
Abstract
Background:
Attention problems are among the most prominent behavioral deficits reported in very preterm children (below 32 wk of gestation) at school age. In this study, we aimed to elucidate the brain abnormalities underlying attention problems in very preterm children by investigating the role of abnormalities in white and gray brain matter during interference control, using functional magnetic resonance imaging (fMRI)–guided probabilistic diffusion tensor tractography.
Methods:
Twenty-nine very preterm children (mean (SD) age: 8.6 (0.3) y), and 47 term controls (mean (SD) age: 8.7 (0.5) y), performed a fMRI version of the Eriksen Flanker task measuring interference control.
Results:
Very preterm children showed slower reaction times than term controls when interfering stimuli were presented, indicating poorer interference control. Very preterm children and term controls did not differ in mean activation of the cortical regions involved in interference control. However, impaired fractional anisotropy (FA) was found in very preterm children in specifically those fiber tracts that innervate the cortical regions involved in interference control. Lower FA was related to poorer interference control in very preterm children.
Conclusion:
White matter alterations have a crucial role in the interference control problems of very preterm children at school age.
Similar content being viewed by others

Main
Improved perinatal care has increased survival rates of very preterm (<32 wk of gestation) infants. However, large alterations in brain development remain present throughout childhood and adolescence (1), and surviving very preterm children appear to have pervasive behavioral problems more frequently than term peers (2,3). At school age, attention problems are among the most prominent behavioral deficits in very preterm children (4,5), having a detrimental impact on school performance and social functioning.
One core aspect of attention is interference control, the ability to suppress distracters that might slow the primary response (6). The brain attention network involved in interference control has been widely investigated using functional magnetic resonance imaging (fMRI) tasks including the Eriksen Flanker task (7,8,9), suggesting involvement of the dorsal anterior cingulate cortex (dACC), inferior and lateral prefrontal cortices, and inferior parietal cortices.
This study aims to elucidate brain abnormalities underpinning attention problems in very preterm children, by investigating the role of impairments in white and gray brain matter in interference control in very preterm children and term controls. First, we examined interference control problems and potential differences in cortical activation between very preterm children and term controls using the Eriksen Flanker task adapted for fMRI. Second, by using fMRI-guided probabilistic diffusion tensor tractography, we studied potential differences between very preterm children and term controls in white matter development using values of fractional anisotropy (FA) of specifically those fiber tracts that innervate cortical regions activated in interference control. Finally, the functional relations between cortical activation, FA values, and interference control were explored in very preterm children and term controls.
Results
Demographic Variables
Sample characteristics are shown in Table 1 . Age at MRI scan (P = 0.28), social economic status (P = 0.42), and gender (P = 0.75) were not different between the 29 very preterm children and the 47 term controls. Very preterm children had significantly lower full-scale intelligence quotient (IQ) scores than term peers as measured using the Wechsler Intelligence Scales for Children III (WISC-III) (d = 0.81; P < 0.001). Full-scale IQ was related to reaction time (RT) performance during each condition (range r = −0.23 to −0.25; all P values <0.05) but not to the interference effect (r = 0.17; P = 0.12).
Eriksen Flanker Task
There was a significant effect of group (F(1,74) = 6.15; P = 0.02) and condition (F(2,73) = 73.40; P < 0.001), on Flanker task RT performance ( Table 1 ). In post hoc analyses, RT was significantly slower in the incongruent trials compared with that in the neutral trials (d = 0.70; P < 0.001) and congruent trials (d = 0.79; P < 0.001), whereas RT was not different between the neutral trials and the congruent trials (d = 0.09; P = 0.61). In addition, there was a significant larger interference effect in very preterm children (d = 0.67; P = 0.005), substantiating the presence of poorer interference control in very preterm children as compared with term controls. A significant larger interference effect in very preterm children remained present when covarying for the differences in full-scale IQ scores (P = 0.049). No significant differences between very preterm children and term controls were found in error percentage for each of the three conditions. In addition, there were no differences in task performance between children from which data of two blocks were included and children from which data of one block were removed due to too much head motion.
