
Abstract
Racial discrimination (RD) has been consistently linked to adverse brain health outcomes. These may be due in part to RD effects on neural networks involved with threat appraisal and regulation; RD has been linked to altered activity in the rostral anterior cingulate cortex (rACC) and structural decrements in the anterior cingulum bundle and hippocampus. In the present study, we examined associations of RD with cingulate, hippocampus and amygdala gray matter morphology in a sample of trauma-exposed Black women. Eighty-one Black women aged 19–62 years were recruited as part of an ongoing study of trauma. Participants completed assessments of RD, trauma exposure, and posttraumatic stress disorder (PTSD), and underwent T1-weighted anatomical imaging. Cortical thickness, surface area and gray matter volume were extracted from subregions of cingulate cortex, and gray matter volume was extracted from amygdala and hippocampus, and entered into partial correlation analyses that included RD and other socio-environmental variables. After correction for multiple comparisons and accounting for variance associated with other stressors and socio-environmental factors, participants with more RD exposure showed proportionally lower cortical thickness in the left rACC, caudal ACC, and posterior cingulate cortex (ps < = 0.01). These findings suggest that greater experiences of RD are linked to compromised cingulate gray matter thickness. In the context of earlier findings indicating that RD produces increased response in threat neurocircuitry, our data suggest that RD may increase vulnerability for brain health problems via cingulate cortex alterations. Further research is needed to elucidate biological mechanisms for these changes.
Similar content being viewed by others

Introduction
Racial discrimination (RD) is a chronic stressor faced by a majority of Black individuals [1, 2]. The mental and physical burden of RD is clear through a substantial body of research indicating strong links with poorer mental, cardiovascular, and brain health [3]; for example, RD has been linked to disproportionately higher risk for the development of anxiety, depression and PTSD [4] and has been linked to incidence of vascular disease and cerebrovascular events, as well as earlier cognitive decline [5,6,7]. Experiences of RD, which are characterized by social exclusion, hostile and unjust treatment on the basis of race or ethnicity, can live on through persistent negative ruminations about the nature of event and mental preparation for future insults [8]. Further, such racist injuries can activate the body’s stress response system (hypothalamic-pituitary-adrenal axis) [9], and immune system [10] similar to other types of psychological stress and trauma [11]. More frequent exposure to racism-related stress takes a direct toll on overall health [11]. The ever-growing amount of research on the role of racism on numerous adverse mental and physical health outcomes has precipitated recognition of RD as a national public health crisis[12].
Despite the corpus of evidence linking RD to poor health, overall distress and disruptions in autonomic regulation [13, 14], research on the neuropathophysiology of RD—that is, the effects of this type of discrimination on brain and physiological responses—is in its nascency. Data from functional neuroimaging studies indicate that RD is associated with increased threat-cue reactivity in, and connectivity between, brain regions such as the amygdala, hippocampus, and cingulate cortex, regions that consistently demonstrate neuroplastic changes in response to chronic stress [15,16,17,18]. The amygdala and hippocampus facilitate encoding of biologically salient stimuli (e.g., threat-relevant cues) and are involved with the expression of defensive reflexes; subdivisions of the ACC modulate amygdala response [19]. The rostral ACC (rACC) supports response inhibition and emotion modulation, whereas the caudal ACC (cACC) is associated with threat detection and facilitation of amygdala-mediated threat responses [19]. Further, the posterior cingulate cortex (PCC) is a major functional hub and part of the default mode network in conjunction with the rACC [20, 21]; the PCC’s role in directing attentional focus as well as internally-directed cognition, (including rumination), has been consistently highlighted [22]. Abnormalities in PCC structure are apparent across a range of neurological and psychiatric disorders, and it is thought to be among one of the more susceptible structures to neurodegenerative processes [20]. Given the importance of these regions for cognitive and emotional processes and vulnerability to stress-related neuroplastic changes, the potential neurophysiological impact of RD on this circuitry is important to unravel.
Although RD may present as discrete, circumscribed events, over time these experiences have cumulative effects on minoritized individuals; RD is thought to produce long-lasting stress responses in the form of rumination, regular suppression of emotional response and/or enhanced vigilance for future threat [23]. RD is a unique stressor that often experienced on an ongoing basis, with a physiological signature that is distinct from other types of (focal) trauma exposure [2, 24]. This contributes to a context of prolonged stress on peripheral and central nervous systems; chronic stressors like RD are known to impact neural plasticity, particularly within the ACC, amygdala and hippocampus [18, 25, 26]. Indeed, chronic posttraumatic stress disorder (PTSD) has been characterized by reductions in ACC, hippocampal and amygdala volume [27,28,29]. The effects of chronic stress on cingulate, hippocampus and amygdala gray and white matter extend to RD. Our recent study showed that RD is a potent contributor to decrements in brain white matter microstructure in the anterior cingulum bundle, a pathway that is dorsal to the ACC and serves as a primary connection for this region [30], and a study of older adults indicated associations of RD with lower hippocampal volume and increases in hippocampal white matter hyperintensities over time [31]. RD has also shown global adverse effects on intracranial volume and white matter signals [32], with specific decrements observed in fronto-limbic and fronto-parietal white matter pathways [30]. It is possible that race-related adversity, including RD, could induce changes in neural plasticity, particularly within the ACC, hippocampus and amygdala, but to our knowledge, no studies to date have examined unique associations of RD with brain gray matter morphology, independent of other types of trauma exposure.
