
Key Points
-
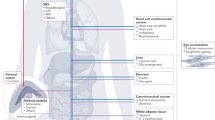
During stress there is activation of the hypothalamic-pituitary-adrenal (HPA) axis, culminating in the production of glucocorticoids. Glucocorticoids can easily access the brain, where they bind to receptors and influence the brain and behaviour.
-
Different outcomes result from exposure to stress at different periods of an individual's life.
-
Exposure to stress in the prenatal period leads to programming effects, as evidenced by increased reactivity to stress later in life and reduced hippocampal volume in adulthood.
-
Exposure to prenatal stress has been associated with learning impairments, enhanced sensitivity to drugs of abuse, and increases in anxiety and depression-related behaviours in adulthood.
-
Maternal separation is a potent stressor in the postnatal period, and it leads to increased secretion of glucocorticoids that can extend into adulthood. By contrast, exposure to severe abuse during infancy is associated with lower levels of glucocorticoids in both primates and humans.
-
Stress during adolescence has more important effects on the HPA axis than a similar stress exposure during adulthood. Moreover, the effects of stress during adolescence can incubate until adulthood, at which time they will become apparent.
-
The effects of stress exposure on the brain and behaviour in adulthood are similar to those that are observed in childhood and adolescence. However, unlike these latter effects, the former effects are reversible; that is, they usually disappear after cessation of the stressor.
-
In adulthood, chronic exposure to high levels of glucocorticoids has been associated with depressive disorder. By contrast, patients with post-traumatic stress disorder present lower levels of glucocorticoids.
-
The effects of stress during aging are associated with both memory impairments and reduced hippocampal volumes.
-
The life cycle model of stress explains why different disorders emerge in populations exposed to stress at different stages of their lives.
Abstract
Chronic exposure to stress hormones, whether it occurs during the prenatal period, infancy, childhood, adolescence, adulthood or aging, has an impact on brain structures involved in cognition and mental health. However, the specific effects on the brain, behaviour and cognition emerge as a function of the timing and the duration of the exposure, and some also depend on the interaction between gene effects and previous exposure to environmental adversity. Advances in animal and human studies have made it possible to synthesize these findings, and in this Review a model is developed to explain why different disorders emerge in individuals exposed to stress at different times in their lives.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others

References
Barker, D. J. The foetal and infant origins of inequalities in health in Britain. J. Public Health Med. 13, 64–68 (1991).
Cadet, R., Pradier, P., Dalle, M. & Delost, P. Effects of prenatal maternal stress on the pituitary adrenocortical reactivity in guinea-pig pups. J. Dev. Physiol. 8, 467–475 (1986).
Dean, F. & Matthews, S. G. Maternal dexamethasone treatment in late gestation alters glucocorticoid and mineralocorticoid receptor mRNA in the fetal guinea pig brain. Brain Res. 846, 253–259 (1999).
Seckl, J. R. Glucocorticoids, developmental 'programming' and the risk of affective dysfunction. Prog. Brain Res. 167, 17–34 (2008). A superb review that summarized prenatal work and linked it to clinical implications.
Koehl, M. et al. Prenatal stress alters circadian activity of hypothalamo-pituitary-adrenal axis and hippocampal corticosteroid receptors in adult rats of both gender. J. Neurobiol. 40, 302–315 (1999).
Barbazanges, A., Piazza, P. V., Le Moal, M. & Maccari, S. Maternal glucocorticoid secretion mediates long-term effects of prenatal stress. J. Neurosci. 16, 3943–3949 (1996).
Meyer, J. S. Early adrenalectomy stimulates subsequent growth and development of the rat brain. Exp. Neurol. 82, 432–446 (1983).
Weaver, I. C. et al. Epigenetic programming by maternal behavior. Nature Neurosci. 7, 847–854 (2004). The first paper to show that early experience has epigenetic effects, altering methylation patterns.
Uno, H. et al. Brain damage induced by prenatal exposure to dexamethasone in fetal rhesus macaques. I. Hippocampus. Brain Res. Dev. Brain Res. 53, 157–167 (1990).
Murmu, M. S. et al. Changes of spine density and dendritic complexity in the prefrontal cortex in offspring of mothers exposed to stress during pregnancy. Eur. J. Neurosci. 24, 1477–1487 (2006).
Cratty, M. S., Ward, H. E., Johnson, E. A., Azzaro, A. J. & Birkle, D. L. Prenatal stress increases corticotropin-releasing factor (CRF) content and release in rat amygdala minces. Brain Res. 675, 297–302 (1995).