Functional Magnetic Resonance Imaging
We did not find any evidence for differences in activated areas between very preterm children and term controls during incongruent trials as compared with neutral and congruent trials. The whole-brain analysis of cluster activation on the whole sample of very preterm children and term controls showed three regions of interest (ROIs) with significantly increased activation during incongruent trials as compared with neutral and congruent trials in the left parietal cortex, right parietal cortex, and dACC ( Table 2 ). Furthermore, during the incongruent condition of the Flanker task, the increase in activation of the left parietal cortex (P = 0.14), right parietal cortex (P = 0.70), and the dACC (P = 0.50) were not different between very preterm children and term controls. To ensure that there were no group differences in the mean activation of the cortical area directly surrounding the three ROIs, we additionally tested differences in mean activation of the three ROIs as determined using less stringent P value (false-discovery rate). This approach also did not show any group differences in increase in activation of the left parietal cortex (P = 0.08), right parietal cortex (P = 0.47), and the dACC (P = 0.21). There were no differences in activation between children from which data of two blocks were included and children from which data of one block were removed due to too much head motion.
Diffusion Tensor Imaging
The three ROIs showing increased activation during the incongruent condition (left parietal cortex, right parietal cortex, and dACC) were used as seeds in tracking analyses. In order to determine the fiber tract connections between the left and right parietal regions and the ACC, we subdivided the tract in parts, with the thalamus as “relay station.” First, connections between the left parietal cortex and the left thalamus, and between the right parietal cortex and the right thalamus, were tracked. Second, connections between the dACC and the bilateral thalami were tracked. In this latter analysis, the left and right thalami were combined, as many fibers from both the left and the right thalamus cross the corpus callosum to the contralateral part of the dACC, creating the risk that (parts of) tracts are erroneously included twice. Data of three children (one very preterm and two term controls) were excluded, as the implemented tracking algorithm was not successful in finding any connections between the left and right thalamus and the dACC. For these three children, more than 10% of the volumes had to be removed due to the presence of artifacts, leading to lower diffusion tensor imaging (DTI) data quality and consequently the inability to track any connections.
The volumes of the three tracts were not different between very preterm children and term controls ( Figure 1 ). Mean FA value was significantly higher in very preterm children as compared with term controls in the fiber tract connecting the right parietal cortex and the right thalamus (d = 0.59; P = 0.02), underpinned by higher axial diffusivity (L1) values in very preterm children (d = 0.66; P = 0.008). In addition, mean FA value was significantly lower in the fiber tracts connecting the left and right thalamus with the dACC (d = 0.61; P = 0.01), which was underpinned by higher radial diffusivity (L23) values in very preterm children as compared with term controls (d = 0.66; P = 0.007).

Diffusion tensor imaging (DTI) results: fractional anisotropy (FA) values, axial diffusivity (L1), radial diffusivity (L23), and tract volume in voxels in very preterm children and term controls. Values of axial diffusivity and radial diffusivity are depicted in 10–5 mm2/s. ACC, anterior cingulate cortex.
We did not find any evidence for an association between the functional activation in the selected ROIs and the average FA value of the fiber tracts that innervated the activated cortical regions (range r = −0.11 to 0.02; all P values >0.34).
Functional Relations
A higher interference effect in the Eriksen Flanker task was significantly associated with increased dACC activation in the whole group (r = 0.34; P = 0.002). Using a Fisher r-to-z transformation test, we found no evidence for a difference between very preterm children and term controls in terms of the association between the dACC activation and the interference effect (z = 0.51; P = 0.31). Furthermore, a higher interference effect was also associated with lower FA values of the tract connecting the left and right thalamus with the dACC in the whole group (r = -0.26; P = 0.03). Interestingly, this association was only significant in very preterm children (r = −0.46; P = 0.02) and nonexistent for term controls (r = 0.03; P = 0.83). Using a Fisher r-to-z transformation test, this difference in the association between lower FA values of the tract connecting the left and right thalamus with the dACC and a higher interference effect between very preterm children and term controls was significant (z = 2.09; P = 0.04).
Discussion
This study investigated the nature of attention deficits in very preterm children by studying the role of white and gray brain matter abnormalities in interference control, as measured using the Eriksen Flanker task. We found significantly larger behavioral interference in very preterm children compared with term controls (d = 0.67; P = 0.005), indicating that very preterm children have more difficulty with interference control than term peers. By using fMRI-guided probabilistic diffusion tensor tractography, we demonstrated differences between very preterm children and term controls in FA values of specifically those fiber tracts that innervate the activated cortical regions involved in interference control.
