
Key Points
-
Many Leishmania spp. can cause human disease, with clinical symptoms dependent upon both host- and parasite-related factors.
-
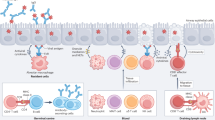
During infection, leishmanial parasites may be phagocytosed by a variety of cells, including neutrophils, monocytes, monocyte-derived dendritic cells (moDCs), macrophages and stromal cells. However, host cell preference and the importance of each cell type to parasite clearance versus parasite persistence may vary following infection by different Leishmania spp.
-
Intracellular Leishmania parasites have multiple ways to manipulate macrophage function, including subverting the control of phagosome biogenesis and maturation. The importance of parasite virulence factors (for example, lipophosphoglycan) in this process is dependent upon leishmanial species and host cell type.
-
Major surface protease (MSP; also known as GP63) of Leishmania spp. has an important role in regulating intracellular survival in some host cells, by cleaving a variety of phosphotyrosine phosphatases such as SRC homology 2 domain phosphotyrosine receptor phosphate (SHP1). MSP may access these cytosolic targets after crossing lipid microdomains in the host cell membrane.
-
Iron has a key role in intracellular survival of leishmanial pathogens, with both host (SLC11A1) and parasite (LIT1) transporters competing to secure this essential resource.
-
Cell-mediated immunity to leishmanial infection is multifactorial, with different models of disease serving to exemplify different aspects, for example, the importance of CD8+ T cells and/or the involvement of moDCs.
-
Parasite persistence and the capacity to induce good vaccine-induced immunity are both strongly influenced by the presence of interleukin-10 (IL-10). There are multiple cellular sources of IL-10 (T helper 1 (TH1) cells, regulatory T (TReg) cells, B cells, macrophages and DCs), but whether each has similar functional significance has yet to be determined.
Abstract
Leishmania is a genus of protozoan parasites that are transmitted by the bite of phlebotomine sandflies and give rise to a range of diseases (collectively known as leishmaniases) that affect over 150 million people worldwide. Cellular immune mechanisms have a major role in the control of infections with all Leishmania spp. However, as discussed in this Review, recent evidence suggests that each host–pathogen combination evokes different solutions to the problems of parasite establishment, survival and persistence. Understanding the extent of this diversity will be increasingly important in ensuring the development of broadly applicable vaccines, drugs and immunotherapeutic interventions.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


References
Kedzierski, L. Leishmaniasis vaccine: where are we today? J. Glob. Infect. Dis. 2, 177–185 (2010).
Peacock, C. S. et al. Comparative genomic analysis of three Leishmania species that cause diverse human disease. Nature Genet. 39, 839–847 (2007).
Kaye, P. M. & Blackwell, J. M. Postgenomic research on leishmaniasis: a critical self-appraisal. Trends Parasitol. 24, 401–405 (2008).
Akopyants, N. S. et al. Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector. Science 324, 265–268 (2009).
Dobson, D. E. et al. Leishmania major survival in selective Phlebotomus papatasi sand fly vector requires a specific SCG-encoded lipophosphoglycan galactosylation pattern. PLoS Pathog. 6, e1001185 (2010).
Sadlova, J. et al. The stage-regulated HASPB and SHERP proteins are essential for differentiation of the protozoan parasite Leishmania major in its sand fly vector, Phlebotomus papatasi. Cell. Microbiol. 12, 1765–1779 (2010).
Rogers, M. E., Ilg, T., Nikolaev, A. V., Ferguson, M. A. & Bates, P. A. Transmission of cutaneous leishmaniasis by sand flies is enhanced by regurgitation of fPPG. Nature 430, 463–467 (2004).
Beattie, L. et al. Dynamic imaging of experimental Leishmania donovani-induced hepatic granulomas detects Kupffer cell-restricted antigen presentation to antigen-specific CD8 T cells. PLoS Pathog. 6, e1000805 (2010).
De Trez, C. et al. iNOS-producing inflammatory dendritic cells constitute the major infected cell type during the chronic Leishmania major infection phase of C57BL/6 resistant mice. PLoS Pathog. 5, e1000494 (2009).
Ravichandran, K. S. & Lorenz, U. Engulfment of apoptotic cells: signals for a good meal. Nature Rev. Immunol. 7, 964–974 (2007).