Given earlier findings indicating RD’s links to: increased function in rostral and ventral prefrontal regions [16, 17], decrements in white matter in the anterior cingulum bundle [30]; and increased reactivity in threat neurocircuitry [15], we assessed the relationship between RD and gray matter morphology for the ACC, amygdala and hippocampus in a sample of Black American women. We examined the unique associations of RD with gray matter architecture, including cortical thickness, surface area and gray matter volume, after controlling for variance associated with economic disadvantage, trauma exposure and PTSD, similar to prior studies [6, 16, 30], as well as intracranial volume (ICV). Secondarily, we examined gray matter volume for two subcortical structures that are implicated in studies of RD as well as trauma and stress-related disorders, the amygdala and hippocampus [15, 33]. Individuals were participants in the Grady Trauma Project (GTP), a large-scale, ongoing study of risk and resilience for trauma-related disorders. Given that prior GTP studies specifically recruited women (e.g., MH101380, MH111682, HD071982), only women were represented in this sample. RD was assessed through a self-report questionnaire, and responses on this measure were examined in association with gray matter metrics for the cingulate cortex (cortical thickness, surface area, and gray matter volume of rostral, caudal, and posterior segments), hippocampus and amygdala (gray matter volume). White matter data from 73 of these participants were published in our recent study of RD and white matter microstructure [30], which indicated significant associations of RD with the left anterior cingulum bundle; as such, we predicted that RD would be particularly linked to decrements in gray matter morphology in left cingulate regions. As with prior research, we examined the unique associations of RD with gray matter architecture after accounting for the effects of economic disadvantage, childhood and adulthood trauma exposure and PTSD [16, 30] as well as ICV. To assess for specificity of significant effects, we repeated analyses with other cortical regions (i.e., postcentral gyrus, cuneus).
Methods
Participants
Eighty-one Black American women aged 19–62 were recruited from the GTP, which represents a group of studies investigating risk and resilience for trauma-related disorders (MH101380, MH071537, MH094757). Individuals were approached in general medical clinics of a publicly-funded hospital in Atlanta, Georgia. Eligibility for participation included the ability to understand English (assessed by a study researcher) and willingness to provide informed consent. Exclusion criteria included current neurological conditions, bipolar disorder, current substance or alcohol dependence or primary psychotic disorder as assessed with the MINI [34]. Participants were also excluded on the basis of MRI contraindications (e.g., metal implants). Clinical assessments for trauma and PTSD were administered, and participants also completed an MRI scan. Clinical and demographic characteristics of this sample are described in Table 1. Monthly household income (self-reported) was dichotomized into two groups (<=$1000 vs > $1000/month) for statistical analyses, similar to prior studies[30]; this classification describes economic disadvantage in participants (low/minimal economic resources vs moderate or greater economic resources). Using this classification, over half the sample (61%) demonstrated significant economic disadvantage. The Institutional Review Board of Emory University and Grady Hospital Research Oversight Committee approved all study procedures.
Clinical assessments
Lifetime (childhood and adult) trauma assessments (Childhood Trauma Questionnaire, Traumatic Events Inventory), as well as PTSD assessments (PTSD Symptom Scale) were administered to all participants, detailed further in the Supplement. These variables used as covariates in analyses. The Experiences of Discrimination (EOD) questionnaire was used to assess experiences of RD, and a summed total of types of RD experienced was entered into statistical analyses.
MRI acquisition, image processing and statistical analyses
Scanning was conducted on a research-dedicated Siemens 3-Tesla TIM-Trio scanner as detailed in the Supplement. Freesurfer’s automated parcellation pipeline (Freesurfer version 5.3) was used to obtain cortical thickness, surface area and cortical volume for the cingulate cortex (left and right; detailed in Supplement), and gray matter volume for the hippocampus and amygdala (left and right), which were entered into analyses [35,36,37]. Cingulate cortex regions of interest included the left and right rostral anterior cingulate cortex (rACC), caudal anterior cingulate cortex (cACC), and posterior cingulate cortex (PCC). The cuneus and postcentral gyrus were examined as control regions.
Statistical analyses
Partial correlation analyses were conducted to examine the unique contributions of RD on cortical thickness, cortical volume and surface area for cingulate subregions, as well as contributions of RD on hippocampal and amygdala volume, after accounting for variance associated with other potentially contributing factors, including total intracranial volume (ICV), current PTSD symptoms, childhood and adult trauma exposure, and monthly income, as with our prior studies [16, 30]. Benjamini–Hochberg procedures were conducted to adjust for error due to multiple comparisons for each independent analysis; thickness, volume, and surface area for the cingulate, and volume of the amygdala and hippocampus, were each treated as independent tests. To assess specificity of significant findings, we repeated analyses with two other cortical regions, the postcentral gyrus and cuneus (left and right hemispheres); statistical significance was set at p < 0.05 for these analyses.