Our findings confirm the increase in mean activation of the dACC as well as the left and right parietal cortex during interference control (7,8). A previous study has shown that the left and right parietal cortex are involved in “activating” all possible responses, whereas the dACC is involved in the selection of the appropriate response during interference control (7). Interestingly, an increase in mean activation of the dACC was related to poorer interference control, suggesting a potential compensatory increase in cortical activation during poor interference control. Together, the poorer interference control performance of very preterm children as compared with term controls, and a potential compensatory increase in cortical activation, may suggest the presence of differences in the cortical activation between both groups. Indeed, some other studies have described differences in cortical activation patterns between very preterm children and term controls, investigating the brain systems underlying language functions, motor response inhibition, and visual memory (10,11,12,13). However, our results did not show differences in the mean activation of cortical regions underlying interference control between very preterm children and term controls at school age. This finding may demonstrate that utilization of additional cortical activation to compensate for poor interference control is only possible to a limited extent and hence does not translate into significant group differences in cortical activation. However, this interpretation warrants confirmation in other studies.
Widespread differences in white matter development have been frequently described in very preterm children throughout childhood and adolescence (14,15,16,17). In general, white matter development comprises a cascade of events including the development of preoligodendrocytes, axons, and subplate neurons, which may be negatively affected in preterm children by multiple factors with accompanying excitotoxicity and ischemic events (18). In this study, we found that altered FA values of fiber tracts connecting the thalamus with the dACC underpin interference control problems of very preterm children. Interestingly, this finding may illustrate a pathway by which the thalamus, as a cortical “relay station,” is associated with cognitive and executive dysfunctioning of very preterm children (19,20). Measures of radial and axial diffusivity may elucidate the origin of differences in FA values between very preterm children and term controls, although interpretation(s) in terms of the underlying pathology might need further validation (21). Our findings suggest that a lower FA value of the tract connecting the left and right thalami with the dACC in very preterm children was characterized by an increase in radial diffusivity. The increase in FA value of the tract connecting the right parietal cortex with the right thalamus was characterized by an increase in axial diffusivity, suggesting a potential compensatory increase in axonal density and axonal caliber of this tract to counterbalance the negative effects of a reduced FA value in the tract connecting the right and left thalami with the dACC in very preterm children. The presence of this compensational increase in particularly the right hemisphere may potentially depend on the spatial properties of the assessed interference control task, given that the right parietal hemisphere is more involved in visuospatial processing than the left parietal hemisphere. Adaptive compensation, in terms of an increase in FA values, has been described in children with cerebral paresis in the motor tracts of the unaffected hemisphere (22). This potential adaptive compensation may be the result of a complex process of activity-dependent “pruning” that normally takes place during childhood (22); however, other factors may also explain the observed increase in FA value in very preterm children. For instance, given the impact of very preterm birth on white matter development, it is not unlikely that less fibers crossed the white matter tracts in very preterm children as compared with term controls, which may have resulted in the relatively higher FA value for very preterm children. Interestingly, other studies also described increases in FA values of similar tracts in very preterm children at term, in childhood, or at adolescent age (23,24,25). Future studies are warranted to further clarify the potential presence of adaptive compensation in white matter tracts underlying interference control in very preterm children.
This study has some limitations that need to be taken into account. First, some studies showed differences in FA values between males and females (14) or illustrated an effect of gender regarding relations between FA values and functional outcomes (26). However, we did not find any evidence for differences between males and females on our measures of cortical activation, FA values, or functional outcomes. Second, in order to maximize reliability of tractography analyses, a conservative approach for defining ROIs was used, potentially underpinning differences with other studies describing an additional involvement of the inferior and/or lateral prefrontal cortices in the selection between competing responses (7). In addition, differences in prefrontal regions involvement across studies using the Eriksen Flanker task may also depend on the type of stimuli (letters or arrows) used to evoke the interference effect (7). Although crucially involved in attentional performance, interference control is only one of multiple aspects underlying attention in children. Other aspects, including the abilities to orient to, to maintain focus, or to shift attention between events (9), may also play an important role in attention deficits of very preterm children (5), indicating that future studies should further explore the underlying pathology of attention deficits in very preterm children. Finally, our sample had a restricted age range. However, some studies indicate that the neural network underpinning interference control alters with age (27), implying that the differences between very preterm children and term controls as described in this study may change over time. Nevertheless, current differences were present 8 y after the neonatal period, suggesting that the effects of very preterm birth on the neural network underpinning interference control are rather persistent.
