-related activities in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 312, G219–g227 (2017).
Mercurio, S. et al. Connective-tissue growth factor modulates WNT signalling and interacts with the WNT receptor complex. Development 131, 2137–2147 (2004).
Lawan, A. et al. Hepatic mitogen-activated protein kinase phosphatase 1 selectively regulates glucose metabolism and energy homeostasis. Mol. Cell Biol. 35, 26–40 (2015).
Vernia, S. et al. The PPARα-FGF21 hormone axis contributes to metabolic regulation by the hepatic JNK signaling pathway. Cell Metab. 20, 512–525 (2014).
Xiang, M. et al. Targeting hepatic TRAF1-ASK1 signaling to improve inflammation, insulin resistance, and hepatic steatosis. J. Hepatol. 64, 1365–1377 (2016).
Xie, L. et al. DKK3 expression in hepatocytes defines susceptibility to liver steatosis and obesity. J. Hepatol. 65, 113–124 (2016).
Schmelzer, C. E. H. et al. Lysyl oxidase-like 2 (LOXL2)-mediated cross-linking of tropoelastin. FASEB J. 33, 5468–5481 (2019).
Siddikuzzaman, Grace, V. M. & Guruvayoorappan, C. Lysyl oxidase: a potential target for cancer therapy. Inflammopharmacology 19, 117–129 (2011).
Vallet, S. D. & Ricard-Blum, S. Lysyl oxidases: from enzyme activity to extracellular matrix cross-links. Essays Biochem 63, 349–364 (2019).
Tenti, P. & Vannucci, L. Lysyl oxidases: linking structures and immunity in the tumor microenvironment. Cancer Immunol. Immunother. 69, 223–235 (2020).
Trackman, P. C. Lysyl Oxidase Isoforms and Potential Therapeutic Opportunities for Fibrosis and Cancer. Expert Opin. Ther. Targets 20, 935–945 (2016).
Ma, L. et al. Knockdown of LOXL1 inhibits TGF-β1-induced proliferation and fibrogenesis of hepatic stellate cells by inhibition of Smad2/3 phosphorylation. Biomed. Pharmacother. 107, 1728–1735 (2018).
Bellaye, P. S. et al. Lysyl Oxidase-Like 1 Protein Deficiency Protects Mice from Adenoviral Transforming Growth Factor-β1-induced Pulmonary Fibrosis. Am. J. Respir. Cell Mol. Biol. 58, 461–470 (2018).
Wen, X. et al. LOXL2, a copper-dependent monoamine oxidase, activates lung fibroblasts through the TGF-β/Smad pathway. Int J. Mol. Med 42, 3530–3541 (2018).
Kersten, S., Desvergne, B. & Wahli, W. Roles of PPARs in health and disease. Nature 405, 421–424 (2000).
Desvergne, B. & Wahli, W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocr. Rev. 20, 649–688 (1999).
Ahmadian, M. et al. PPARγ signaling and metabolism: the good, the bad and the future. Nat. Med 19, 557–566 (2013).
Cheng, H. S. et al. Exploration and Development of PPAR Modulators in Health and Disease: An Update of Clinical Evidence. Int. J. Mol. Sci. 20, 5055 (2019).
Monsalve, F. A., Pyarasani, R. D., Delgado-Lopez, F. & Moore-Carrasco, R. Peroxisome proliferator-activated receptor targets for the treatment of metabolic diseases. Mediators Inflamm. 2013, 549627 (2013).
Dubois, V., Eeckhoute, J., Lefebvre, P. & Staels, B. Distinct but complementary contributions of PPAR isotypes to energy homeostasis. J. Clin. Invest 127, 1202–1214 (2017).
Pawlak, M., Lefebvre, P. & Staels, B. Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 62, 720–733 (2015).
Abdelmegeed, M. A. et al. PPARalpha expression protects male mice from high fat-induced nonalcoholic fatty liver. J. Nutr. 141, 603–610 (2011).
Toyama, T. et al. PPARalpha ligands activate antioxidant enzymes and suppress hepatic fibrosis in rats. Biochem Biophys. Res Commun. 324, 697–704 (2004).
Montagner, A. et al. Liver PPARα is crucial for whole-body fatty acid homeostasis and is protective against NAFLD. Gut 65, 1202–1214 (2016).