Age has an established relationship with gray matter metrics, including ICV [38, 39], and similarly demonstrated a strong relationship between RD and age in this sample. As such, we decided to exclude age from initial analyses to limit multicollinearity. However, regions that emerged as being significantly associated with RD were subject to follow-up sensitivity analyses with age group included as a covariate [younger (age 19–50) vs older (age 51–62)] as in our prior studies [40, 41] to examine whether findings persisted after accounting for age along with other covariates.
Results
Cingulate Cortex thickness
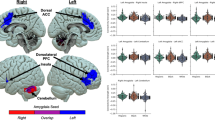
Partial correlation results are presented in Table 2a. After accounting for current PTSD, income level, trauma exposure (childhood and adulthood) and ICV, RD experiences were negatively correlated with thickness of the left rACC (r = −0.30, p = 0.007), left cACC (r = −0.28, p = 0.014), and left PCC (r = −0.30, p = 0.007); see Fig. 1; these results remained statistically significant after Benjamini–Hochberg correction for multiple comparisons. A re-analysis with age added as a covariate to the model revealed that only left rACC thickness emerged as associated with RD experiences [left rACC (r = −0.24, p = 0.032); cACC (r = −0.20, p = 0.082); left PCC (r = −0.18, p = 0.116)] but this association did not survive Benjamini–Hochberg correction.

A mask of the rostral cingulate cortex is shown in red; mask of the caudal cingulate cortex is shown in green, and mask of the posterior cingulate cortex is shown in yellow. a Partial correlation plot of residuals for racial discrimination and left posterior cingulate cortex thickness (r = −0.30, p = 0.007). b Partial correlation plot of residuals for racial discrimination and left caudal cingulate cortex thickness (r = −0.28, p = 0.014). c Partial correlation plot of residuals for racial discrimination and left rostral cingulate cortex thickness (r = −0.30, p = 0.007).
RD experiences were not significantly associated with thickness of the right rACC (r = −0.18; p = 0.117), right cACC (r = −0.08, p = 0.493), or right posterior cingulate cortex (r = −0.13, p = 0.271). As expected, RD was not significantly correlated with cortical thickness in our specified control regions including left cuneus (r = −0.05, p = 0.68), right cuneus (r = −0.05, p = 0.69), left postcentral gyrus (r = −0.19, p = 0.11) and right postcentral gyrus (r = −0.17, p = 0.15).
Cingulate cortex surface area
After accounting for current PTSD, income level, trauma exposure (childhood and adulthood) and ICV, RD experiences were not significantly correlated with surface area of any cingulate region; see Table 2b.
Cingulate cortex volume
After accounting for current PTSD, income level, trauma exposure (childhood and adulthood) and ICV, there were no relationships between RD and cingulate volume that surpassed our statistical threshold. RD demonstrated a nominally significant negative association with left posterior cingulate volume (r = −0.25, p = 0.03); see Table 2c.
Amygdala and hippocampal volume
Partial correlation results are presented in Table 3. After accounting for current PTSD, income level, trauma exposure (childhood and adulthood) and ICV, no associations between RD experiences and gray matter subcortical (amygdala, hippocampus) volume survived Benjamini–Hochberg correction for multiple comparisons. A nominally significant positive association of RD was observed with right hippocampus volume (r = 0.25, p = 0.03).
Discussion
In this study we examined associations between RD and thickness, surface area and volume of the cingulate cortex, as well as volume of the hippocampus and amygdala. We observed that, even after accounting for PTSD symptoms, socio-environmental factors such as economic disadvantage and other psychological trauma exposure, as well as ICV, more frequent RD was associated with proportionally less thickness throughout the left cingulate, including rostral, caudal and posterior regions. Repeat analyses with other brain regions (cuneus, postcentral gyrus) indicated that findings were specific to the cingulate. No associations were observed with cingulate surface area and volume. Given our own and others’ data indicating particularly potent associations of RD with functional and structural variations in cingulate brain regions and pathways [16, 42, 43], these findings shed light on the possible ways in which RD may have deleterious effects on the brain, paving the way for brain health disparities in Black Americans.
Racial discrimination is a type of racial trauma that is often encountered as a chronic, ongoing psychosocial stressor; this has been noted in the general U.S. population as well as individuals within the GTP population (e.g., [44,45,46,47]). Other types of chronic stressors, such as childhood maltreatment, potentiate adverse effects on prefrontal gray and white matter [48, 49], particularly the cingulate cortex [50]. Emerging data from longitudinal research suggests that, even after accounting for the effects of prior psychological trauma and other socio-environmental factors, RD increases vulnerability for later PTSD severity [4]. Our neurobiological findings extend this prior research, revealing a pathway through which this vulnerability can emerge.
