
Key Points
-
Recent improvements in mRNA vaccines act to increase protein translation, modulate innate and adaptive immunogenicity and improve delivery.
-
mRNA vaccines have elicited potent immunity against infectious disease targets in animal models of influenza virus, Zika virus, rabies virus and others, especially in recent years, using lipid-encapsulated or naked forms of sequence-optimized mRNA.
-
Diverse approaches to mRNA cancer vaccines, including dendritic cell vaccines and various types of directly injectable mRNA, have been employed in numerous cancer clinical trials, with some promising results showing antigen-specific T cell responses and prolonged disease-free survival in some cases.
-
Therapeutic considerations and challenges include scaling up good manufacturing practice (GMP) production, establishing regulations, further documenting safety and increasing efficacy.
-
Important future directions of research will be to compare and elucidate the immune pathways activated by various mRNA vaccine platforms, to improve current approaches based on these mechanisms and to initiate new clinical trials against additional disease targets.
Abstract
mRNA vaccines represent a promising alternative to conventional vaccine approaches because of their high potency, capacity for rapid development and potential for low-cost manufacture and safe administration. However, their application has until recently been restricted by the instability and inefficient in vivo delivery of mRNA. Recent technological advances have now largely overcome these issues, and multiple mRNA vaccine platforms against infectious diseases and several types of cancer have demonstrated encouraging results in both animal models and humans. This Review provides a detailed overview of mRNA vaccines and considers future directions and challenges in advancing this promising vaccine platform to widespread therapeutic use.
Similar content being viewed by others
Main
Vaccines prevent many millions of illnesses and save numerous lives every year1. As a result of widespread vaccine use, the smallpox virus has been completely eradicated and the incidence of polio, measles and other childhood diseases has been drastically reduced around the world2. Conventional vaccine approaches, such as live attenuated and inactivated pathogens and subunit vaccines, provide durable protection against a variety of dangerous diseases3. Despite this success, there remain major hurdles to vaccine development against a variety of infectious pathogens, especially those better able to evade the adaptive immune response4. Moreover, for most emerging virus vaccines, the main obstacle is not the effectiveness of conventional approaches but the need for more rapid development and large-scale deployment. Finally, conventional vaccine approaches may not be applicable to non-infectious diseases, such as cancer. The development of more potent and versatile vaccine platforms is therefore urgently needed.
Nucleic acid therapeutics have emerged as promising alternatives to conventional vaccine approaches. The first report of the successful use of in vitro transcribed (IVT) mRNA in animals was published in 1990, when reporter gene mRNAs were injected into mice and protein production was detected5. A subsequent study in 1992 demonstrated that administration of vasopressin-encoding mRNA in the hypothalamus could elicit a physiological response in rats6. However, these early promising results did not lead to substantial investment in developing mRNA therapeutics, largely owing to concerns associated with mRNA instability, high innate immunogenicity and inefficient in vivo delivery. Instead, the field pursued DNA-based and protein-based therapeutic approaches7,8.
Over the past decade, major technological innovation and research investment have enabled mRNA to become a promising therapeutic tool in the fields of vaccine development and protein replacement therapy. The use of mRNA has several beneficial features over subunit, killed and live attenuated virus, as well as DNA-based vaccines. First, safety: as mRNA is a non-infectious, non-integrating platform, there is no potential risk of infection or insertional mutagenesis. Additionally, mRNA is degraded by normal cellular processes, and its in vivo half-life can be regulated through the use of various modifications and delivery methods9,10,11,12. The inherent immunogenicity of the mRNA can be down-modulated to further increase the safety profile9,12,13. Second, efficacy: various modifications make mRNA more stable and highly translatable9,12,13. Efficient in vivo delivery can be achieved by formulating mRNA into carrier molecules, allowing rapid uptake and expression in the cytoplasm (reviewed in Refs 10,11). mRNA is the minimal genetic vector; therefore, anti-vector immunity is avoided, and mRNA vaccines can be administered repeatedly. Third, production: mRNA vaccines have the potential for rapid, inexpensive and scalable manufacturing, mainly owing to the high yields of in vitro transcription reactions.