Vallee, M. et al. Long-term effects of prenatal stress and postnatal handling on age-related glucocorticoid secretion and cognitive performance: a longitudinal study in the rat. Eur. J. Neurosci. 11, 2906–2916 (1999).
Deminiere, J. M. et al. Increased locomotor response to novelty and propensity to intravenous amphetamine self-administration in adult offspring of stressed mothers. Brain Res. 586, 135–139 (1992).
Vallee, M. et al. Prenatal stress induces high anxiety and postnatal handling induces low anxiety in adult offspring: correlation with stress-induced corticosterone secretion. J. Neurosci. 17, 2626–2636 (1997).
Lemaire, V., Koehl, M., Le Moal, M. & Abrous, D. N. Prenatal stress produces learning deficits associated with an inhibition of neurogenesis in the hippocampus. Proc. Natl Acad. Sci. USA 97, 11032–11037 (2000).
Piazza, P. V. & Le Moal, M. L. Pathophysiological basis of vulnerability to drug abuse: role of an interaction between stress, glucocorticoids, and dopaminergic neurons. Annu. Rev. Pharmacol. Toxicol. 36, 359–378 (1996).
Kapoor, A., Petropoulos, S. & Matthews, S. G. Fetal programming of hypothalamic-pituitary-adrenal (HPA) axis function and behavior by synthetic glucocorticoids. Brain Res. Rev. 57, 586–595 (2008).
Hedegaard, M., Henriksen, T. B., Sabroe, S. & Secher, N. J. Psychological distress in pregnancy and preterm delivery. BMJ 307, 234–239 (1993).
Orr, S. T. & Miller, C. A. Maternal depressive symptoms and the risk of poor pregnancy outcome. Review of the literature and preliminary findings. Epidemiol. Rev. 17, 165–171 (1995).
Lyons-Ruth, K., Wolfe, R. & Lyubchik, A. Depression and the parenting of young children: making the case for early preventive mental health services. Harv. Rev. Psychiatry 8, 148–153 (2000).
Gutteling, B. M., de Weerth, C. & Buitelaar, J. K. Prenatal stress and children's cortisol reaction to the first day of school. Psychoneuroendocrinology 30, 541–549 (2005).
O'Connor, T. G. et al. Prenatal anxiety predicts individual differences in cortisol in pre-adolescent children. Biol. Psychiatry 58, 211–217 (2005).
Glover, V. Maternal stress or anxiety in pregnancy and emotional development of the child. Br. J. Psychiatry 171, 105–106 (1997).
Stott, D. H. Follow-up study from birth of the effects of prenatal stresses. Dev. Med. Child. Neurol. 15, 770–787 (1973).
Trautman, P. D., Meyer-Bahlburg, H. F., Postelnek, J. & New, M. I. Effects of early prenatal dexamethasone on the cognitive and behavioral development of young children: results of a pilot study. Psychoneuroendocrinology 20, 439–449 (1995).
Buss, C. et al. Maternal care modulates the relationship between prenatal risk and hippocampal volume in women but not in men. J. Neurosci. 27, 2592–2595 (2007).
Levine, S. & Wiener, S. G. Psychoendocrine aspects of mother-infant relationships in nonhuman primates. Psychoneuroendocrinology 13, 143–154 (1988).
Anisman, H., Zaharia, M. D., Meaney, M. J. & Merali, Z. Do early-life events permanently alter behavioral and hormonal responses to stressors? Int. J. Dev. Neurosci. 16, 149–164 (1998).
Liu, D. et al. Maternal care, hippocampal glucocorticoid receptors, and hypothalamic-pituitary-adrenal responses to stress. Science 277, 1659–1662 (1997).
Fenoglio, K. A., Brunson, K. L. & Baram, T. Z. Hippocampal neuroplasticity induced by early-life stress: functional and molecular aspects. Front. Neuroendocrinol. 27, 180–192 (2006).
Schulkin, J., Gold, P. W. & McEwen, B. S. Induction of corticotropin-releasing hormone gene expression by glucocorticoids: implication for understanding the states of fear and anxiety and allostatic load. Psychoneuroendocrinology 23, 219–243 (1998).
de Kloet, E. R. & Oitzl, M. S. Who cares for a stressed brain? The mother, the kid or both? Neurobiol. Aging 24 (Suppl. 1), S61–S65; discussion S67–S68 (2003).