The direction of our findings align with prior studies of PTSD and cingulate cortex volume [28, 51, 52] showing some possible convergence in pathophysiology. Cingulate findings were also localized to the left hemisphere, similar to prior PTSD meta and mega-analyses [51,52,53]. However, even after accounting for PTSD symptoms as well as socio-environmental factors such as economic disadvantage and other psychological trauma exposure, more frequent RD continued to be linked to proportionally less thickness throughout the left cingulate, including rostral, caudal and posterior regions. Repeat analyses with other brain regions (cuneus, postcentral gyrus) indicated that findings were specific to the cingulate. Although the reason for the laterality of our findings is unclear, localization of results to the left hemisphere are consistent with some of our prior RD white matter research, in which RD demonstrated particularly strong associations with the white matter integrity of the left anterior cingulum bundle [40]. Data from multiple longitudinal lifespan cohort studies has pointed to accelerated left-sided cingulate cortical atrophy in Alzheimer’s disease [54, 55] and early stage Parkinson’s disease [56]. Taken together, this raises the possibility that the present findings are markers of vulnerability to brain health problems, although longitudinal data is needed to confirm this.
Across morphological features, RD was specifically linked to cortical thickness, although a non-significant trend was also observed with volume, which is a function of cortical thickness and surface area. The latter two features, though related, are phenotypically different and appear to encompass distinct neural properties; while cortical thickness reflects neuronal/synaptic density and intracranial myelination, cortical surface area reflects folding and gyrification [57,58,59,60]. Given that the two indices appear to be related to different neurobiological processes, socio-environmental stressors such as RD may have dissociable effects on each. As the present findings were specific to cortical thickness, this gray matter metric may be a more sensitive biomarker of RD-related brain health vulnerabilities. However, this needs to be confirmed in long-term research that includes data on relevant cognitive and behavioral functions to determine if RD differentially affects these gray matter features over time.
It is clinically meaningful that these findings are consistent with earlier functional studies of RD that implicate frontal cortical regions, specifically the rostral and ventromedial prefrontal cortex [16, 17]. These anterior aspects of the cingulate gyrus are a hub for self-regulation, including regulation of cognitive and emotional processes such as response inhibition and emotion suppression [61]. Racist experiences can trigger a number of coping responses, including rumination about the event, emotion suppression and vigilance for similar threats[62]; these responses may encumber other cognitive and affective processes. In the context of other stressors (including other trauma) and demands, RD may unduly burden affective and cognitive control networks. By chronically hampering regulatory networks, RD reduces available resources for processes such as selective inhibition of distracting information, efficient attentional shifting, and acquisition and recall of new information. It can also reduce available resources for attending to and savoring pleasurable stimuli, including social interactions. These processes are essential to affect regulation, efficient completion of daily tasks and effective interpersonal communication. In conjunction with prior findings, these results point to a possible brain mechanism through which RD leads to cognitive and affective dysregulation. In addition, studies of social pain (including those with a rejection/exclusion experimental condition) converge to indicate the dorsal ACC as a node of increased activation under conditions of exclusion [63]. Some theorists suggest that social exclusion is frequently relived, preferentially activating these regions and prolonging the impact of the stressor. As such, it is possible that our structural findings indicate the impact of re-living of racially discriminatory experiences on stress-sensitive cingulate regions; however, a more granular assessment of rumination/distress related to RD is needed to test this hypothesis.
Racial discrimination’s contributions to allostatic load [9] can lead to “weathering” of cognitive control neural networks [64, 65], which increases risk for brain health problems; thinning of the cingulate cortex predicts attention and memory disruptions, setting the stage for later brain health problems [66]. Prior studies of trauma-exposed Black Americans show increased engagement of frontal cognitive control network regions during exposure to social threat [16, 17, 67]. We recently observed that RD corresponded with increased activation in the rostral ACC during attention to threat cues; notably, this pattern of response did not correspond with impairments in task performance [16]. Separately, we previously found that more frequent discrimination corresponded with lower fractional anisotropy (FA; an index of white matter integrity) in the anterior cingulum bundle, which extends through the length of the cingulate cortex [42], and FA in this region mediated the relationship between RD and number of medical disorders in Black American women [41]. Neuronal loss and cortical thinning in the cingulate cortex, particularly the posterior cingulate, has been consistently linked to Alzheimer’s disease [68, 69], conversely, conserved cingulate thickness is linked to preserved cognitive functioning and fewer histological markers of risk for Alzheimer’s Disease in older-aged individuals [70]. Given that Black Americans have a significantly elevated risk for Alzheimer’s disease than White Americans [71], our study findings may help to explain this health disparity. Our results converge with neurobiological studies of structural racism, which also point to altered function and structure of the cingulate cortex [72, 73]. In the context of these adversities, RD provides a further contribution to the cumulative burden of race-related stress. Together, these stressors may lead to neural remodeling of the cingulate cortex, potentially inducing effects on neural architecture that increase vulnerability for brain health morbidities.
There were no associations of RD with amygdala and hippocampal morphology that survived our statistical threshold, although a nominally significant positive association of RD was observed with volume of the right hippocampus. These findings are opposite of prior studies of chronic stress, PTSD and stigma which indicate hippocampal volume reductions [74, 75]. It is possible that our findings reflect adaptive neural mechanisms to RD, including increased sensitivity to social context, similar to a prior study of depression [76]. However, given that we did not assess relationships of RD and brain morphology with this psychological construct, this interpretation is merely speculative. Corresponding task data is needed to help elucidate these relationships.