The mRNA vaccine field is developing extremely rapidly; a large body of preclinical data has accumulated over the past several years, and multiple human clinical trials have been initiated. In this Review, we discuss current mRNA vaccine approaches, summarize the latest findings, highlight challenges and recent successes, and offer perspectives on the future of mRNA vaccines. The data suggest that mRNA vaccines have the potential to solve many of the challenges in vaccine development for both infectious diseases and cancer.
Basic mRNA vaccine pharmacology
mRNA is the intermediate step between the translation of protein-encoding DNA and the production of proteins by ribosomes in the cytoplasm. Two major types of RNA are currently studied as vaccines: non-replicating mRNA and virally derived, self-amplifying RNA. Conventional mRNA-based vaccines encode the antigen of interest and contain 5′ and 3′ untranslated regions (UTRs), whereas self-amplifying RNAs encode not only the antigen but also the viral replication machinery that enables intracellular RNA amplification and abundant protein expression.
The construction of optimally translated IVT mRNA suitable for therapeutic use has been reviewed previously14,15. Briefly, IVT mRNA is produced from a linear DNA template using a T7, a T3 or an Sp6 phage RNA polymerase16. The resulting product should optimally contain an open reading frame that encodes the protein of interest, flanking UTRs, a 5′ cap and a poly(A) tail. The mRNA is thus engineered to resemble fully processed mature mRNA molecules as they occur naturally in the cytoplasm of eukaryotic cells.
Complexing of mRNA for in vivo delivery has also been recently detailed10,11. Naked mRNA is quickly degraded by extracellular RNases17 and is not internalized efficiently. Thus, a great variety of in vitro and in vivo transfection reagents have been developed that facilitate cellular uptake of mRNA and protect it from degradation. Once the mRNA transits to the cytosol, the cellular translation machinery produces protein that undergoes post-translational modifications, resulting in a properly folded, fully functional protein. This feature of mRNA pharmacology is particularly advantageous for vaccines and protein replacement therapies that require cytosolic or transmembrane proteins to be delivered to the correct cellular compartments for proper presentation or function. IVT mRNA is finally degraded by normal physiological processes, thus reducing the risk of metabolite toxicity.
Recent advances in mRNA vaccine technology
Various mRNA vaccine platforms have been developed in recent years and validated in studies of immunogenicity and efficacy18,19,20. Engineering of the RNA sequence has rendered synthetic mRNA more translatable than ever before. Highly efficient and non-toxic RNA carriers have been developed that in some cases21,22 allow prolonged antigen expression in vivo (Table 1). Some vaccine formulations contain novel adjuvants, while others elicit potent responses in the absence of known adjuvants. The following section summarizes the key advances in these areas of mRNA engineering and their impact on vaccine efficacy.
Optimization of mRNA translation and stability
This topic has been extensively discussed in previous reviews14,15; thus, we briefly summarize the key findings (Box 1). The 5′ and 3′ UTR elements flanking the coding sequence profoundly influence the stability and translation of mRNA, both of which are critical concerns for vaccines. These regulatory sequences can be derived from viral or eukaryotic genes and greatly increase the half-life and expression of therapeutic mRNAs23,24. A 5′ cap structure is required for efficient protein production from mRNA25. Various versions of 5′ caps can be added during or after the transcription reaction using a vaccinia virus capping enzyme26 or by incorporating synthetic cap or anti-reverse cap analogues27,28. The poly(A) tail also plays an important regulatory role in mRNA translation and stability25; thus, an optimal length of poly(A)24 must be added to mRNA either directly from the encoding DNA template or by using poly(A) polymerase. The codon usage additionally has an impact on protein translation. Replacing rare codons with frequently used synonymous codons that have abundant cognate tRNA in the cytosol is a common practice to increase protein production from mRNA29, although the accuracy of this model has been questioned30. Enrichment of G:C content constitutes another form of sequence optimization that has been shown to increase steady-state mRNA levels in vitro31 and protein expression in vivo12.