Laskay, T., van Zandbergen, G. & Solbach, W. Neutrophil granulocytes — Trojan horses for Leishmania major and other intracellular microbes? Trends Microbiol. 11, 210–214 (2003).
Peters, N. C. et al. In vivo imaging reveals an essential role for neutrophils in leishmaniasis transmitted by sand flies. Science 321, 970–974 (2008).
Thalhofer, C. J., Chen, Y., Sudan, B., Love-Homan, L. & Wilson, M. E. Leukocytes infiltrating the skin and draining lymph nodes in response to the protozoan Leishmania infantum chagasi. Infect. Immun. 79, 108–117 (2010).
Bianchi, M. E. DAMPs, PAMPs and alarmins: all we need to know about danger. J. Leukoc. Biol. 81, 1–5 (2007).
Haraldsen, G., Balogh, J., Pollheimer, J., Sponheim, J. & Kuchler, A. M. Interleukin-33 — cytokine of dual function or novel alarmin? Trends Immunol. 30, 227–233 (2009).
Oghumu, S., Lezama-Davila, C. M., Isaac-Marquez, A. P. & Satoskar, A. R. Role of chemokines in regulation of immunity against leishmaniasis. Exp. Parasitol. 126, 389–396 (2010).
Lopez Kostka, S. et al. IL-17 promotes progression of cutaneous leishmaniasis in susceptible mice. J. Immunol. 182, 3039–3046 (2009).
Xin, L. et al. Type I IFN receptor regulates neutrophil functions and innate immunity to Leishmania parasites. J. Immunol. 184, 7047–7056 (2010).
Depaquit, J., Grandadam, M., Fouque, F., Andry, P. E. & Peyrefitte, C. Arthropod-borne viruses transmitted by Phlebotomine sandflies in Europe: a review. Euro. Surveill. 15, 19507 (2010).
Hillesland, H. et al. Identification of aerobic gut bacteria from the kala azar vector, Phlebotomus argentipes: a platform for potential paratransgenic manipulation of sand flies. Am. J. Trop. Med. Hyg. 79, 881–886 (2008).
Novais, F. O. et al. Neutrophils and macrophages cooperate in host resistance against Leishmania braziliensis infection. J. Immunol. 183, 8088–8098 (2009).
Ribeiro-Gomes, F. L. et al. Macrophage interactions with neutrophils regulate Leishmania major infection. J. Immunol. 172, 4454–4462 (2004).
Gabriel, C., McMaster, W. R., Girard, D. & Descoteaux, A. Leishmania donovani promastigotes evade the antimicrobial activity of neutrophil extracellular traps. J. Immunol. 185, 4319–4327 (2010).
Guimaraes-Costa, A. B. et al. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc. Natl Acad. Sci. USA 106, 6748–6753 (2009).
Ng, L. G. et al. Migratory dermal dendritic cells act as rapid sensors of protozoan parasites. PLoS Pathog. 4, e1000222 (2008).
Rittig, M. G. et al. Coiling phagocytosis of trypanosomatids and fungal cells. Infect. Immun. 66, 4331–4339 (1998).
Charmoy, M. et al. Neutrophil-derived CCL3 is essential for the rapid recruitment of dendritic cells to the site of Leishmania major inoculation in resistant mice. PLoS Pathog. 6, e1000755 (2010).
Leon, B., Lopez-Bravo, M. & Ardavin, C. Monocyte-derived dendritic cells formed at the infection site control the induction of protective T helper 1 responses against Leishmania. Immunity 26, 519–531 (2007).
Horst, A. K. et al. CEACAM1+ myeloid cells control angiogenesis in inflammation. Blood 113, 6726–6736 (2009).
Sato, N. et al. CC chemokine receptor (CCR)2 is required for Langerhans cell migration and localization of T helper cell type 1 (Th1)-inducing dendritic cells: absence of CCR2 shifts the Leishmania major–resistant phenotype to a susceptible state dominated by Th2 cytokines, B cell outgrowth, and sustained neutrophilic inflammation. J. Exp. Med. 192, 205–218 (2000).
Carvalho, L. P., Pearce, E. J. & Scott, P. Functional dichotomy of dendritic cells following interaction with Leishmania braziliensis: infected cells produce high levels of TNFα, whereas bystander dendritic cells are activated to promote T cell responses. J. Immunol. 181, 6473–6480 (2008).