In summary, by using fMRI-guided diffusion tensor tractography, this study presents evidence that differences in FA values underpin interference control problems of very preterm children. The important role of abnormal white matter development in interference control problems of very preterm children suggests a window of opportunity for future interventions directed at the improvement of the attention problems of these children.
Methods
Sample
All very preterm children (<32 wk of gestation) admitted to the level III neonatal intensive care unit of the VU University Medical Center Amsterdam between September 2001 and July 2003 were eligible for inclusion in a randomized controlled trial on the effects of glutamine supplementation (28). As we found some evidence that glutamine supplementation may have influenced brain development, only very preterm children of the control group of this trial were included in this study. At 7–8 y of age, (parents of) 30 of the 39 children (77%) of the control group agreed to participate in the follow-up study, and 29 (97%) successfully finished MRI follow-up at the mean (SD) age of 8.6 (0.3) y. MRI data for one child were not collected due to restrictions in scanning time availability.
Age-matched, term born peers (>37 wk of gestation) from the same classrooms as attended by the very preterm children, or recruited from other schools located in the same area, were invited to participate in the study. Controls had to be born without any perinatal complications as reported by their parents, to attend regular classes, and to be free of behavioral and emotional problems and academic difficulties as reported by their teacher. A total of 51 term born peers agreed to participate in the scanning session. Data for one child were not collected due to restrictions in scanning time availability, and data of three children had to be excluded due to too much head movement (>3 mm) during the fMRI scanning blocks. A total of 47 term born peers successfully finished MRI at the mean (SD) age of 8.7 (0.5) y.
Cognitive functioning was estimated using full-scale IQ score, as measured by a short form of WISC-III (29), including the subtests Vocabulary and Block Design. This short form composite score has satisfactory reliability (r = 0.91) and correlates highly (r = 0.86) with full-scale IQ.
Procedure
All parents completed written informed consent prior to the study, which was approved by the medical ethical committee of the VU University Medical Center. MRI took place at the VU Medical Center. A simulation scanner was used for subjects to get comfortable with the scanner environment and procedures (30).
Acquisition
MRI was performed with a 1.5 T MRI scanner, equipped with an 8-channel phased-array head coil (Siemens Sonata, Erlangen, Germany). Structural three-dimensional (3D) T1-weighted images were obtained in the sagittal plane with an MP-RAGE (magnetization-prepared rapid acquisition gradient echo) sequence (repetition time = 2,730 ms, echo time = 3.7 ms, inversion time = 1,000 ms, flip angle = 7°, with a 1 × 1 mm in-plane resolution and slice thickness of 1 mm). Functional MRI images were acquired using an echo planar imaging sequence (repetition time = 2,400 ms, echo time = 40 ms, flip angle = 90°, 36 slices, slice gap = 17%, with a 3.1 × 3.1 mm in-plane resolution and slice thickness of 3.1 mm). DTI images were collected during one acquisition with single-shot echo planar imaging consisting of four volumes without directional weighting, and 24 volumes with 24 noncollinear gradient directions (b value = 750 s/mm2, repetition time = 7,500 ms, echo time = 85 ms, with a 2.5 × 2.5 mm in-plane resolution and 54 contiguous slices with slice thickness of 2.5 mm).
Eriksen Flanker Task
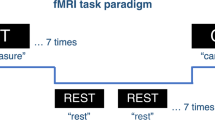
During fMRI scanning, performance on an adapted version of the Eriksen Flanker task (31) was measured. In contrast to other inhibition related tasks, stimuli are presented spatially adjacent to the target in the Eriksen Flanker task, which may better reflect interference as experienced in everyday activities (6). In this task, children had to respond as fast as possible to a centrally presented arrow pointing toward the left or the right, by pressing a button with their left or right index finger, respectively. Two flanker arrows were presented on both sides of the central arrow, which were congruent (four arrows pointing in the same direction as the central arrow), incongruent (four arrows pointing in the opposite direction as the central arrow), or neutral (four dashes) with the central arrow, yielding three different conditions. Incongruent flanker arrows interfere with the response to the central arrow and therefore need to be inhibited. This inhibition leads to an interference effect (31), a well-documented increase in RT, providing a measure of the interference control. The interference effect was calculated by subtracting the average RT in the incongruent condition from the average RT in the congruent and neutral condition. A total of 90 trials were administered, divided over two scanning blocks of 45 trials each. There were 30 trials for each condition, presented in a fixed random order, completely balanced across left and right pointing central arrows. Stimuli were presented for 3,000 ms but disappeared visually when a button was pressed. An additional interstimulus interval, varying from 1,500–2,500 ms, was present in a fixed random order between two trials.