Sanchez, M. M. et al. Alterations in diurnal cortisol rhythm and acoustic startle response in nonhuman primates with adverse rearing. Biol. Psychiatry 57, 373–381 (2005).
Coplan, J. D. et al. Persistent elevations of cerebrospinal fluid concentrations of corticotropin-releasing factor in adult nonhuman primates exposed to early-life stressors: implications for the pathophysiology of mood and anxiety disorders. Proc. Natl Acad. Sci. USA 93, 1619–1623 (1996).
Sanchez, M. M. The impact of early adverse care on HPA axis development: nonhuman primate models. Horm. Behav. 50, 623–631 (2006).
Rosenblum, L. A. et al. Differing concentrations of corticotropin-releasing factor and oxytocin in the cerebrospinal fluid of bonnet and pigtail macaques. Psychoneuroendocrinology 27, 651–660 (2002).
Siegel, S. J. et al. Effects of social deprivation in prepubescent rhesus monkeys: immunohistochemical analysis of the neurofilament protein triplet in the hippocampal formation. Brain Res. 619, 299–305 (1993).
Sanchez, M. M., Ladd, C. O. & Plotsky, P. M. Early adverse experience as a developmental risk factor for later psychopathology: evidence from rodent and primate models. Dev. Psychopathol. 13, 419–449 (2001).
Gunnar, M. R. & Donzella, B. Social regulation of the cortisol levels in early human development. Psychoneuroendocrinology 27, 199–220 (2002).
Geoffroy, M. C., Cote, S. M., Parent, S. & Seguin, J. R. Daycare attendance, stress, and mental health. Can. J. Psychiatry 51, 607–615 (2006).
NICHD Early Child Care Research Network. Early child care and children's development prior to school entry: results from the NICHD Study of Early Child Care. Am. Educ. Res. J. 39, 133–164 (2002).
Albers, E. M., Riksen-Walraven, J. M., Sweep, F. C. & de Weerth, C. Maternal behavior predicts infant cortisol recovery from a mild everyday stressor. J. Child. Psychol. Psychiatry 49, 97–103 (2008).
Lupien, S. J., King, S., Meaney, M. J. & McEwen, B. S. Child's stress hormone levels correlate with mother's socioeconomic status and depressive state. Biol. Psychiatry 48, 976–980 (2000).
Halligan, S. L., Herbert, J., Goodyer, I. & Murray, L. Disturbances in morning cortisol secretion in association with maternal postnatal depression predict subsequent depressive symptomatology in adolescents. Biol. Psychiatry 62, 40–46 (2007). Provided some of the first evidence that adverse early life experiences in humans, in this case rearing by a mother suffering from post-partum depression, are associated with heightened HPA activity years later, and that the HPA axis hyperactivity mediates the association between early risk exposure and later psychiatric symptoms.
Jones, N. A., Field, T. & Davalos, M. Right frontal EEG asymmetry and lack of empathy in preschool children of depressed mothers. Child. Psychiatry Hum. Dev. 30, 189–204 (2000).
Fries, E., Hesse, J., Hellhammer, J. & Hellhammer, D. H. A new view on hypocortisolism. Psychoneuroendocrinology 30, 1010–1016 (2005).
Yehuda, R., Yang, R. K., Buchsbaum, M. S. & Golier, J. A. Alterations in cortisol negative feedback inhibition as examined using the ACTH response to cortisol administration in PTSD. Psychoneuroendocrinology 31, 447–451 (2006).
Gunnar, M. R. & Quevedo, K. M. Early care experiences and HPA axis regulation in children: a mechanism for later trauma vulnerability. Prog. Brain Res. 167, 137–149 (2008).
McGowan, P. O. et al. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nature Neurosci. 12, 342–348 (2009). This study examined epigenetic differences in a neuron-specific glucocorticoid receptor (NR3C1) promoter between post-mortem hippocampus obtained from suicide victims with a history of childhood abuse and hippocampus from either suicide victims with no childhood abuse or controls. It found decreased levels of glucocorticoid receptor mRNA, as well as mRNA transcripts bearing the glucocorticoid receptor 1F splice variant and increased cytosine methylation of an NR3C1 promoter in suicide victims with early abuse.
McCormick, C. M. & Mathews, I. Z. HPA function in adolescence: role of sex hormones in its regulation and the enduring consequences of exposure to stressors. Pharmacol. Biochem. Behav. 86, 220–233 (2007). A very good review on the acute and chronic effects of stress during adolescence.