Konstantinopoulos, P. A. et al. NF-kappaB/PPAR gamma and/or AP-1/PPAR gamma ‘on/off’ switches and induction of CBP in colon adenocarcinomas: correlation with COX-2 expression. Int J. Colorectal Dis. 22, 57–68 (2007).
Bouhlel, M. A. et al. PPARgamma activation primes human monocytes into alternative M2 macrophages with anti-inflammatory properties. Cell Metab. 6, 137–143 (2007).
Abd El-Haleim, E. A., Bahgat, A. K. & Saleh, S. Effects of combined PPAR-γ and PPAR-α agonist therapy on fructose induced NASH in rats: Modulation of gene expression. Eur. J. Pharm. 773, 59–70 (2016).
Jain, M. R. et al. Dual PPARα/γ agonist saroglitazar improves liver histopathology and biochemistry in experimental NASH models. Liver Int 38, 1084–1094 (2018).
Iglarz, M. et al. Peroxisome proliferator-activated receptor-alpha and receptor-gamma activators prevent cardiac fibrosis in mineralocorticoid-dependent hypertension. Hypertension 42, 737–743 (2003).
Kawai, T. et al. PPAR-gamma agonist attenuates renal interstitial fibrosis and inflammation through reduction of TGF-beta. Lab Invest 89, 47–58 (2009).
Kulkarni, A. A. et al. PPAR-γ ligands repress TGFβ-induced myofibroblast differentiation by targeting the PI3K/Akt pathway: implications for therapy of fibrosis. PLoS One 6, e15909 (2011).
Liss, K. H. & Finck, B. N. PPARs and nonalcoholic fatty liver disease. Biochimie 136, 65–74 (2017).
Girroir, E. E. et al. Quantitative expression patterns of peroxisome proliferator-activated receptor-beta/delta (PPARbeta/delta) protein in mice. Biochem Biophys. Res Commun. 371, 456–461 (2008).
Shan, W. et al. Peroxisome proliferator-activated receptor-beta/delta protects against chemically induced liver toxicity in mice. Hepatology 47, 225–235 (2008).
Hellemans, K. et al. Peroxisome proliferator-activated receptor-beta signaling contributes to enhanced proliferation of hepatic stellate cells. Gastroenterology 124, 184–201 (2003).
Kostadinova, R. et al. GW501516-activated PPARβ/δ promotes liver fibrosis via p38-JNK MAPK-induced hepatic stellate cell proliferation. Cell Biosci. 2, 34 (2012).
Iwaisako, K. et al. Protection from liver fibrosis by a peroxisome proliferator-activated receptor δ agonist. Proc. Natl Acad. Sci. USA 109, E1369–E1376 (2012).
Matsubara, T., Li, F. & Gonzalez, F. J. FXR signaling in the enterohepatic system. Mol. Cell Endocrinol. 368, 17–29 (2013).
Wang, H. et al. FXR modulators for enterohepatic and metabolic diseases. Expert Opin. Ther. Pat. 28, 765–782 (2018).
Kim, K. H. et al. Hepatic FXR/SHP axis modulates systemic glucose and fatty acid homeostasis in aged mice. Hepatology 66, 498–509 (2017).
Laffitte, B. A. et al. Identification of the DNA binding specificity and potential target genes for the farnesoid X-activated receptor. J. Biol. Chem. 275, 10638–10647 (2000).
Chen, B. et al. Bile acids induce activation of alveolar epithelial cells and lung fibroblasts through farnesoid X receptor-dependent and independent pathways. Respirology 21, 1075–1080 (2016).
Wang, X. X. et al. The farnesoid X receptor modulates renal lipid metabolism and diet-induced renal inflammation, fibrosis, and proteinuria. Am. J. Physiol. Ren. Physiol. 297, F1587–F1596 (2009).
Han, C. Y. et al. FXR Inhibits Endoplasmic Reticulum Stress-Induced NLRP3 Inflammasome in Hepatocytes and Ameliorates Liver Injury. Cell Rep. 24, 2985–2999 (2018).
Kong, B., Luyendyk, J. P., Tawfik, O. & Guo, G. L. Farnesoid X receptor deficiency induces nonalcoholic steatohepatitis in low-density lipoprotein receptor-knockout mice fed a high-fat diet. J. Pharm. Exp. Ther. 328, 116–122 (2009).
Kok, T. et al. Induction of hepatic ABC transporter expression is part of the PPARalpha-mediated fasting response in the mouse. Gastroenterology 124, 160–171 (2003).