fMRI Data Processing
Processing of fMRI data was performed using BrainVoyager QX software (Brain Innovation, Maastricht, The Netherlands). Series of 113 volumes were acquired for each of the two scanning blocks of 45 trials of the Eriksen Flanker task. The first two volumes were discarded in order to avoid differences in T1 saturation. Voxel time series of the remaining 111 volumes were slice time– and motion-corrected, high-pass filtered (0.02 Hz), temporally (3.0-s full width at half maximum Gaussian kernel), and 3D spatially (6-mm full width at half maximum Gaussian kernel) smoothed. A block of trials was excluded from further analyses when head motion during the block exceeded 3 mm in the x, y, or z direction. As indicated, this was the case for three term controls for both blocks of trials. For 19 children (9 very preterm children and 10 term controls), data of one block of trials were removed due to too much head motion. For each child, the individual 3D T1 scan was registered to standard Talairach space. The individual echo planar imaging volumes were first coregistered with the individual 3D T1 scan, followed by the corresponding transformation into Talairach space. Voxel time series were corrected for serial correlations. First, whole-brain activation obtained in response to the incongruent condition as opposed to all other trials (congruent and neutral trials) was compared between very preterm children and term controls using a P value set at false-discovery rate. Next to the regressor for the interference effect, we included a separate regressor for errors, as the activation during these trials should not be confounded with the activation involved in suppressing the interference effect. Both regressors were convolved with a standard hemodynamic response function. Second, ROIs were defined based on whole-brain activation obtained for the whole sample of very preterm children and term controls. To ensure involvement of cortical regions in interference control, and to make sure that regions were not erroneously being used as seed in tractography analyses, ROIs were determined using a Bonferroni corrected statistical threshold, and a minimum cluster size of 0.3 ml.
DTI Data Processing
DTI data processing was performed using the FMRIB’s Diffusion Toolbox as implemented in the FSL software package version 4.1.9 (FMRIB Analysis group, Oxford, UK (32)). After eddy current and motion correction, all volumes for each child were screened for the presence of artifacts. If an artifact was present within a volume, this volume was removed for this child. For all included children, on average 96.6% (range 71.4–100%) of all volumes were found to be suitable for DTI analyses. After that, analyses were conducted using default settings of bedpostx from the FMRIB’s Diffusion Toolbox. Probabilistic fiber tracking was performed using the ROIs as derived from the fMRI outcomes as seeding regions. First, coordinates of each voxel of a ROI were transposed from Talairach space into Montreal Neurological Institute space by the Lancaster method (33) using the GingerALE software package (Ginger ALE, version 2.0.4; http://www.brainmap.org/) (34) and imported in FSL. Second, the ROI in Montreal Neurological Institute space was dilated 1 mm in every direction to make sure that the area of contact between neuronal gray matter and their innervating white matter fiber tracts was included in the seeding region. The ROIs were transformed from Montreal Neurological Institute space into subject DTI space using the inverse nonlinear transformation needed to register individual FA maps to Montreal Neurological Institute space. Third, the regions of the left and right thalamus were determined using the 3D T1 series of each child using the FIRST tool provided by FSL. These thalamus ROIs were also transformed into subject DTI space. Finally, path tracing was performed, using a total of 5,000 permutations for each voxel, from the seeding regions toward the left and right thalamus, and vice versa. For analyses, a threshold was set for each voxel to include at least 15% of the total number of fibers in the tract, in order to limit the possibility that fiber tract connections between the seeding region and the thalamus were erroneously included by chance. For each participant, mean volume, mean FA value, and the mean of the eigenvalues L1 and L23 (the mean of L2 and L3) were derived from all tracts between the left and right thalamus and seeding regions.
Statistical Analyses
Statistical analyses were performed using PASW Statistics 20.0 (SPSS, Chicago, IL). Independent t-test (for normally distributed continuous data) and χ2 test (for dichotomous data) were performed to investigate whether there were differences between very preterm children and term controls in terms of age, socio economic status, full-scale IQ, and gender, respectively. Group differences were investigated using analysis of variance for (i) RT during the three conditions of the Flanker task, (ii) beta weights of mean activation of each involved ROI in interference control, and (iii) volume, mean FA, L1, and L23 values of fiber tracts innervating the cortical regions involved in interference control. Finally, Pearson correlations were determined between beta weights of mean activation, mean FA value, and the interference effect, to signify the functional relations between cortical activation, differences in white matter development, and interference control separately for very preterm children and term controls. Group differences were analyzed using an alpha of 0.05 and quantified in terms of effect sizes (Cohen’s d) (35).