Maroof, A., Beattie, L., Kirby, A., Coles, M. & Kaye, P. M. Dendritic cells matured by inflammation induce CD86-dependent priming of naive CD8+ T cells in the absence of their cognate peptide antigen. J. Immunol. 183, 7095–7103 (2009).
Engwerda, C. R., Ato, M. & Kaye, P. M. Macrophages, pathology and parasite persistence in experimental visceral leishmaniasis. Trends Parasitol. 20, 524–530 (2004).
Bogdan, C. et al. Fibroblasts as host cells in latent leishmaniosis. J. Exp. Med. 191, 2121–2130 (2000).
Svensson, M., Maroof, A., Ato, M. & Kaye, P. M. Stromal cells direct local differentiation of regulatory dendritic cells. Immunity 21, 805–816 (2004).
Gorgani, N. N., Ma, Y. & Clark, H. F. Gene signatures reflect the marked heterogeneity of tissue-resident macrophages. Immunol. Cell Biol. 86, 246–254 (2008).
Gueirard, P., Laplante, A., Rondeau, C., Milon, G. & Desjardins, M. Trafficking of Leishmania donovani promastigotes in non-lytic compartments in neutrophils enables the subsequent transfer of parasites to macrophages. Cell. Microbiol. 10, 100–111 (2008).
Desjardins, M. & Descoteaux, A. Inhibition of phagolysosomal biogenesis by the Leishmania lipophosphoglycan. J. Exp. Med. 185, 2061–2068 (1997).
Mollinedo, F., Janssen, H., de la Iglesia-Vicente, J., Villa-Pulgarin, J. A. & Calafat, J. Selective fusion of azurophilic granules with Leishmania-containing phagosomes in human neutrophils. J. Biol. Chem. 285, 34528–34536 (2010).
Ndjamen, B., Kang, B. H., Hatsuzawa, K. & Kima, P. E. Leishmania parasitophorous vacuoles interact continuously with the host cell's endoplasmic reticulum; parasitophorous vacuoles are hybrid compartments. Cell. Microbiol. 12, 1480–1494 (2010).
Lippuner, C. et al. Real-time imaging of Leishmania mexicana-infected early phagosomes: a study using primary macrophages generated from green fluorescent protein-Rab5 transgenic mice. FASEB J. 23, 483–491 (2009).
Vinet, A. F., Fukuda, M., Turco, S. J. & Descoteaux, A. The Leishmania donovani lipophosphoglycan excludes the vesicular proton-ATPase from phagosomes by impairing the recruitment of synaptotagmin V. PLoS Pathog. 5, e1000628 (2009).
Spath, G. F., Schlesinger, P., Schreiber, R. & Beverley, S. M. A novel role for Stat1 in phagosome acidification and natural host resistance to intracellular infection by Leishmania major. PLoS Pathog. 5, e1000381 (2009).
Alcolea, P. J. et al. Temperature increase prevails over acidification in gene expression modulation of amastigote differentiation in Leishmania infantum. BMC Genomics 11, 31 (2010).
Wilson, J. et al. Control of parasitophorous vacuole expansion by LYST/Beige restricts the intracellular growth of Leishmania amazonensis. PLoS Pathog. 4, e1000179 (2008).
Huynh, C. & Andrews, N. W. Iron acquisition within host cells and the pathogenicity of Leishmania. Cell. Microbiol. 10, 293–300 (2008).
Carvalho, S. et al. Heme as a source of iron to Leishmania infantum amastigotes. Acta Trop. 109, 131–135 (2009).
Blackwell, J. M. et al. SLC11A1 (formerly NRAMP1) and disease resistance. Cell. Microbiol. 3, 773–784 (2001).
Jacques, I., Andrews, N. W. & Huynh, C. Functional characterization of LIT1, the Leishmania amazonensis ferrous iron transporter. Mol. Biochem. Parasitol. 170, 28–36 (2010).
Das, N. K., Biswas, S., Solanki, S. & Mukhopadhyay, C. K. Leishmania donovani depletes labile iron pool to exploit iron uptake capacity of macrophage for its intracellular growth. Cell. Microbiol. 11, 83–94 (2009).