We acknowledge limitations to this study, with a primary limitation being its cross-sectional design and the inherent difficulties with causal inference. Certainly, long-term longitudinal research can help to clarify temporal links between RD and gray matter alterations; it is possible that racist experiences could serve to increase gray matter in cingulate regions at earlier stages of development [in accordance with the stress acceleration hypothesis [77], and the nature of this relationship may reverse at later developmental stages. Nonetheless, given that other types of stressors are often concurrent with racially discriminatory experiences, distinguishing effects of these different stressors with high precision is not always possible. We are likewise unable to comment on mechanistic precursors for these findings. Further inquiry on potential mechanisms linked to individual (and systemic) racism are warranted in large-scale studies. Finally, intersectional discrimination, including gendered racism, was not assessed in this study, which is particularly relevant given that all participants were Black women. These intersectionalities merit exploration in further studies with a diverse range of gender representation. Finally, results were weakened by the addition of age as a covariate; this is unsurprising, as age has a potent association with gray matter metrics and with RD itself. The fact that some marginally significant associations remained after covarying age as well as other relevant variables (other trauma exposure) highlights the salience of RD, and implicates this stressor as a unique social determinant of brain health.
Altogether, the findings from this study complement our own and other data demonstrating a focal impact of RD on cingulate regions. It is clear that, like other types of trauma, RD can contribute to allostatic load and related deleterious effects on gray matter architecture. Distinct lines of inquiry around RD converge to suggest a specific effect on cingulate systems, possibly due to chronically heightened regulatory function. These decrements in cingulate gray matter are likely contributors to brain health disparities in Black individuals. In the context of earlier findings, it is clear that RD may contribute to the neural remodeling of cingulate pathways. Our findings indicate a compelling pathway through which brain health vulnerabilities may emerge in Black individuals related to the pernicious effects of racial trauma.
References
Kessler RC, Mickelson KD, Williams DR. The prevalence, distribution, and mental health correlates of perceived discrimination in the United States. J Health Soc Behav. 1999;40:208–30.
Lee RT, Perez AD, Boykin CM, Mendoza-Denton R. On the prevalence of racial discrimination in the United States. PloS One. 2019;14:e0210698.
Paradies Y, Ben J, Denson N, Elias A, Priest N, Pieterse A, et al. Racism as a determinant of health: a systematic review and meta-analysis. PloS One. 2015;10:e0138511.
Bird CM, Webb EK, Schramm AT, Torres L, Larson C, deRoon-Cassini TA. Racial discrimination is associated with acute posttraumatic stress symptoms and predicts future posttraumatic stress disorder symptom severity in trauma-exposed black adults in the United States. J Trauma Stress. 2021;34:995–1004. https://doi.org/10.1002/jts.22670.
Everson-Rose SA, Lutsey PL, Roetker NS, Lewis TT, Kershaw KN, Alonso A, et al. Perceived discrimination and incident cardiovascular events: the multi-ethnic study of atherosclerosis. Am J Epidemiol. 2015;182:225–34.
Zahodne LB, Kraal AZ, Sharifian N, Zaheed AB, Sol K. Inflammatory mechanisms underlying the effects of everyday discrimination on age-related memory decline. Brain, Behav, Immun. 2019;75:149–54.
Zahodne LB, Morris EP, Sharifian N, Zaheed AB, Kraal AZ, Sol K. Everyday discrimination and subsequent cognitive abilities across five domains. Neuropsychology. 2020;34:783–90. https://doi.org/10.1037/neu0000693.
Brosschot JF. Markers of chronic stress: prolonged physiological activation and (un)conscious perseverative cognition. Neurosci Biobehav Rev. 2010;35:46–50.
Ong AD, Williams DR, Nwizu U, Gruenewald TL. Everyday unfair treatment and multisystem biological dysregulation in African American adults. Cult Divers Ethn Minor Psychol. 2017;23:27–35.
Cuevas AG, Ong AD, Carvalho K, Ho T, Chan SWC, Allen JD, et al. Discrimination and systemic inflammation: A critical review and synthesis. Brain, Behav, Immun. 2020;89:465–79.
Kirkinis K, Pieterse AL, Martin C, Agiliga A, Brownell A. Racism, racial discrimination, and trauma: a systematic review of the social science literature. Ethn Health. 2021;26:392–412.
Causadias JM, Korous KM. Racial discrimination in the United States: a national health crisis that demands a national health solution. J Adolesc Health. 2019;64:147–8.
Williams DR, Lawrence JA, Davis BA. Racism and health: evidence and needed research. Annu Rev Public Health. 2019;40:105–25.
Hill LK, Hu DD, Koenig J, Sollers JJ 3rd, Kapuku G, Wang X, et al. Ethnic differences in resting heart rate variability: a systematic review and meta-analysis. Psychosom Med. 2015;77:16–25.