Statement of Financial Support
This work was supported by an unrestricted research grant from Danone, Friederichsdorf, Germany. Danone had no involvement in (i) the study design; (ii) collection, analysis, and interpretation of data; (iii) preparation of the current paper; and (iv) the decision to submit the current paper for publication. Scanning costs were in part funded by the Amsterdam Brain Imaging Platform (ABIP), Amsterdam, The Netherlands.
Disclosure
None of the authors have any conflict of interest.
References
de Kieviet JF, Zoetebier L, van Elburg RM, Vermeulen RJ, Oosterlaan J . Brain development of very preterm and very low-birthweight children in childhood and adolescence: a meta-analysis. Dev Med Child Neurol 2012;54:313–23.
Aarnoudse-Moens CS, Weisglas-Kuperus N, van Goudoever JB, Oosterlaan J . Meta-analysis of neurobehavioral outcomes in very preterm and/or very low birth weight children. Pediatrics 2009;124:717–28.
Bhutta AT, Cleves MA, Casey PH, Cradock MM, Anand KJ . Cognitive and behavioral outcomes of school-aged children who were born preterm: a meta-analysis. JAMA 2002;288:728–37.
Mulder H, Pitchford NJ, Hagger MS, Marlow N . Development of executive function and attention in preterm children: a systematic review. Dev Neuropsychol 2009;34:393–421.
Van de Weijer-Bergsma E, Wijnroks L, Jongmans MJ . Attention development in infants and preschool children born preterm: a review. Infant Behav Dev 2008;31:333–51.
Nigg JT . On inhibition/disinhibition in developmental psychopathology: views from cognitive and personality psychology and a working inhibition taxonomy. Psychol Bull 2000;126:220–46.
Bunge SA, Hazeltine E, Scanlon MD, Rosen AC, Gabrieli JD . Dissociable contributions of prefrontal and parietal cortices to response selection. Neuroimage 2002;17:1562–71.
Bush G, Vogt BA, Holmes J, et al. Dorsal anterior cingulate cortex: a role in reward-based decision making. Proc Natl Acad Sci USA 2002;99:523–8.
Fan J, McCandliss BD, Fossella J, Flombaum JI, Posner MI . The activation of attentional networks. Neuroimage 2005;26:471–9.
Lawrence EJ, Rubia K, Murray RM, et al. The neural basis of response inhibition and attention allocation as mediated by gestational age. Hum Brain Mapp 2009;30:1038–50.
Ment LR, Constable RT . Injury and recovery in the developing brain: evidence from functional MRI studies of prematurely born children. Nat Clin Pract Neurol 2007;3:558–71.
Narberhaus A, Lawrence E, Allin MP, et al. Neural substrates of visual paired associates in young adults with a history of very preterm birth: alterations in fronto-parieto-occipital networks and caudate nucleus. Neuroimage 2009;47:1884–93.
Nosarti C, Rubia K, Smith AB, et al. Altered functional neuroanatomy of response inhibition in adolescent males who were born very preterm. Dev Med Child Neurol 2006;48:265–71.
Constable RT, Ment LR, Vohr BR, et al. Prematurely born children demonstrate white matter microstructural differences at 12 years of age, relative to term control subjects: an investigation of group and gender effects. Pediatrics 2008;121:306–16.
Kontis D, Catani M, Cuddy M, et al. Diffusion tensor MRI of the corpus callosum and cognitive function in adults born preterm. Neuroreport 2009;20:424–8.
Skranes J, Vangberg TR, Kulseng S, et al. Clinical findings and white matter abnormalities seen on diffusion tensor imaging in adolescents with very low birth weight. Brain 2007;130(Pt 3):654–66.
Vangberg TR, Skranes J, Dale AM, Martinussen M, Brubakk AM, Haraldseth O . Changes in white matter diffusion anisotropy in adolescents born prematurely. Neuroimage 2006;32:1538–48.
Volpe JJ . Cerebral white matter injury of the premature infant-more common than you think. Pediatrics 2003;112(1 Pt 1):176–80.