Holt, J. A. et al. Definition of a novel growth factor-dependent signal cascade for the suppression of bile acid biosynthesis. Genes Dev. 17, 1581–1591 (2003).
Kir, S. et al. FGF19 as a postprandial, insulin-independent activator of hepatic protein and glycogen synthesis. Science 331, 1621–1624 (2011).
Cyphert, H. A. et al. Activation of the farnesoid X receptor induces hepatic expression and secretion of fibroblast growth factor 21. J. Biol. Chem. 287, 25123–25138 (2012).
Al-Aqil, F. A. et al. Interaction of glucocorticoids with FXR/FGF19/FGF21-mediated ileum-liver crosstalk. Biochim Biophys. Acta Mol. Basis Dis. 1864, 2927–2937 (2018).
Bhattacharyya, S. et al. TLR4-dependent fibroblast activation drives persistent organ fibrosis in skin and lung. JCI Insight. 3, e98850 (2018).
Seki, E. et al. TLR4 enhances TGF-beta signaling and hepatic fibrosis. Nat. Med 13, 1324–1332 (2007).
Das, S. et al. NADPH Oxidase-Derived Peroxynitrite Drives Inflammation in Mice and Human Nonalcoholic Steatohepatitis via TLR4-Lipid Raft Recruitment. Am. J. Pathol. 185, 1944–1957 (2015).
Sutter, A. G. et al. Dietary Saturated Fat Promotes Development of Hepatic Inflammation Through Toll-Like Receptor 4 in Mice. J. Cell Biochem 117, 1613–1621 (2016).
Liang, J. et al. Hyaluronan and TLR4 promote surfactant-protein-C-positive alveolar progenitor cell renewal and prevent severe pulmonary fibrosis in mice. Nat. Med 22, 1285–1293 (2016).
Gasbjerg, L. S. et al. GIP and GLP-1 Receptor Antagonism During a Meal in Healthy Individuals. J. Clin. Endocrinol. Metab. 105, dgz175 (2020).
Smith, N. K., Hackett, T. A., Galli, A. & Flynn, C. R. GLP-1: Molecular mechanisms and outcomes of a complex signaling system. Neurochem Int 128, 94–105 (2019).
Baggio, L. L. & Drucker, D. J. Biology of incretins: GLP-1 and GIP. Gastroenterology 132, 2131–2157 (2007).
Seino, Y., Fukushima, M. & Yabe, D. GIP and GLP-1, the two incretin hormones: Similarities and differences. J. Diabetes Investig. 1, 8–23 (2010).
Gabe, M. B. N. et al. Molecular interactions of full-length and truncated GIP peptides with the GIP receptor - A comprehensive review. Peptides 125, 170224 (2020).
Adriaenssens, A. E. et al. Glucose-Dependent Insulinotropic Polypeptide Receptor-Expressing Cells in the Hypothalamus Regulate Food Intake. Cell Metab. 30, 987–-996.e986 (2019).
Kim, S. J. et al. GIP-overexpressing mice demonstrate reduced diet-induced obesity and steatosis, and improved glucose homeostasis. PLoS One 7, e40156 (2012).
Chen, S., Okahara, F., Osaki, N. & Shimotoyodome, A. Increased GIP signaling induces adipose inflammation via a HIF-1α-dependent pathway and impairs insulin sensitivity in mice. Am. J. Physiol. Endocrinol. Metab. 308, E414–E425 (2015).
Drucker, D. J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 27, 740–756 (2018).
Valdecantos, M. P., Ruiz, L., Pardo, V. & Castro-Sanchez, L. Differential Effects of a Glucagon-Like Peptide 1 Receptor Agonist in Non-Alcoholic Fatty Liver Disease and in Response to Hepatectomy. Sci. Rep. 8, 16461 (2018).
Trevaskis, J. L. et al. Glucagon-like peptide-1 receptor agonism improves metabolic, biochemical, and histopathological indices of nonalcoholic steatohepatitis in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 302, G762–G772 (2012).
Liu, J. et al. Glucagon-like peptide-1 (GLP-1) improved diabetic lung fibrosis via AMPK and microRNA-27a (miR-27a). Ann. Transl. Med 9, 492 (2021).
Kannt, A., Madsen, A. N., Kammermeier, C. & Elvert, R. Incretin combination therapy for the treatment of non-alcoholic steatohepatitis. Diabetes Obes. Metab. 22, 1328–1338 (2020).