Clark US, Miller ER, Hegde RR. Experiences of discrimination are associated with greater resting amygdala activity and functional connectivity. Biol Psychiatry Cogn Neurosci Neuroimaging. 2018;3:367–78.
Fani N, Carter SE, Harnett NG, Ressler KJ, Bradley B. Association of racial discrimination with neural response to threat in black women in the US exposed to trauma. JAMA Psychiatry. 2021;78:1005–12.
Masten CL, Telzer EH, Eisenberger NI. An fMRI investigation of attributing negative social treatment to racial discrimination. J Cogn Neurosci. 2011;23:1042–51.
McEwen BS, Nasca C, Gray JD. Stress effects on neuronal structure: hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacol: Off Publ Am Coll Neuropsychopharmacol. 2016;41:3–23.
Stevens FL, Hurley RA, Taber KH. Anterior cingulate cortex: unique role in cognition and emotion. J Neuropsychiatry Clin Neurosci. 2011;23:121–5.
Leech R, Sharp DJ. The role of the posterior cingulate cortex in cognition and disease. Brain J Neurol. 2014;137:12–32.
Leech R, Smallwood J. The posterior cingulate cortex: Insights from structure and function. Handb Clin Neurol. 2019;166:73–85.
Nejad AB, Fossati P, Lemogne C. Self-referential processing, rumination, and cortical midline structures in major depression. Front Hum Neurosci. 2013;7:666.
Harrell CJ, Burford TI, Cage BN, Nelson TM, Shearon S, Thompson A, et al. Multiple pathways linking racism to health outcomes. Du Bois Rev. 2011;8:143–57.
Nagata JM, Ganson KT, Sajjad OM, Benabou SE, Bibbins-Domingo K. Prevalence of perceived racism and discrimination among US children aged 10 and 11 years: the Adolescent Brain Cognitive Development (ABCD) study. JAMA Pediatr. 2021;175:861–3.
Hinojosa CA, Kaur N, VanElzakker MB, Shin LM. Cingulate subregions in posttraumatic stress disorder, chronic stress, and treatment. Handb Clin Neurol. 2019;166:355–70.
Tottenham N, Sheridan MA. A review of adversity, the amygdala and the hippocampus: a consideration of developmental timing. Front Hum Neurosci. 2009;3:68.
Yamasue H, Kasai K, Iwanami A, Ohtani T, Yamada H, Abe O, et al. Voxel-based analysis of MRI reveals anterior cingulate gray-matter volume reduction in posttraumatic stress disorder due to terrorism. Proc Natl Acad Sci USA 2003;100:9039–43.
Shin LM, Liberzon I. The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacol: Off Publ Am Coll Neuropsychopharmacol. 2010;35:169–91.
Logue MW, van Rooij SJH, Dennis EL, Davis SL, Hayes JP, Stevens JS, et al. Smaller hippocampal volume in posttraumatic stress disorder: a multisite ENIGMA-PGC study: subcortical volumetry results from posttraumatic stress disorder consortia. Biol psychiatry. 2018;83:244–53.
Fani N, Harnett NG, Bradley B, Mekawi Y, Powers A, Stevens JS, et al. Racial discrimination and white matter microstructure in trauma-exposed Black women. Biological Psychiatry. In press.
Zahodne LB, Sharifian N, Kraal AZ, Morris EP, Sol K, Zaheed AB, et al. Longitudinal associations between racial discrimination and hippocampal and white matter hyperintensity volumes among older Black adults. Soc Sci Med. 2022:114789. https://doi.org/10.1016/j.socscimed.2022.114789.
Meyer CS, Schreiner PJ, Lim K, Battapady H, Launer LJ. Depressive symptomatology, racial discrimination experience, and brain tissue volumes observed on magnetic resonance imaging. Am J Epidemiol. 2019;188:656–63.
Webb EK, Bird CM, deRoon-Cassini TA, Weis CN, Huggins AA, Fitzgerald JM, et al. Racial discrimination and resting-state functional connectivity of salience network nodes in trauma-exposed black adults in the United States. JAMA Netw Open. 2022;5:e2144759.
Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, et al. The Mini-International Neuropsychiatric Interview (M.I.N.I.): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry. 1998;59:22–33.
Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis. I. Segmentation and surface reconstruction. NeuroImage. 1999;9:179–94.
Fischl B, Sereno MI, Dale AM. Cortical surface-based analysis. II: Inflation, flattening, and a surface-based coordinate system. NeuroImage. 1999;9:195–207.
Fischl B, van der Kouwe A, Destrieux C, Halgren E, Segonne F, Salat DH, et al. Automatically parcellating the human cerebral cortex. Cereb Cortex. 2004;14:11–22.
Gunning-Dixon FM, Brickman AM, Cheng JC, Alexopoulos GS. Aging of cerebral white matter: a review of MRI findings. Int J Geriatr Psychiatry. 2009;24:109–17.
Lebel C, Treit S, Beaulieu C. A review of diffusion MRI of typical white matter development from early childhood to young adulthood. NMR Biomed. 2019;32:e3778.