Gregory, D. J. & Olivier, M. Subversion of host cell signalling by the protozoan parasite Leishmania. Parasitology 130 Suppl S1, S27–S35 (2005).
Joshi, P. B., Kelly, B. L., Kamhawi, S., Sacks, D. L. & McMaster, W. R. Targeted gene deletion in Leishmania major identifies leishmanolysin (GP63) as a virulence factor. Mol. Biochem. Parasitol. 120, 33–40 (2002).
Victoir, K. et al. Complexity of the major surface protease (msp) gene organization in Leishmania (Viannia) braziliensis: evolutionary and functional implications. Parasitology 131, 207–214 (2005).
Hsiao, C. H., Yao, C., Storlie, P., Donelson, J. E. & Wilson, M. E. The major surface protease (MSP or GP63) in the intracellular amastigote stage of Leishmania chagasi. Mol. Biochem. Parasitol. 157, 148–159 (2008).
Gomez, M. A. et al. Leishmania GP63 alters host signaling through cleavage-activated protein tyrosine phosphatases. Sci. Signal. 2, ra58 (2009).
Halle, M. et al. The Leishmania surface protease GP63 cleaves multiple intracellular proteins and actively participates in p38 mitogen-activated protein kinase inactivation. J. Biol. Chem. 284, 6893–6908 (2009).
Yao, C., Donelson, J. E. & Wilson, M. E. Internal and surface-localized major surface proteases of Leishmania spp. and their differential release from promastigotes. Eukaryot. Cell 6, 1905–1912 (2007).
Winnen, B. et al. Hierarchical effector protein transport by the Salmonella Typhimurium SPI-1 type III secretion system. PLoS ONE 3, e2178 (2008).
Barclay, A. N. & Brown, M. H. The SIRP family of receptors and immune regulation. Nature Rev. Immunol. 6, 457–464 (2006).
Takizawa, H. & Manz, M. G. Macrophage tolerance: CD47–SIRP-α–mediated signals matter. Nature Immunol. 8, 1287–1289 (2007).
Beattie, L. et al. Leishmania donovani-induced expression of signal regulatory protein alpha on Kupffer cells enhances hepatic invariant NKT-cell activation. Eur. J. Immunol. 40, 117–123 (2010).
Pathak, M. K. & Yi, T. Sodium stibogluconate is a potent inhibitor of protein tyrosine phosphatases and augments cytokine responses in hemopoietic cell lines. J. Immunol. 167, 3391–3397 (2001).
Forget, G. et al. Role of host phosphotyrosine phosphatase SHP-1 in the development of murine leishmaniasis. Eur. J. Immunol. 31, 3185–3196 (2001).
Spath, G. F., McDowell, M. A. & Beverley, S. M. Leishmania major intracellular survival is not altered in SHP-1 deficient mev or CD45-/- mice. Exp. Parasitol. 120, 275–279 (2008).
Olivier, M., Baimbridge, K. G. & Reiner, N. E. Stimulus-response coupling in monocytes infected with Leishmania. Attenuation of calcium transients is related to defective agonist-induced accumulation of inositol phosphates. J. Immunol. 148, 1188–1196 (1992).
Alvarez-Rueda, N., Biron, M. & Le Pape, P. Infectivity of Leishmania mexicana is associated with differential expression of protein kinase C-like triggered during a cell–cell contact. PLoS ONE 4, e7581 (2009).
Ben-Othman, R., Dellagi, K. & Guizani-Tabbane, L. Leishmania major parasites induced macrophage tolerance: implication of MAPK and NF-κB pathways. Mol. Immunol. 46, 3438–3444 (2009).
Ben-Othman, R., Guizani-Tabbane, L. & Dellagi, K. Leishmania initially activates but subsequently down-regulates intracellular mitogen-activated protein kinases and nuclear factor-κB signaling in macrophages. Mol. Immunol. 45, 3222–3229 (2008).
Calegari-Silva, T. C. et al. NF-κB-mediated repression of iNOS expression in Leishmania amazonensis macrophage infection. Immunol. Lett. 127, 19–26 (2009).
Rodriguez, N. E., Gaur, U. & Wilson, M. E. Role of caveolae in Leishmania chagasi phagocytosis and intracellular survival in macrophages. Cell. Microbiol. 8, 1106–1120 (2006).