O'Donnell, S., Noseworthy, M. D., Levine, B. & Dennis, M. Cortical thickness of the frontopolar area in typically developing children and adolescents. Neuroimage 24, 948–954 (2005).
Vazquez, D. M. & Akil, H. Pituitary-adrenal response to ether vapor in the weanling animal: characterization of the inhibitory effect of glucocorticoids on adrenocorticotropin secretion. Pediatr. Res. 34, 646–653 (1993).
Goldman, L., Winget, C., Hollingshead, G. W. & Levine, S. Postweaning development of negative feedback in the pituitary-adrenal system of the rat. Neuroendocrinology 12, 199–211 (1973).
Girotti, M. et al. Habituation to repeated restraint stress is associated with lack of stress-induced c-fos expression in primary sensory processing areas of the rat brain. Neuroscience 138, 1067–1081 (2006).
Romeo, R. D. et al. Stress history and pubertal development interact to shape hypothalamic-pituitary-adrenal axis plasticity. Endocrinology 147, 1664–1674 (2006).
Avital, A. & Richter-Levin, G. Exposure to juvenile stress exacerbates the behavioural consequences of exposure to stress in the adult rat. Int. J. Neuropsychopharmacol. 8, 163–173 (2005).
Lee, P. R., Brady, D. & Koenig, J. I. Corticosterone alters N-methyl-D-aspartate receptor subunit mRNA expression before puberty. Brain Res. Mol. Brain Res. 115, 55–62 (2003).
Isgor, C., Kabbaj, M., Akil, H. & Watson, S. J. Delayed effects of chronic variable stress during peripubertal-juvenile period on hippocampal morphology and on cognitive and stress axis functions in rats. Hippocampus 14, 636–648 (2004). One of the first papers to show protracted effects of adolescent stress on adulthood stress reactivity in rodents.
Tsoory, M. & Richter-Levin, G. Learning under stress in the adult rat is differentially affected by 'juvenile' or 'adolescent' stress. Int. J. Neuropsychopharmacol. 9, 713–728 (2006).
Kabbaj, M., Isgor, C., Watson, S. J. & Akil, H. Stress during adolescence alters behavioral sensitization to amphetamine. Neuroscience 113, 395–400 (2002).
McCormick, C. M., Robarts, D., Gleason, E. & Kelsey, J. E. Stress during adolescence enhances locomotor sensitization to nicotine in adulthood in female, but not male, rats. Horm. Behav. 46, 458–466 (2004).
Gunnar, M. R., Wewerka, S., Frenn, K., Long, J. D. & Griggs, C. Developmental changes in hypothalamus-pituitary-adrenal activity over the transition to adolescence: normative changes and associations with puberty. Dev. Psychopathol. 21, 69–85 (2009).
Giedd, J. N. et al. Quantitative magnetic resonance imaging of human brain development: ages 4–18. Cereb. Cortex 6, 551–560 (1996).
Perlman, W. R., Webster, M. J., Herman, M. M., Kleinman, J. E. & Weickert, C. S. Age-related differences in glucocorticoid receptor mRNA levels in the human brain. Neurobiol. Aging 28, 447–458 (2007).
Dahl, R. E. Adolescent brain development: a period of vulnerabilities and opportunities. Keynote address. Ann. NY Acad. Sci. 1021, 1–22 (2004).
Paus, T., Keshavan, M. & Giedd, J. N. Why do many psychiatric disorders emerge during adolescence? Nature Rev. Neurosci. 9, 947–957 (2008). A very interesting review on the state of research into why adolescents have a greater vulnerability to mental health disorders.
Evans, G. W. & English, K. The environment of poverty: multiple stressor exposure, psychophysiological stress, and socioemotional adjustment. Child Dev. 73, 1238–1248 (2002).
Andersen, S. L. & Teicher, M. H. Stress, sensitive periods and maturational events in adolescent depression. Trends Neurosci. 31, 183–191 (2008).
De Bellis, M. D. et al. A. E. Bennett Research Award. Developmental traumatology. Part II: brain development. Biol. Psychiatry 45, 1271–1284 (1999). One of the first clear demonstrations that, in children who were physically healthy at birth, severe abuse in the early years of life is associated with reduced brain volume. The reduction correlates negatively with the age of onset and positively with the duration of the maltreatment.
Cohen, R. A. et al. Early life stress and morphometry of the adult anterior cingulate cortex and caudate nuclei. Biol. Psychiatry 59, 975–982 (2006).