Fani N, Harnett NG, Bradley B, Mekawi Y, Powers A, Stevens JS, et al. Racial discrimination and white matter microstructure in trauma-exposed black women. Biol Psychiatry. 2022;91:254–61.
Okeke O, Elbasheir A, Carter S, Powers A, Mekawi Y, Gillespie CF, et al. Indirect effects of racial discrimination on health outcomes through prefrontal cortical white matter integrity. Biol Psychiatry Cogn Neurosci Neuroimaging. 2022. https://doi.org/10.1016/j.bpsc.2022.05.004.
Fani N, Harnett NG, Bradley B, Mekawi Y, Powers A, Stevens JS, et al. Racial discrimination and white matter microstructure in trauma-exposed Black women. Biol Psychiatry. in press.
Isamah N, Faison W, Payne ME, MacFall J, Steffens DC, Beyer JL, et al. Variability in frontotemporal brain structure: the importance of recruitment of African Americans in neuroscience research. PloS One. 2010;5:e13642.
Mekawi Y, Carter S, Brown B, Martinez de Andino A, Fani N, Michopoulos V, et al. Interpersonal trauma and posttraumatic stress disorder among black women: does racial discrimination matter? J Trauma Dissociation: Off J Int Soc Study Dissociation. 2021;22:154–69.
Mekawi Y, Watson-Singleton NN, Kuzyk E, Dixon HD, Carter S, Bradley-Davino B, et al. Racial discrimination and posttraumatic stress: examining emotion dysregulation as a mediator in an African American community sample. Eur J Psychotraumatology. 2020;11:1824398.
Smith WA, Allen WR, Danley LL. “Assume the position… You fit the description” - Psychosocial experiences and racial battle fatigue among African American male college students. Am Behav Sci. 2007;51:551–78.
Comas-Diaz L, Hall GN, Neville HA. Racial trauma: theory, research, and healing: introduction to the special issue. Am Psychol. 2019;74:1–5.
Gehred MZ, Knodt AR, Ambler A, Bourassa KJ, Danese A, Elliott ML, et al. Long-term neural embedding of childhood adversity in a population-representative birth cohort followed for five decades. Biol Psychiatry. 2021;74:1–5. https://doi.org/10.1037/amp0000442.
Gur RE, Moore TM, Rosen AFG, Barzilay R, Roalf DR, Calkins ME, et al. Burden of environmental adversity associated with psychopathology, maturation, and brain behavior parameters in youths. JAMA Psychiatry. 2019;90:182–193. https://doi.org/10.1016/j.biopsych.2021.02.971.
Hart H, Rubia K. Neuroimaging of child abuse: a critical review. Front Hum Neurosci. 2012;6:52.
Meng L, Jiang J, Jin C, Liu J, Zhao Y, Wang W, et al. Trauma-specific grey matter alterations in PTSD. Sci Rep. 2016;6:33748.
Wang X, Xie H, Chen T, Cotton AS, Salminen LE, Logue MW, et al. Cortical volume abnormalities in posttraumatic stress disorder: an ENIGMA-psychiatric genomics consortium PTSD workgroup mega-analysis. Mol Psychiatry. 2021;26:4331–43.
Bromis K, Calem M, Reinders A, Williams SCR, Kempton MJ. Meta-analysis of 89 structural mri studies in posttraumatic stress disorder and comparison with major depressive disorder. Am J Psychiatry. 2018;175:989–98.
Thompson PM, Hayashi KM, de Zubicaray G, Janke AL, Rose SE, Semple J, et al. Dynamics of gray matter loss in Alzheimer’s disease. J Neurosci Off J Soc Neurosci. 2003;23:994–1005.
Roe JM, Vidal-Pineiro D, Sorensen O, Brandmaier AM, Duzel S, Gonzalez HA, et al. Asymmetric thinning of the cerebral cortex across the adult lifespan is accelerated in Alzheimer’s disease. Nat Commun. 2021;12:721.
Yau WY, Bischoff-Grethe A, Theilmann RJ, Torres L, Wagner A, Kaye WH, et al. Alterations in white matter microstructure in women recovered from anorexia nervosa. Int J Eat Disord. 2013;46:701–8.
Rakic P. Evolution of the neocortex: a perspective from developmental biology. Nat Rev Neurosci. 2009;10:724–35.
Schuz A, Palm G. Density of neurons and synapses in the cerebral cortex of the mouse. J Comp Neurol. 1989;286:442–55.
Panizzon MS, Fennema-Notestine C, Eyler LT, Jernigan TL, Prom-Wormley E, Neale M, et al. Distinct genetic influences on cortical surface area and cortical thickness. Cereb Cortex. 2009;19:2728–35.
Winkler AM, Kochunov P, Blangero J, Almasy L, Zilles K, Fox PT, et al. Cortical thickness or grey matter volume? The importance of selecting the phenotype for imaging genetics studies. NeuroImage. 2010;53:1135–46.
Posner MI, Rothbart MK, Sheese BE, Tang Y. The anterior cingulate gyrus and the mechanism of self-regulation. Cogn Affect Behav Neurosci. 2007;7:391–5.