Osorio y Fortea, J. et al. Transcriptional signatures of BALB/c mouse macrophages housing multiplying Leishmania amazonensis amastigotes. BMC Genomics 10, 119 (2009).
Phillips, R. et al. Innate killing of Leishmania donovani by macrophages of the splenic marginal zone requires IRF-7. PLoS Pathog. 6, e1000813 (2010).
Chakraborty, D. et al. Leishmania donovani affects antigen presentation of macrophage by disrupting lipid rafts. J. Immunol. 175, 3214–3224 (2005).
Rub, A. et al. Cholesterol depletion associated with Leishmania major infection alters macrophage CD40 signalosome composition and effector function. Nature Immunol. 10, 273–280 (2009).
Srivastava, N., Sudan, R. & Saha, B. CD40-modulated dual-specificity phosphatases MAPK phosphatase (MKP)-1 and MKP-3 reciprocally regulate Leishmania major infection. J. Immunol. 186, 5863–5872 (2011).
Winberg, M. E. et al. Leishmania donovani lipophosphoglycan inhibits phagosomal maturation via action on membrane rafts. Microbes Infect. 11, 215–222 (2009).
Garg, R., Lodge, R., Descoteaux, A. & Tremblay, M. J. Leishmania infantum promastigotes reduce entry of HIV-1 into macrophages through a lipophosphoglycan-mediated disruption of lipid rafts. J. Infect. Dis. 197, 1701–1708 (2008).
Silverman, J. M. & Reiner, N. E. Exosomes and other microvesicles in infection biology: organelles with unanticipated phenotypes. Cell. Microbiol. 13, 1–9 (2010).
Silverman, J. M. et al. An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages. J. Cell Sci. 123, 842–852 (2010).
Silverman, J. M. et al. Leishmania exosomes modulate innate and adaptive immune responses through effects on monocytes and dendritic cells. J. Immunol. 185, 5011–5022 (2010).
Pereira, R. M. et al. Novel role for the double-stranded RNA-activated protein kinase PKR: modulation of macrophage infection by the protozoan parasite Leishmania. FASEB J. 24, 617–626 (2010).
Guilbride, L., Myler, P. J. & Stuart, K. Distribution and sequence divergence of LRV1 viruses among different Leishmania species. Mol. Biochem. Parasitol. 54, 101–104 (1992).
Ives, A. et al. Leishmania RNA virus controls the severity of mucocutaneous leishmaniasis. Science 331, 775–778 (2011).
Stern, J. J., Oca, M. J., Rubin, B. Y., Anderson, S. L. & Murray, H. W. Role of L3T4+ and LyT-2+ cells in experimental visceral leishmaniasis. J. Immunol. 140, 3971–3977 (1988).
Muller, I., Kropf, P., Etges, R. J. & Louis, J. A. Gamma interferon response in secondary Leishmania major infection: role of CD8+ T cells. Infect. Immun. 61, 3730–3738 (1993).
Belkaid, Y. et al. CD8+ T cells are required for primary immunity in C57BL/6 mice following low-dose, intradermal challenge with Leishmania major. J. Immunol. 168, 3992–4000 (2002).
Uzonna, J. E., Joyce, K. L. & Scott, P. Low dose Leishmania major promotes a transient T helper cell type 2 response that is down-regulated by interferon-γ-producing CD8+ T cells. J. Exp. Med. 199, 1559–1566 (2004).
Faria, D. R. et al. Recruitment of CD8+ T cells expressing granzyme A is associated with lesion progression in human cutaneous leishmaniasis. Parasite Immunol. 31, 432–439 (2009).
Joshi, T., Rodriguez, S., Perovic, V., Cockburn, I. A. & Stager, S. B7-H1 blockade increases survival of dysfunctional CD8+ T cells and confers protection against Leishmania donovani infections. PLoS Pathog. 5, e1000431 (2009).
Brewig, N. et al. Priming of CD8+ and CD4+ T cells in experimental leishmaniasis is initiated by different dendritic cell subtypes. J. Immunol. 182, 774–783 (2009).
Polley, R., Sanos, S. L., Prickett, S., Haque, A. & Kaye, P. M. Chronic Leishmania donovani infection promotes bystander CD8+-T-cell expansion and heterologous immunity. Infect. Immun. 73, 7996–8001 (2005).