Diamond, D. M., Bennett, M. C., Fleshner, M. & Rose, G. M. Inverted-U relationship between the level of peripheral corticosterone and the magnitude of hippocampal primed burst potentiation. Hippocampus 2, 421–430 (1992).
Vouimba, R. M., Yaniv, D. & Richter-Levin, G. Glucocorticoid receptors and β-adrenoceptors in basolateral amygdala modulate synaptic plasticity in hippocampal dentate gyrus, but not in area CA1. Neuropharmacology 52, 244–252 (2007).
Roozendaal, B., Brunson, K. L., Holloway, B. L., McGaugh, J. L. & Baram, T. Z. Involvement of stress-released corticotropin-releasing hormone in the basolateral amygdala in regulating memory consolidation. Proc. Natl Acad. Sci. USA 99, 13908–13913 (2002).
Magarinos, A. M. & McEwen, B. S. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: involvement of glucocorticoid secretion and excitatory amino acid receptors. Neuroscience 69, 89–98 (1995).
Conrad, C. D., LeDoux, J. E., Magarinos, A. M. & McEwen, B. S. Repeated restraint stress facilitates fear conditioning independently of causing hippocampal CA3 dendritic atrophy. Behav. Neurosci. 113, 902–913 (1999).
Gould, E., McEwen, B. S., Tanapat, P., Galea, L. A. & Fuchs, E. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. J. Neurosci. 17, 2492–2498 (1997).
McEwen, B. S. Effects of adverse experiences for brain structure and function. Biol. Psychiatry 48, 721–731 (2000).
Pham, K., Nacher, J., Hof, P. R. & McEwen, B. S. Repeated restraint stress suppresses neurogenesis and induces biphasic PSA-NCAM expression in the adult rat dentate gyrus. Eur. J. Neurosci. 17, 879–886 (2003).
McEwen, B. S. Plasticity of the hippocampus: adaptation to chronic stress and allostatic load. Ann. NY Acad. Sci. 933, 265–277 (2001).
Luine, V., Villegas, M., Martinez, C. & McEwen, B. S. Repeated stress causes reversible impairments of spatial memory performance. Brain Res. 639, 167–170 (1994).
Joels, M., Karst, H., Krugers, H. J. & Lucassen, P. J. Chronic stress: implications for neuronal morphology, function and neurogenesis. Front. Neuroendocrinol. 28, 72–96 (2007).
Izquierdo, A., Wellman, C. L. & Holmes, A. Brief uncontrollable stress causes dendritic retraction in infralimbic cortex and resistance to fear extinction in mice. J. Neurosci. 26, 5733–5738 (2006).
Shansky, R. M., Hamo, C., Hof, P. R., McEwen, B. S. & Morrison, J. H. Stress-induced dendritic remodeling in the prefrontal cortex is circuit specific. Cereb. Cortex 4 Feb 2009 (doi:10.1093/cercor/bhp003).
Cerqueira, J. J. et al. Corticosteroid status influences the volume of the rat cingulate cortex - a magnetic resonance imaging study. J. Psychiatr. Res. 39, 451–460 (2005).
Mitra, R., Jadhav, S., McEwen, B. S., Vyas, A. & Chattarji, S. Stress duration modulates the spatiotemporal patterns of spine formation in the basolateral amygdala. Proc. Natl Acad. Sci. USA 102, 9371–9376 (2005).
Mitra, R. & Sapolsky, R. M. Acute corticosterone treatment is sufficient to induce anxiety and amygdaloid dendritic hypertrophy. Proc. Natl Acad. Sci. USA 105, 5573–5578 (2008). This interesting study addressed endocrine effects on the brain, with a focus on the amygdala and anxiety (rather than on hippocampus and memory). Of note, a single dose of glucocorticoids was sufficient to induce changes in amygdala structure 10 days later, which might be useful to model in animals PTSD.
Lupien, S. J. & McEwen, B. S. The acute effects of corticosteroids on cognition: integration of animal and human model studies. Brain Res. Brain Res. Rev. 24, 1–27 (1997).
Roozendaal, B. Glucocorticoids and the regulation of memory consolidation. Psychoneuroendocrinology 25, 213–238 (2000).
Lupien, S. J. et al. Stress hormones and human memory function across the lifespan. Psychoneuroendocrinology 30, 225–242 (2005).