Derynck, R., Turley, S. J. & Akhurst, R. J. TGFβ biology in cancer progression and immunotherapy. Nat. Rev. Clin. Oncol. 18, 9–34 (2021).
Ong, C. H. et al. TGF-β-induced fibrosis: A review on the underlying mechanism and potential therapeutic strategies. Eur. J. Pharm. 911, 174510 (2021).
Neighbors, M. et al. Prognostic and predictive biomarkers for patients with idiopathic pulmonary fibrosis treated with pirfenidone: post-hoc assessment of the CAPACITY and ASCEND trials. Lancet Respir. Med 6, 615–626 (2018).
Antoniu, S. A. Pirfenidone for the treatment of idiopathic pulmonary fibrosis. Expert Opin. Investig. Drugs 15, 823–828 (2006).
Molina-Molina, M. et al. Anti-fibrotic effects of pirfenidone and rapamycin in primary IPF fibroblasts and human alveolar epithelial cells. BMC Pulm. Med 18, 63 (2018).
Lehmann, M. et al. Differential effects of Nintedanib and Pirfenidone on lung alveolar epithelial cell function in ex vivo murine and human lung tissue cultures of pulmonary fibrosis. Respir. Res 19, 175 (2018).
Epstein Shochet, G., Wollin, L. & Shitrit, D. Fibroblast-matrix interplay: Nintedanib and pirfenidone modulate the effect of IPF fibroblast-conditioned matrix on normal fibroblast phenotype. Respirology 23, 756–763 (2018).
Knüppel, L. et al. A Novel Antifibrotic Mechanism of Nintedanib and Pirfenidone. Inhibition of Collagen Fibril Assembly. Am. J. Respir. Cell Mol. Biol. 57, 77–90 (2017).
Iwata, T. et al. A phase II trial evaluating the efficacy and safety of perioperative pirfenidone for prevention of acute exacerbation of idiopathic pulmonary fibrosis in lung cancer patients undergoing pulmonary resection: West Japan Oncology Group 6711 L (PEOPLE Study). Respir. Res 17, 90 (2016).
Vancheri, C. et al. Nintedanib with Add-on Pirfenidone in Idiopathic Pulmonary Fibrosis. Results of the INJOURNEY Trial. Am. J. Respir. Crit. Care Med 197, 356–363 (2018).
Yoon, H. Y., Kim, D. S. & Song, J. W. Efficacy and Safety of Pirfenidone in Advanced Idiopathic Pulmonary Fibrosis. Respiration 97, 242–251 (2019).
Zurkova, M. & Kriegova, E. Effect of pirfenidone on lung function decline and survival: 5-yr experience from a real-life IPF cohort from the Czech EMPIRE registry. Respir. Res 20, 16 (2019).
Noble, P. W. et al. Pirfenidone in patients with idiopathic pulmonary fibrosis (CAPACITY): two randomised trials. Lancet 377, 1760–1769 (2011).
Barratt, S. L. et al. South-West of England’s Experience of the Safety and Tolerability Pirfenidone and Nintedanib for the Treatment of Idiopathic Pulmonary Fibrosis (IPF). Front Pharm. 9, 1480 (2018).
Liu, Y. et al. Tolerability and Pharmacokinetics of Hydronidone, an Antifibrotic Agent for Hepatic Fibrosis, after Single and Multiple Doses in Healthy Subjects: an Open-Label, Randomized, Dose-Escalating, First-in-Human Study. Eur. J. Drug Metab. Pharmacokinet. 42, 37–48 (2017).
Tremblay, D. & Mascarenhas, J. Next Generation Therapeutics for the Treatment of Myelofibrosis. Cells. 10, 1034 (2021).
Varricchio, L. et al. TGF-β1 protein trap AVID200 beneficially affects hematopoiesis and bone marrow fibrosis in myelofibrosis. JCI Insight. 6, 1–8 (2021).
Rivera-Ortega, P. et al. Nintedanib in the management of idiopathic pulmonary fibrosis: clinical trial evidence and real-world experience. Ther. Adv. Respir. Dis. 12, 1753466618800618 (2018).
Wollin, L. et al. Antifibrotic and anti-inflammatory activity of the tyrosine kinase inhibitor nintedanib in experimental models of lung fibrosis. J. Pharm. Exp. Ther. 349, 209–220 (2014).