Taylor HG, Filipek PA, Juranek J, Bangert B, Minich N, Hack M . Brain volumes in adolescents with very low birth weight: effects on brain structure and associations with neuropsychological outcomes. Dev Neuropsychol 2011;36:96–117.
Ligam P, Haynes RL, Folkerth RD, et al. Thalamic damage in periventricular leukomalacia: novel pathologic observations relevant to cognitive deficits in survivors of prematurity. Pediatr Res 2009;65:524–9.
Wheeler-Kingshott CA, Cercignani M . About “axial” and “radial” diffusivities. Magn Reson Med 2009;61:1255–60.
Thomas B, Eyssen M, Peeters R, et al. Quantitative diffusion tensor imaging in cerebral palsy due to periventricular white matter injury. Brain 2005;128(Pt 11):2562–77.
Feldman HM, Lee ES, Loe IM, Yeom KW, Grill-Spector K, Luna B . White matter microstructure on diffusion tensor imaging is associated with conventional magnetic resonance imaging findings and cognitive function in adolescents born preterm. Dev Med Child Neurol 2012;54:809–14.
Giménez M, Miranda MJ, Born AP, Nagy Z, Rostrup E, Jernigan TL . Accelerated cerebral white matter development in preterm infants: a voxel-based morphometry study with diffusion tensor MR imaging. Neuroimage 2008;41:728–34.
Allin MP, Kontis D, Walshe M, et al. White matter and cognition in adults who were born preterm. PLoS One 2011;6:e24525.
van Kooij BJ, van Pul C, Benders MJ, van Haastert IC, de Vries LS, Groenendaal F . Fiber tracking at term displays gender differences regarding cognitive and motor outcome at 2 years of age in preterm infants. Pediatr Res 2011;70:626–32.
Fjell AM, Walhovd KB, Brown TT, et al.; Pediatric Imaging, Neurocognition, and Genetics Study. Multimodal imaging of the self-regulating developing brain. Proc Natl Acad Sci USA 2012;109:19620–5.
van den Berg A, van Elburg RM, Westerbeek EA, Twisk JW, Fetter WP . Glutamine-enriched enteral nutrition in very-low-birth-weight infants and effects on feeding tolerance and infectious morbidity: a randomized controlled trial. Am J Clin Nutr 2005;81:1397–404.
Wechsler D . WISC-III Manual. London, UK: The Psychological Corporation, 2002.
de Bie HM, Boersma M, Wattjes MP, et al. Preparing children with a mock scanner training protocol results in high quality structural and functional MRI scans. Eur J Pediatr 2010;169:1079–85.
Eriksen CW . The flankers task and response competition: A useful tool for investigating a variety of cognitive problems. Vis Cogn 1995;2:101–18.
Woolrich MW, Jbabdi S, Patenaude B, et al. Bayesian analysis of neuroimaging data in FSL. Neuroimage 2009;45(Suppl):S173–86.
Lancaster JL, Tordesillas-Gutiérrez D, Martinez M, et al. Bias between MNI and Talairach coordinates analyzed using the ICBM-152 brain template. Hum Brain Mapp 2007;28:1194–205.
Eickhoff SB, Laird AR, Grefkes C, Wang LE, Zilles K, Fox PT . Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: a random-effects approach based on empirical estimates of spatial uncertainty. Hum Brain Mapp 2009;30:2907–26.
Cohen J . Statistical Power Analyses for the Behavioral Sciences, 2nd ed. Hillsdale, NY: Erlbaum, 1988.
Papile L-A,, Burstein J, Burstein R, Koffler H. Incidence and evolution of subependymal and intraventricular hemorrhage: A study of infants with birth weights less than 1,500 gm. J Pediatr 1978; 92:529–34.
Author information
Authors and Affiliations
Corresponding author
PowerPoint slides
Rights and permissions
About this article
Cite this article
de Kieviet, J., Heslenfeld, D., Pouwels, P. et al. A crucial role for white matter alterations in interference control problems of very preterm children. Pediatr Res 75, 731–737 (2014). https://doi.org/10.1038/pr.2014.31
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/pr.2014.31
This article is cited by
-
Executive function training in very preterm children: a randomized controlled trial
European Child & Adolescent Psychiatry (2021)
-
Altered brain metabolism contributes to executive function deficits in school-aged children born very preterm
Pediatric Research (2020)
-
Nurturing the preterm infant brain: leveraging neuroplasticity to improve neurobehavioral outcomes
Pediatric Research (2019)