Lupien, S. J. et al. Hippocampal volume is as variable in young as in older adults: implications for the notion of hippocampal atrophy in humans. Neuroimage 34, 479–485 (2007). This study showed that ∼25% of young adults present hippocampal volumes as small as those of older adults. The presence of small hippocampal volumes in healthy young individuals supports the vulnerability hypothesis.
Pruessner, J. C., Lord, C., Meaney, M. & Lupien, S. Effects of self-esteem on age-related changes in cognition and the regulation of the hypothalamic-pituitary-adrenal axis. Ann. NY Acad. Sci. 1032, 186–194 (2004).
Pruessner, J. C. et al. Self-esteem, locus of control, hippocampal volume, and cortisol regulation in young and old adulthood. Neuroimage 28, 815–826 (2005).
Burke, H. M., Davis, M. C., Otte, C. & Mohr, D. C. Depression and cortisol responses to psychological stress: a meta-analysis. Psychoneuroendocrinology 30, 846–856 (2005).
Yehuda, R., Golier, J. A. & Kaufman, S. Circadian rhythm of salivary cortisol in Holocaust survivors with and without PTSD. Am. J. Psychiatry 162, 998–1000 (2005).
Meewisse, M. L., Reitsma, J. B., de Vries, G. J., Gersons, B. P. & Olff, M. Cortisol and post-traumatic stress disorder in adults: systematic review and meta-analysis. Br. J. Psychiatry 191, 387–392 (2007). This paper presented the first meta-analysis of cortisol findings in PTSD, to elucidate the determinants of hypocortisolism and resolve the inconsistency in findings.
Heim, C. et al. Pituitary-adrenal and autonomic responses to stress in women after sexual and physical abuse in childhood. JAMA 284, 592–597 (2000).
Heim, C., Mletzko, T., Purselle, D., Musselman, D. L. & Nemeroff, C. B. The dexamethasone/corticotropin-releasing factor test in men with major depression: role of childhood trauma. Biol. Psychiatry 63, 398–405 (2008).
Carpenter, L. L. et al. Cerebrospinal fluid corticotropin-releasing factor and perceived early-life stress in depressed patients and healthy control subjects. Neuropsychopharmacology 29, 777–784 (2004).
Heim, C., Newport, D. J., Mletzko, T., Miller, A. H. & Nemeroff, C. B. The link between childhood trauma and depression: insights from HPA axis studies in humans. Psychoneuroendocrinology 33, 693–710 (2008). A crucially important review which documents that the disturbances in the HPA axis that are observed in many adults with depression may be specific to those who experienced trauma or maltreatment in childhood.
Videbech, P. & Ravnkilde, B. Hippocampal volume and depression: a meta-analysis of MRI studies. Am. J. Psychiatry 161, 1957–1966 (2004).
Smith, M. E. Bilateral hippocampal volume reduction in adults with post-traumatic stress disorder: a meta-analysis of structural MRI studies. Hippocampus 15, 798–807 (2005).
Vythilingam, M. et al. Childhood trauma associated with smaller hippocampal volume in women with major depression. Am. J. Psychiatry 159, 2072–2080 (2002).
Gilbertson, M. W. et al. Smaller hippocampal volume predicts pathologic vulnerability to psychological trauma. Nature Neurosci. 5, 1242–1247 (2002). The first paper to study whether the reduced hippocampal volume observed in PTSD patients is due to the disorder, to trauma exposure or to a pre-existing factor.
Issa, A. M., Rowe, W., Gauthier, S. & Meaney, M. J. Hypothalamic-pituitary-adrenal activity in aged, cognitively impaired and cognitively unimpaired rats. J. Neurosci. 10, 3247–3254 (1990).
Landfield, P. W., Waymire, J. C. & Lynch, G. Hippocampal aging and adrenocorticoids: quantitative correlations. Science 202, 1098–1102 (1978).
Landfield, P. W., Baskin, R. K. & Pitler, T. A. Brain aging correlates: retardation by hormonal-pharmacological treatments. Science 214, 581–584 (1981). The first study to show that chronic exposure to high levels of glucocorticoids in rodents is associated with memory impairments and reduced hippocampal volume.
Landfield, P. W., Blalock, E. M., Chen, K. C. & Porter, N. M. A new glucocorticoid hypothesis of brain aging: implications for Alzheimer's disease. Curr. Alzheimer Res. 4, 205–212 (2007).
Kulstad, J. J. et al. Effects of chronic glucocorticoid administration on insulin-degrading enzyme and amyloid-β peptide in the aged macaque. J. Neuropathol. Exp. Neurol. 64, 139–146 (2005).