Although protein expression may be positively modulated by altering the codon composition or by introducing modified nucleosides (discussed below), it is also possible that these forms of sequence engineering could affect mRNA secondary structure32, the kinetics and accuracy of translation and simultaneous protein folding33,34, and the expression of cryptic T cell epitopes present in alternative reading frames30. All these factors could potentially influence the magnitude or specificity of the immune response.
Modulation of immunogenicity
Exogenous mRNA is inherently immunostimulatory, as it is recognized by a variety of cell surface, endosomal and cytosolic innate immune receptors (Fig. 1) (reviewed in Ref. 35). Depending on the therapeutic application, this feature of mRNA could be beneficial or detrimental. It is potentially advantageous for vaccination because in some cases it may provide adjuvant activity to drive dendritic cell (DC) maturation and thus elicit robust T and B cell immune responses. However, innate immune sensing of mRNA has also been associated with the inhibition of antigen expression and may negatively affect the immune response9,13. Although the paradoxical effects of innate immune sensing on different formats of mRNA vaccines are incompletely understood, some progress has been made in recent years in elucidating these phenomena.

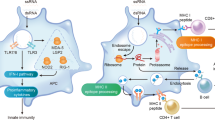
Innate immune sensing of two types of mRNA vaccine by a dendritic cell (DC), with RNA sensors shown in yellow, antigen in red, DC maturation factors in green, and peptide−major histocompatibility complex (MHC) complexes in light blue and red; an example lipid nanoparticle carrier is shown at the top right. A non-exhaustive list of the major known RNA sensors that contribute to the recognition of double-stranded and unmodified single-stranded RNAs is shown. Unmodified, unpurified (part a) and nucleoside-modified, fast protein liquid chromatography (FPLC)-purified (part b) mRNAs were selected for illustration of two formats of mRNA vaccines where known forms of mRNA sensing are present and absent, respectively. The dashed arrow represents reduced antigen expression. Ag, antigen; PKR, interferon-induced, double-stranded RNA-activated protein kinase; MDA5, interferon-induced helicase C domain-containing protein 1 (also known as IFIH1); IFN, interferon; m1Ψ, 1-methylpseudouridine; OAS, 2′-5′-oligoadenylate synthetase; TLR, Toll-like receptor.
Studies over the past decade have shown that the immunostimulatory profile of mRNA can be shaped by the purification of IVT mRNA and the introduction of modified nucleosides as well as by complexing the mRNA with various carrier molecules9,13,36,37. Enzymatically synthesized mRNA preparations contain double-stranded RNA (dsRNA) contaminants as aberrant products of the IVT reaction13. As a mimic of viral genomes and replication intermediates, dsRNA is a potent pathogen-associated molecular pattern (PAMP) that is sensed by pattern recognition receptors in multiple cellular compartments (Fig. 1). Recognition of IVT mRNA contaminated with dsRNA results in robust type I interferon production13, which upregulates the expression and activation of protein kinase R (PKR; also known as EIF2AK2) and 2′-5′-oligoadenylate synthetase (OAS), leading to the inhibition of translation38 and the degradation of cellular mRNA and ribosomal RNA39, respectively. Karikó and colleagues13 have demonstrated that contaminating dsRNA can be efficiently removed from IVT mRNA by chromatographic methods such as reverse-phase fast protein liquid chromatography (FPLC) or high-performance liquid chromatography (HPLC). Strikingly, purification by FPLC has been shown to increase protein production from IVT mRNA by up to 1,000-fold in primary human DCs13. Thus, appropriate purification of IVT mRNA seems to be critical for maximizing protein (immunogen) production in DCs and for avoiding unwanted innate immune activation.
Besides dsRNA contaminants, single-stranded mRNA molecules are themselves a PAMP when delivered to cells exogenously. Single-stranded oligoribonucleotides and their degradative products are detected by the endosomal sensors Toll-like receptor 7 (TLR7) and TLR8 (Refs 40,41), resulting in type I interferon production42. Crucially, it was discovered that the incorporation of naturally occurring chemically modified nucleosides, including but not limited to pseudouridine9,43,44 and 1-methylpseudouridine45, prevents activation of TLR7, TLR8 and other innate immune sensors46,47, thus reducing type I interferon signalling48. Nucleoside modification also partially suppresses the recognition of dsRNA species46,47,48. As a result, Karikó and others have shown that nucleoside-modified mRNA is translated more efficiently than unmodified mRNA in vitro9, particularly in primary DCs, and in vivo in mice45. Notably, the highest level of protein production in DCs was observed when mRNA was both FPLC-purified and nucleoside-modified13. These advances in understanding the sources of innate immune sensing and how to avoid their adverse effects have substantially contributed to the current interest in mRNA-based vaccines and protein replacement therapies.