Belkaid, Y. et al. The role of interleukin (IL)-10 in the persistence of Leishmania major in the skin after healing and the therapeutic potential of anti-IL-10 receptor antibody for sterile cure. J. Exp. Med. 194, 1497–1506 (2001).
Belkaid, Y., Piccirillo, C. A., Mendez, S., Shevach, E. M. & Sacks, D. L. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 420, 502–507 (2002).
Anderson, C. F., Oukka, M., Kuchroo, V. J. & Sacks, D. CD4+CD25−Foxp3− Th1 cells are the source of IL-10–mediated immune suppression in chronic cutaneous leishmaniasis. J. Exp. Med. 204, 285–297 (2007).
Nylen, S. et al. Splenic accumulation of IL-10 mRNA in T cells distinct from CD4+CD25+ (Foxp3) regulatory T cells in human visceral leishmaniasis. J. Exp. Med. 204, 805–817 (2007).
Stager, S. et al. Distinct roles for IL-6 and IL-12p40 in mediating protection against Leishmania donovani and the expansion of IL-10+ CD4+ T cells. Eur. J. Immunol. 36, 1764–1771 (2006).
Ronet, C. et al. Regulatory B cells shape the development of Th2 immune responses in BALB/c mice infected with Leishmania major through IL-10 production. J. Immunol. 184, 886–894 (2010).
Maroof, A. et al. Posttranscriptional regulation of IL-10 gene expression allows natural killer cells to express immunoregulatory function. Immunity 29, 295–305 (2008).
Miles, S. A., Conrad, S. M., Alves, R. G., Jeronimo, S. M. & Mosser, D. M. A role for IgG immune complexes during infection with the intracellular pathogen Leishmania. J. Exp. Med. 201, 747–754 (2005).
Charmoy, M. et al. Leishmania major induces distinct neutrophil phenotypes in mice that are resistant or susceptible to infection. J. Leukoc. Biol. 82, 288–299 (2007).
Groux, H. et al. A transgenic model to analyze the immunoregulatory role of IL-10 secreted by antigen-presenting cells. J. Immunol. 162, 1723–1729 (1999).
Ranatunga, D. et al. A human IL10 BAC transgene reveals tissue-specific control of IL-10 expression and alters disease outcome. Proc. Natl Acad. Sci. USA 106, 17123–17128 (2009).
Martin, S., Pahari, S., Sudan, R. & Saha, B. CD40 signaling in CD8+CD40+ T cells turns on contra-T regulatory cell functions. J. Immunol. 184, 5510–5518 (2010).
Dalton, J. E. et al. Inhibition of receptor tyrosine kinases restores immunocompetence and improves immune-dependent chemotherapy against experimental leishmaniasis in mice. J. Clin. Invest. 120, 1204–1216 (2010).
Ato, M., Stager, S., Engwerda, C. R. & Kaye, P. M. Defective CCR7 expression on dendritic cells contributes to the development of visceral leishmaniasis. Nature Immunol. 3, 1185–1191 (2002).
Machado, P. R. et al. Oral pentoxifylline combined with pentavalent antimony: a randomized trial for mucosal leishmaniasis. Clin. Infect. Dis. 44, 788–793 (2007).
Filipe-Santos, O. et al. A dynamic map of antigen recognition by CD4 T cells at the site of Leishmania major infection. Cell Host Microbe 6, 23–33 (2009).
Reckling, S. et al. Proapoptotic Bcl-2 family member Bim promotes persistent infection and limits protective immunity . Infect. Immun. 76, 1179–1185 (2008).
Zaph, C., Uzonna, J., Beverley, S. M. & Scott, P. Central memory T cells mediate long-term immunity to Leishmania major in the absence of persistent parasites. Nature Med. 10, 1104–1110 (2004).
Colpitts, S. L., Dalton, N. M. & Scott, P. IL-7 receptor expression provides the potential for long-term survival of both CD62Lhigh central memory T cells and Th1 effector cells during Leishmania major infection. J. Immunol. 182, 5702–5711 (2009).
Darrah, P. A. et al. Multifunctional TH1 cells define a correlate of vaccine-mediated protection against Leishmania major. Nature Med. 13, 843–850 (2007).