Sapolsky, R. M., Krey, L. C. & McEwen, B. S. The neuroendocrinology of stress and aging: the glucocorticoid cascade hypothesis. Endocr. Rev. 7, 284–301 (1986). The first paper to present the glucocorticoid cascade hypothesis, now referred to as the neurotoxicity hypothesis.
Lowy, M. T., Wittenberg, L. & Yamamoto, B. K. Effect of acute stress on hippocampal glutamate levels and spectrin proteolysis in young and aged rats. J. Neurochem. 65, 268–274 (1995).
Raskind, M. A., Peskind, E. R. & Wilkinson, C. W. Hypothalamic-pituitary-adrenal axis regulation and human aging. Ann. NY Acad. Sci. 746, 327–335 (1994).
Lupien, S. J. et al. Cortisol levels during human aging predict hippocampal atrophy and memory deficits. Nature Neurosci. 1, 69–73 (1998).
Giubilei, F. et al. Altered circadian cortisol secretion in Alzheimer's disease: clinical and neuroradiological aspects. J. Neurosci. Res. 66, 262–265 (2001).
Aisen, P. S. et al. A randomized controlled trial of prednisone in Alzheimer's disease. Alzheimer's Disease Cooperative Study. Neurology 54, 588–593 (2000).
Dai, J., Buijs, R. & Swaab, D. Glucocorticoid hormone (cortisol) affects axonal transport in human cortex neurons but shows resistance in Alzheimer's disease. Br. J. Pharmacol. 143, 606–610 (2004).
Chen, Y., Dube, C. M., Rice, C. J. & Baram, T. Z. Rapid loss of dendritic spines after stress involves derangement of spine dynamics by corticotropin-releasing hormone. J. Neurosci. 28, 2903–2911 (2008).
Charney, D. S. & Manji, H. K. Life stress, genes, and depression: multiple pathways lead to increased risk and new opportunities for intervention. Sci. STKE 2004, re5 (2004).
Maercker, A., Michael, T., Fehm, L., Becker, E. S. & Margraf, J. Age of traumatisation as a predictor of post-traumatic stress disorder or major depression in young women. Br. J. Psychiatry 184, 482–487 (2004).
Teicher, M. H., Tomoda, A. & Andersen, S. L. Neurobiological consequences of early stress and childhood maltreatment: are results from human and animal studies comparable? Ann. NY Acad. Sci. 1071, 313–323 (2006).
Hall, F. S. Social deprivation of neonatal, adolescent, and adult rats has distinct neurochemical and behavioral consequences. Crit. Rev. Neurobiol. 12, 129–162 (1998).
Andersen, S. L. Trajectories of brain development: point of vulnerability or window of opportunity? Neurosci. Biobehav. Rev. 27, 3–18 (2003). A superb review paper which suggested that trauma at different time points during early development might be associated with different outcomes, depending on the brain structure that was affected at the time of exposure to adversity.
Widom, C. S., DuMont, K. & Czaja, S. J. A prospective investigation of major depressive disorder and comorbidity in abused and neglected children grown up. Arch. Gen. Psychiatry 64, 49–56 (2007).
Clayton, N. S. & Krebs, J. R. Hippocampal growth and attrition in birds affected by experience. Proc. Natl Acad. Sci. USA 91, 7410–7414 (1994).
Kudielka, B. M., Buske-Kirschbaum, A., Hellhammer, D. H. & Kirschbaum, C. HPA axis responses to laboratory psychosocial stress in healthy elderly adults, younger adults, and children: impact of age and gender. Psychoneuroendocrinology 29, 83–98 (2004).
Kessler, R. C. Epidemiology of women and depression. J. Affect. Disord. 74, 5–13 (2003).
Harlow, B. L., Cohen, L. S., Otto, M. W., Spiegelman, D. & Cramer, D. W. Early life menstrual characteristics and pregnancy experiences among women with and without major depression: the Harvard study of moods and cycles. J. Affect. Disord. 79, 167–176 (2004).
Zabin, L. S., Emerson, M. R. & Rowland, D. L. Childhood sexual abuse and early menarche: the direction of their relationship and its implications. J. Adolesc. Health 36, 393–400 (2005).
Jones, K. C. & de Voogt, P. Persistent organic pollutants (POPs): state of the science. Environ. Pollut. 100, 209–221 (1999).
Centers for Disease Control and Prevention. Second National Report on Human Exposure to Environmental Chemicals. (CDC, Atlanta, Georgia, 2003).