Brown, K. K. et al. Lung function outcomes in the INPULSIS(®) trials of nintedanib in idiopathic pulmonary fibrosis. Respir. Med 146, 42–48 (2019).
Richeldi, L. et al. Efficacy and safety of nintedanib in idiopathic pulmonary fibrosis. N. Engl. J. Med 370, 2071–2082 (2014).
Yoon, H. Y., Park, S., Kim, D. S. & Song, J. W. Efficacy and safety of nintedanib in advanced idiopathic pulmonary fibrosis. Respir. Res 19, 203 (2018).
Wuyts, W. A. et al. First Data on Efficacy and Safety of Nintedanib in Patients with Idiopathic Pulmonary Fibrosis and Forced Vital Capacity of ≤50 % of Predicted Value. Lung 194, 739–743 (2016).
Kolb, M., Richeldi, L., Behr, J. & Maher, T. M. Nintedanib in patients with idiopathic pulmonary fibrosis and preserved lung volume. Thorax 72, 340–346 (2017).
Pereira, C. A. C. & Baddini-Martinez, J. A. Safety and tolerability of nintedanib in patients with idiopathic pulmonary fibrosis in Brazil. J. Bras. Pneumol. 45, e20180414 (2019).
Crestani, B. et al. Long-term safety and tolerability of nintedanib in patients with idiopathic pulmonary fibrosis: results from the open-label extension study, INPULSIS-ON. Lancet Respir. Med 7, 60–68 (2019).
Kato, M. et al. Gastrointestinal adverse effects of nintedanib and the associated risk factors in patients with idiopathic pulmonary fibrosis. Sci. Rep. 9, 12062 (2019).
Yao, Y. et al. Design, synthesis and pharmacological evaluation of 4-(3-chloro-4-(3-cyclopropylthioureido)-2-fluorophenoxy)-7-methoxyquinoline-6-carboxamide (WXFL-152): a novel triple angiokinase inhibitor for cancer therapy. Acta Pharmaceutica Sin. B 10, 1453–1475 (2020).
Liu, Z. W. et al. The antifibrotic effect and mechanism of a novel tyrosine kinase inhibitor, ZSP1603, in preclinical models of pulmonary fibrosis. Eur. Rev. Med Pharm. Sci. 24, 1481–1491 (2020).
Richeldi, L. et al. Pamrevlumab, an anti-connective tissue growth factor therapy, for idiopathic pulmonary fibrosis (PRAISE): a phase 2, randomised, double-blind, placebo-controlled trial. Lancet Respiratory Med. 8, 25–33 (2020).
Shin, N. et al. Parsaclisib Is a Next-Generation Phosphoinositide 3-Kinase δ Inhibitor with Reduced Hepatotoxicity and Potent Antitumor and Immunomodulatory Activities in Models of B-Cell Malignancy. J. Pharm. Exp. Ther. 374, 211–222 (2020).
Li, J. et al. An Open-Label Study to Assess the Effect of Itraconazole and Rifampin on Parsaclisib Pharmacokinetics When Administered Orally in Healthy Participants. J. Clin. Pharmacol. 60, 1519–1526 (2020).
Lukey, P. T. et al. A randomised, placebo-controlled study of omipalisib (PI3K/mTOR) in idiopathic pulmonary fibrosis. Eur. Respir. J. 53, 1801992 (2019).
Mylonas, E. et al. Single-cell analysis based dissection of clonality in myelofibrosis. Nat. Commun. 11, 73 (2020).
Cervantes, F. et al. Efficacy and safety of a novel dosing strategy for ruxolitinib in the treatment of patients with myelofibrosis and anemia: the REALISE phase 2 study. Leukemia 35, 3455–3465 (2021).
Tyner, J. W. et al. CYT387, a novel JAK2 inhibitor, induces hematologic responses and normalizes inflammatory cytokines in murine myeloproliferative neoplasms. Blood 115, 5232–5240 (2010).
Harrison, C. N. et al. Momelotinib versus best available therapy in patients with myelofibrosis previously treated with ruxolitinib (SIMPLIFY 2): a randomised, open-label, phase 3 trial. Lancet Haematol. 5, e73–e81 (2018).
Mesa, R. A. et al. SIMPLIFY-1: A Phase III Randomized Trial of Momelotinib Versus Ruxolitinib in Janus Kinase Inhibitor-Naïve Patients With Myelofibrosis. J. Clin. Oncol. 35, 3844–3850 (2017).