Del Giudice M, Ellis BJ, Shirtcliff EA. The Adaptive Calibration Model of stress responsivity. Neurosci Biobehav Rev. 2011;35:1562–92.
Eisenberger NI. Social pain and the brain: controversies, questions, and where to go from here. Annu Rev Psychol. 2015;66:601–29.
Dufford AJ, Kim P. Family income, cumulative risk exposure, and white matter structure in middle childhood. Front Hum Neurosci. 2017;11:547.
Zsoldos E, Filippini N, Mahmood A, Mackay CE, Singh-Manoux A, Kivimaki M, et al. Allostatic load as a predictor of grey matter volume and white matter integrity in old age: The Whitehall II MRI study. Sci Rep. 2018;8:6411.
Peterson BS, Warner V, Bansal R, Zhu H, Hao X, Liu J, et al. Cortical thinning in persons at increased familial risk for major depression. Proc Natl Acad Sci USA 2009;106:6273–8.
Fani N, Jovanovic T, Ely TD, Bradley B, Gutman D, Tone EB, et al. Neural correlates of attention bias to threat in post-traumatic stress disorder. Biol Psychol. 2012;90:134–42.
Matsuda H. Role of neuroimaging in Alzheimer’s disease, with emphasis on brain perfusion SPECT. J Nucl Med. 2007;48:1289–300.
Scheff SW, Price DA, Ansari MA, Roberts KN, Schmitt FA, Ikonomovic MD, et al. Synaptic change in the posterior cingulate gyrus in the progression of Alzheimer’s disease. J Alzheimers Dis. 2015;43:1073–90.
Gefen T, Peterson M, Papastefan ST, Martersteck A, Whitney K, Rademaker A, et al. Morphometric and histologic substrates of cingulate integrity in elders with exceptional memory capacity. J Neurosci: Off J Soc Neurosci. 2015;35:1781–91.
Alzheimer’s Disease Facts and Figures Report 2020 [Available from: https://www.alz.org/media/Documents/alzheimers-facts-and-figures.pdf.
Webb EK, Weis CN, Huggins AA, Fitzgerald JM, Bennett K, Bird CM, et al. Neural impact of neighborhood socioeconomic disadvantage in traumatically injured adults. Neurobiol Stress. 2021;15:100385.
Harnett NG, Wheelock MD, Wood KH, Goodman AM, Mrug S, Elliott MN, et al. Negative life experiences contribute to racial differences in the neural response to threat. NeuroImage. 2019;202:116086.
Kim EJ, Pellman B, Kim JJ. Stress effects on the hippocampus: a critical review. Learn Mem. 2015;22:411–6.
Hatzenbuehler ML, Weissman DG, McKetta S, Lattanner MR, Ford JV, Barch DM, et al. Smaller hippocampal volume among black and latinx youth living in high-stigma contexts. J Am Acad Child Adolesc Psychiatry. 2022;61:809-819.
Schriber RA, Anbari Z, Robins RW, Conger RD, Hastings PD, Guyer AE. Hippocampal volume as an amplifier of the effect of social context on adolescent depression. Clin Psychol Sci. 2017;5:632–49.
Callaghan BL, Tottenham N. The stress acceleration hypothesis: effects of early-life adversity on emotion circuits and behavior. Curr Opin Behav Sci. 2016;7:76–81.
Acknowledgements
We wish to thank Allen Graham, Rebecca Hinrichs, Angelo Brown and other members of the Grady Trauma Project, as well as members of the Fani Affective Neuroscience Lab, for their assistance with data collection. We thank Timothy Ely for his technical assistance. We thank participants of the Grady Trauma Project for their time and involvement in this study.
Funding
This work was primarily supported by National Institute of Mental Health (MH101380 to NF, MH119603 to NGH, MH-071537 and MH094757 to KJR, HD071982 to BB) and National Center for Complementary and Integrative Health (AT011267 to NF). Support was also received from the Emory Medical Care Foundation, Emory University Research Council, American Psychological Association, Society for Clinical Neuropsychology and the Frazier Foundation Grant for Mood and Anxiety Research at McLean Hospital, and the National Cancer Institute (CA220254-02S1).
Author information
Authors and Affiliations
Contributions
The authors confirm contribution to the paper as follows: study conception and design: NF, SC, KR, BB; data collection: NF, JS; analysis and interpretation of results: LE, NF, NGH, SEC, MP, SJHvR; draft manuscript preparation: LE, NF, NGH. All authors reviewed the results and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
KJR has received unrelated consulting income or sponsored research from Alkermes, Brainsway and Genomind, and is on scientific advisory boards for Janssen, Takeda, and Verily. The remaining authors have no financial conflicts of interest to disclose.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Fani, N., Eghbalzad, L., Harnett, N.G. et al. Racial discrimination associates with lower cingulate cortex thickness in trauma-exposed black women. Neuropsychopharmacol. 47, 2230–2237 (2022). https://doi.org/10.1038/s41386-022-01445-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41386-022-01445-8