Daston, G. P., Cook, J. C. & Kavlock, R. J. Uncertainties for endocrine disrupters: our view on progress. Toxicol. Sci. 74, 245–252 (2003).
Gump, B. B. et al. Low-level prenatal and postnatal blood lead exposure and adrenocortical responses to acute stress in children. Environ. Health Perspect. 116, 249–255 (2008).
Denham, M. et al. Relationship of lead, mercury, mirex, dichlorodiphenyldichloroethylene, hexachlorobenzene, and polychlorinated biphenyls to timing of menarche among Akwesasne Mohawk girls. Pediatrics 115, e127–e134 (2005).
Turek, F. W. From circadian rhythms to clock genes in depression. Int. Clin. Psychopharmacol. 22 (Suppl. 2), S1–S8 (2007).
Lamarche, L. J. & De Koninck, J. Sleep disturbance in adults with posttraumatic stress disorder: a review. J. Clin. Psychiatry 68, 1257–1270 (2007).
Antoch, M. P. et al. Functional identification of the mouse circadian Clock gene by transgenic BAC rescue. Cell 89, 655–667 (1997).
Yakovlev, P. L. & Lecours, A. R. in Regional Development of the Brain in Early Life (ed. Minkowski, A.) 3–70 (Blackwell, Oxford, 1967).
Pruessner, J. C. et al. Volumetry of hippocampus and amygdala with high-resolution MRI and three-dimensional analysis software: minimizing the discrepancies between laboratories. Cereb. Cortex 10, 433–442 (2000).
Tisserand, D. J. et al. Regional frontal cortical volumes decrease differentially in aging: an MRI study to compare volumetric approaches and voxel-based morphometry. Neuroimage 17, 657–669 (2002).
Insel, T. R., Battaglia, G., Fairbanks, D. W. & De Souza, E. B. The ontogeny of brain receptors for corticotropin-releasing factor and the development of their functional association with adenylate cyclase. J. Neurosci. 8, 4151–4158 (1988).
Levine, S. The ontogeny of the hypothalamic-pituitary-adrenal axis. The influence of maternal factors. Ann. NY Acad. Sci. 746, 275–288; discussion 289–293 (1994).
Gunnar, M. R. & Cheatham, C. L. Brain and behavior interfaces: stress and the developing brain. Infant Ment. Health J. 24, 195–211 (2003). A superb paper that summarized the effects of stress during development and how this knowledge can be used to develop effective interventions.
LeDoux, J. E. The emotional brain: The mysterious underpinnings of emotional life (Simon & Schuster, New York, 1996).
Acknowledgements
Sonia Lupien holds a Research Chair on Gender and Mental Health by the Canadian Institutes of Health Research.
Author information
Authors and Affiliations
Corresponding author
Related links
Glossary
- Programming
-
When an environmental factor that acts during a sensitive developmental period affects the structure and function of tissues, leading to effects that persist throughout life.
- Mineralocorticoid receptor
-
A receptor that is activated by mineralocorticoids, such as aldosterone and deoxycorticosterone, as well as glucocorticoids, such as cortisol and cortisone. It also responds to progestins.
- Glucocorticoid receptor
-
A receptor that is activated by cortisol, corticosterone and other glucocorticoids and is expressed in almost every cell in the body. It regulates genes controlling development, metabolism and the immune response.
Rights and permissions
About this article
Cite this article
Lupien, S., McEwen, B., Gunnar, M. et al. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat Rev Neurosci 10, 434–445 (2009). https://doi.org/10.1038/nrn2639
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrn2639
This article is cited by
-
Effects of game-based physical education program on enjoyment in children and adolescents: a systematic review and meta-analysis
BMC Public Health (2024)
-
Emotion regulation shows an age- and sex-specific moderating effect on the relationship between chronic stress and cognitive performance
Scientific Reports (2024)
-
Changes in psychological and cognitive variables as well as cortisol levels in recovered Covid-19 patients: a longitudinal study
Current Psychology (2024)
-
Adverse childhood experiences and risk of late-life dementia: a systematic review and meta-analysis
Social Psychiatry and Psychiatric Epidemiology (2024)
-
Astaxanthin and DHA Supplementation Modulates the Maternal Undernutrition-induced Impairment of Cognitive Behavior and Synaptic Plasticity in Adult Life of Offspring’s -Exploring the Molecular Mechanism
Molecular Neurobiology (2024)