Talpaz, M. & Kiladjian, J. J. Fedratinib, a newly approved treatment for patients with myeloproliferative neoplasm-associated myelofibrosis. Leukemia 35, 1–17 (2021).
Pardanani, A. et al. Updated results of the placebo-controlled, phase III JAKARTA trial of fedratinib in patients with intermediate-2 or high-risk myelofibrosis. Br. J. Haematol. 195, 244–248 (2021).
Verstovsek, S. et al. Phase 1/2 study of pacritinib, a next generation JAK2/FLT3 inhibitor, in myelofibrosis or other myeloid malignancies. J. Hematol. Oncol. 9, 137 (2016).
Komrokji, R. S. et al. Results of a phase 2 study of pacritinib (SB1518), a JAK2/JAK2(V617F) inhibitor, in patients with myelofibrosis. Blood 125, 2649–2655 (2015).
Mascarenhas, J. et al. Pacritinib vs Best Available Therapy, Including Ruxolitinib, in Patients With Myelofibrosis: A Randomized Clinical Trial. JAMA Oncol. 4, 652–659 (2018).
Beatty, G. L. et al. A Phase Ib/II Study of the JAK1 Inhibitor, Itacitinib, plus nab-Paclitaxel and Gemcitabine in Advanced Solid Tumors. Oncologist 24, 14–e10 (2019).
Lescoat, A. et al. Combined anti-fibrotic and anti-inflammatory properties of JAK-inhibitors on macrophages in vitro and in vivo: Perspectives for scleroderma-associated interstitial lung disease. Biochem Pharm. 178, 114103 (2020).
Mascarenhas, J. O. et al. Primary analysis of a phase II open-label trial of INCB039110, a selective JAK1 inhibitor, in patients with myelofibrosis. Haematologica 102, 327–335 (2017).
Eguchi, M., Nguyen, C., Lee, S. C. & Kahn, M. ICG-001, a novel small molecule regulator of TCF/beta-catenin transcription. Med Chem. 1, 467–472 (2005).
Akcora, B. O., Storm, G. & Bansal, R. Inhibition of canonical WNT signaling pathway by beta-catenin/CBP inhibitor ICG-001 ameliorates liver fibrosis in vivo through suppression of stromal CXCL12. Biochim Biophys. Acta Mol. Basis Dis. 1864, 804–818 (2018).
Tokunaga, Y. et al. Selective inhibitor of Wnt/β-catenin/CBP signaling ameliorates hepatitis C virus-induced liver fibrosis in mouse model. Sci. Rep. 7, 325 (2017).
Kimura, K. et al. Safety, Tolerability, and Preliminary Efficacy of the Anti-Fibrotic Small Molecule PRI-724, a CBP/beta-Catenin Inhibitor, in Patients with Hepatitis C Virus-related Cirrhosis: A Single-Center, Open-Label, Dose Escalation Phase 1 Trial. EBioMedicine 23, 79–87 (2017).
Kimura, K. et al. Safety, tolerability, and anti-fibrotic efficacy of the CBP/β-catenin inhibitor PRI-724 in patients with hepatitis C and B virus-induced liver cirrhosis: An investigator-initiated, open-label, non-randomised, multicentre, phase 1/2a study. EBioMedicine 80, 104069 (2022).
Loomba, R. et al. The ASK1 inhibitor selonsertib in patients with nonalcoholic steatohepatitis: A randomized, phase 2 trial. Hepatology 67, 549–559 (2018).
Harrison, S. A. et al. Selonsertib for patients with bridging fibrosis or compensated cirrhosis due to NASH: Results from randomized phase III STELLAR trials. J. Hepatol. 73, 26–39 (2020).
Li, X. et al. Treatment with PPARδ agonist alleviates non-alcoholic fatty liver disease by modulating glucose and fatty acid metabolic enzymes in a rat model. Int J. Mol. Med 36, 767–775 (2015).
Westerouen Van Meeteren, M. J., Drenth, J. P. H. & Tjwa, E. Elafibranor: a potential drug for the treatment of nonalcoholic steatohepatitis (NASH). Expert Opin. Investig. Drugs 29, 117–123 (2020).
Liu, L. et al. The pharmacodynamic and differential gene expression analysis of PPAR α/δ agonist GFT505 in CDAHFD-induced NASH model. PLoS One 15, e0243911 (2020).