Darrah, P. A. et al. IL-10 production differentially influences the magnitude, quality, and protective capacity of Th1 responses depending on the vaccine platform. J. Exp. Med. 207, 1421–1433 (2010).
Working Group on Research Priorities for Development of Leishmaniasis Vaccines et al. Vaccines for the leishmaniases: proposals for a research agenda. PLoS Negl. Trop. Dis. 5, e943 (2011).
Mosser, D. M. & Edwards, J. P. Exploring the full spectrum of macrophage activation. Nature Rev. Immunol. 8, 958–969 (2008).
Geissmann, F. et al. Development of monocytes, macrophages, and dendritic cells. Science 327, 656–661 (2010).
Sacks, D. L. Leishmania-sand fly interactions controlling species-specific vector competence. Cell. Microbiol. 3, 189–196 (2001).
Beverley, S. M. & Turco, S. J. Lipophosphoglycan (LPG) and the identification of virulence genes in the protozoan parasite Leishmania. Trends Microbiol. 6, 35–40 (1998).
Korner, U., Fuss, V., Steigerwald, J. & Moll, H. Biogenesis of Leishmania major-harboring vacuoles in murine dendritic cells. Infect. Immun. 74, 1305–1312 (2006).
World Health Organization. Control of the leishmaniasis: report of a meeting of the WHO Expert Committee on the Control of Leishmaniases (WHO, Geneva, 2010).
Acknowledgements
Work in the P.K. laboratory is funded by grants from the UK Medical Research Council and the Wellcome Trust. Work in the P.S. laboratory is funded by grants from the National Institute of Allergy and Infectious Diseases, US National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Related links
Glossary
- Two-photon intravital imaging
-
A fluorescent-laser-based microscopy technique that allows real-time imaging of cells that are deep within the tissues of live animals.
- Chemokines
-
A family of small (∼810 kDa) chemotactic cytokines that regulate cell migration and function. Different chemokine families are defined by the presence of specific motifs known as C, CC, CXC and CX3C.
- Granules
-
Cytosolic particles within neutrophils that are referred to as primary, tertiary or specific, based on their staining characteristics and their content. Primary granules (also called azurophilic granules) contain the enzyme myeloperoxidase, which is responsible for the green colour of pus.
- RAB GTPases
-
A large family of monomeric guanine nucleotide binding proteins that act as important regulators of vesicle transport and docking in eukaryotic cells.
- Fc receptors
-
Cell surface receptors that recognize the Fc tails of antibody molecules. The binding of particles that are coated with antibody by the Fc receptors on phagocytic cells triggers internalisation and potent antimicrobial effector responses.
- Lipid microdomains
-
Regions of a biological membrane that are enriched in a particular protein or lipid and can vary in size, composition and function. For example, lipid rafts are highly dynamic microdomains, 10–200 nm in size, that are enriched in cholesterol and sphingolipids and serve to compartmentalize different membrane functions.
- Signalosome
-
A high-molecular-mass cytosolic or membrane-bound complex that contains many of the individual proteins that are associated with a signalling pathway, often together with various scaffold proteins.
- Concomitant immunity
-
A situation in which immunological resistance to reinfection co-exists at the same time as persistence of the original infection.
Rights and permissions
About this article
Cite this article
Kaye, P., Scott, P. Leishmaniasis: complexity at the host–pathogen interface. Nat Rev Microbiol 9, 604–615 (2011). https://doi.org/10.1038/nrmicro2608
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrmicro2608
This article is cited by
-
Anti-Leishmania major activity of Calotropis procera extract by increasing ROS production and upregulating TNF-α, IFN-γ and iNOS mRNA expression under in vitro conditions
Tropical Medicine and Health (2024)
-
Antigen recognition reinforces regulatory T cell mediated Leishmania major persistence
Nature Communications (2023)
-
Centrin-deficient Leishmania mexicana confers protection against Old World visceral leishmaniasis
npj Vaccines (2022)
-
Revisiting nature: a review of iridoids as a potential antileishmanial class
Phytochemistry Reviews (2022)
-
Epigenetic paradigms/exemplars of the macrophage: inflammasome axis in Leishmaniasis
Molecular and Cellular Biochemistry (2022)