Ratziu, V. et al. Elafibranor, an Agonist of the Peroxisome Proliferator-Activated Receptor-α and -δ, Induces Resolution of Nonalcoholic Steatohepatitis Without Fibrosis Worsening. Gastroenterology 150, 1147–1159.e1145 (2016).
Gawrieh, S. et al. Saroglitazar, a PPAR-α/γ Agonist, for Treatment of NAFLD: A Randomized Controlled Double-Blind Phase 2 Trial. Hepatology 74, 1809–1824 (2021).
Lefere, S. et al. Differential effects of selective- and pan-PPAR agonists on experimental steatohepatitis and hepatic macrophages(☆). J. Hepatol. 73, 757–770 (2020).
Francque, S. M. et al. A Randomized, Controlled Trial of the Pan-PPAR Agonist Lanifibranor in NASH. N. Engl. J. Med 385, 1547–1558 (2021).
Honda, Y. et al. Pemafibrate, a novel selective peroxisome proliferator-activated receptor alpha modulator, improves the pathogenesis in a rodent model of nonalcoholic steatohepatitis. Sci. Rep. 7, 42477 (2017).
Sasaki, Y. et al. Pemafibrate, a selective PPARα modulator, prevents non-alcoholic steatohepatitis development without reducing the hepatic triglyceride content. Sci. Rep. 10, 7818 (2020).
Nakajima, A. et al. Randomised clinical trial: Pemafibrate, a novel selective peroxisome proliferator-activated receptor α modulator (SPPARMα), versus placebo in patients with non-alcoholic fatty liver disease. Aliment Pharm. Ther. 54, 1263–1277 (2021).
Younossi, Z. M. et al. Obeticholic acid for the treatment of non-alcoholic steatohepatitis: interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial. Lancet 394, 2184–2196 (2019).
Patel, K. et al. Cilofexor, a Nonsteroidal FXR Agonist, in Patients With Noncirrhotic NASH: A Phase 2 Randomized Controlled Trial. Hepatology 72, 58–71 (2020).
An, P., Wei, G. & Huang, P. A novel non-bile acid FXR agonist EDP-305 potently suppresses liver injury and fibrosis without worsening of ductular reaction. Liver Int. 40, 1655–1669 (2020).
Ratziu, V. et al. EDP-305 in patients with NASH: A phase II double-blind placebo-controlled dose-ranging study. J. Hepatol. 76, 506–517 (2022).
Hernandez, E. D. et al. Tropifexor-Mediated Abrogation of Steatohepatitis and Fibrosis Is Associated With the Antioxidative Gene Expression Profile in Rodents. Hepatol. Commun. 3, 1085–1097 (2019).
Badman, M. K. et al. Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of the Novel Non-Bile Acid FXR Agonist Tropifexor (LJN452) in Healthy Volunteers. Clin. Pharm. Drug Dev. 9, 395–410 (2020).
Sumida, Y. & Yoneda, M. Current and future pharmacological therapies for NAFLD/NASH. J. Gastroenterol. 53, 362–376 (2018).
Hartman, M. L. et al. Effects of Novel Dual GIP and GLP-1 Receptor Agonist Tirzepatide on Biomarkers of Nonalcoholic Steatohepatitis in Patients With Type 2 Diabetes. Diabetes Care 43, 1352–1355 (2020).
Frias, J. P. et al. Efficacy and safety of LY3298176, a novel dual GIP and GLP-1 receptor agonist, in patients with type 2 diabetes: a randomised, placebo-controlled and active comparator-controlled phase 2 trial. Lancet 392, 2180–2193 (2018).
Newsome, P. N. et al. A Placebo-Controlled Trial of Subcutaneous Semaglutide in Nonalcoholic Steatohepatitis. N. Engl. J. Med 384, 1113–1124 (2021).
Ambery, P. D. et al. MEDI0382, a GLP-1/glucagon receptor dual agonist, meets safety and tolerability endpoints in a single-dose, healthy-subject, randomized, Phase 1 study. Br. J. Clin. Pharm. 84, 2325–2335 (2018).
Boland, M. L. et al. Resolution of NASH and hepatic fibrosis by the GLP-1R/GcgR dual-agonist Cotadutide via modulating mitochondrial function and lipogenesis. Nat. Metab. 2, 413–431 (2020).
Goor, F. V., Hadida, S. & Grootenhuis, P. Pharmacological Rescue of Mutant CFTR Function for the Treatment of Cystic Fibrosis. (Ion Channels, 2008).
Jih, K. Y., Lin, W. Y., Sohma, Y. & Hwang, T. C. CFTR potentiators: from bench to bedside. Curr. Opin. Pharm. 34, 98–104 (2017).
Rowe, S. M. et al. Tezacaftor-Ivacaftor in Residual-Function Heterozygotes with Cystic Fibrosis. N. Engl. J. Med 377, 2024–2035 (2017).
Donaldson, S. H. et al. Tezacaftor/Ivacaftor in Subjects with Cystic Fibrosis and F508del/F508del-CFTR or F508del/G551D-CFTR. Am. J. Respir. Crit. Care Med 197, 214–224 (2018).
Connett, G. J. Lumacaftor-ivacaftor in the treatment of cystic fibrosis: design, development and place in therapy. Drug Des. Devel Ther. 13, 2405–2412 (2019).
Milla, C. E. et al. Lumacaftor/Ivacaftor in Patients Aged 6-11 Years with Cystic Fibrosis and Homozygous for F508del-CFTR. Am. J. Respir. Crit. Care Med 195, 912–920 (2017).
Wainwright, C. E. et al. Lumacaftor-Ivacaftor in Patients with Cystic Fibrosis Homozygous for Phe508del CFTR. N. Engl. J. Med 373, 220–231 (2015).
Wang, X. et al. Discovery of 4-[(2R,4R)-4-({[1-(2,2-Difluoro-1,3-benzodioxol-5-yl)cyclopropyl]carbonyl}amino)-7-(difluoromethoxy)-3,4-dihydro-2H-chromen-2-yl]benzoic Acid (ABBV/GLPG-2222), a Potent Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Corrector for the Treatment of Cystic Fibrosis. J. Med. Chem. 61, 1436–1449 (2018).
Bell, S. C. et al. CFTR activity is enhanced by the novel corrector GLPG2222, given with and without ivacaftor in two randomized trials. J. Cyst. Fibros. 18, 700–707 (2019).
Kim, J. et al. Metabolite Profiling of the Antisense Oligonucleotide Eluforsen Using Liquid Chromatography-Mass Spectrometry. Mol. Ther. Nucleic Acids 17, 714–725 (2019).
Drevinek, P. et al. Antisense oligonucleotide eluforsen is safe and improves respiratory symptoms in F508DEL cystic fibrosis. J. Cyst. Fibros. 19, 99–107 (2020).
Acknowledgements
This work was supported by the National Science Foundation of China (No. 81773375) and the National Major Scientific and Technological Special Project (No. 2019ZX09201001). The authors thank the Research Center for Prevention and Therapy of Occupational Diseases, West China-PUMC C.C.Chen Institute of Health, Sichuan University.
Author information
Authors and Affiliations
Contributions
Y.Y. and J.Y. designed the research. M.Z., M.W., S.Z., and Y.L. searched and analyzed the papers. M.Z. and L.W. wrote the initial manuscript. M.Z. and Y.Y. prepared the figures. A.C.R., L.Z., B.D., T.Y., H.C., and B.Z. critically reviewed and revised the final manuscript. All the authors have read and approved the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhao, M., Wang, L., Wang, M. et al. Targeting fibrosis: mechanisms and clinical trials. Sig Transduct Target Ther 7, 206 (2022). https://doi.org/10.1038/s41392-022-01070-3
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41392-022-01070-3
This article is cited by
-
Hygrothermal stress increases malignant arrhythmias susceptibility by inhibiting the LKB1-AMPK-Cx43 pathway
Scientific Reports (2024)
-
A two-way street – cellular metabolism and myofibroblast contraction
npj Regenerative Medicine (2024)
-
Identification of non-coding RNA signatures in idiopathic pulmonary fibrosis
Irish Journal of Medical Science (1971 -) (2024)
-
AMPK signaling inhibits the differentiation of myofibroblasts: impact on age-related tissue fibrosis and degeneration
Biogerontology (2024)
-
Connective tissue disorders in COVID-19: Reply to “People with a connective tissue disorder may be especially vulnerable to the endothelial damage that characterizes long COVID due to the fragility of their vasculature and slow wound healing”
Angiogenesis (2024)