In contrast to the findings described above, a study by Thess and colleagues found that sequence-optimized, HPLC-purified, unmodified mRNA produced higher levels of protein in HeLa cells and in mice than its nucleoside-modified counterpart12. Additionally, Kauffman and co-workers demonstrated that unmodified, non-HPLC-purified mRNA yielded more robust protein production in HeLa cells than nucleoside-modified mRNA, and resulted in similar levels of protein production in mice49. Although not fully clear, the discrepancies between the findings of Karikó9,13 and these authors12,49 may have arisen from variations in RNA sequence optimization, the stringency of mRNA purification to remove dsRNA contaminants and the level of innate immune sensing in the targeted cell types.
The immunostimulatory properties of mRNA can conversely be increased by the inclusion of an adjuvant to increase the potency of some mRNA vaccine formats. These include traditional adjuvants as well as novel approaches that take advantage of the intrinsic immunogenicity of mRNA or its ability to encode immune-modulatory proteins. Self-replicating RNA vaccines have displayed increased immunogenicity and effectiveness after formulating the RNA in a cationic nanoemulsion based on the licensed MF59 (Novartis) adjuvant50. Another effective adjuvant strategy is TriMix, a combination of mRNAs encoding three immune activator proteins: CD70, CD40 ligand (CD40L) and constitutively active TLR4. TriMix mRNA augmented the immunogenicity of naked, unmodified, unpurified mRNA in multiple cancer vaccine studies and was particularly associated with increased DC maturation and cytotoxic T lymphocyte (CTL) responses (reviewed in Ref. 51). The type of mRNA carrier and the size of the mRNA–carrier complex have also been shown to modulate the cytokine profile induced by mRNA delivery. For example, the RNActive (CureVac AG) vaccine platform52,53 depends on its carrier to provide adjuvant activity. In this case, the antigen is expressed from a naked, unmodified, sequence-optimized mRNA, while the adjuvant activity is provided by co-delivered RNA complexed with protamine (a polycationic peptide), which acts via TLR7 signalling52,54. This vaccine format has elicited favourable immune responses in multiple preclinical animal studies for vaccination against cancer and infectious diseases18,36,55,56. A recent study provided mechanistic information on the adjuvanticity of RNActive vaccines in mice in vivo and human cells in vitro54. Potent activation of TLR7 (mouse and human) and TLR8 (human) and production of type I interferon, pro-inflammatory cytokines and chemokines after intradermal immunization was shown54. A similar adjuvant activity was also demonstrated in the context of non-mRNA-based vaccines using RNAdjuvant (CureVac AG), an unmodified, single-stranded RNA stabilized by a cationic carrier peptide57.
Progress in mRNA vaccine delivery
Efficient in vivo mRNA delivery is critical to achieving therapeutic relevance. Exogenous mRNA must penetrate the barrier of the lipid membrane in order to reach the cytoplasm to be translated to functional protein. mRNA uptake mechanisms seem to be cell type dependent, and the physicochemical properties of the mRNA complexes can profoundly influence cellular delivery and organ distribution. There are two basic approaches for the delivery of mRNA vaccines that have been described to date. First, loading of mRNA into DCs ex vivo, followed by re-infusion of the transfected cells58; and second, direct parenteral injection of mRNA with or without a carrier. Ex vivo DC loading allows precise control of the cellular target, transfection efficiency and other cellular conditions, but as a form of cell therapy, it is an expensive and labour-intensive approach to vaccination. Direct injection of mRNA is comparatively rapid and cost-effective, but it does not yet allow precise and efficient cell-type-specific delivery, although there has been recent progress in this regard59. Both of these approaches have been explored in a variety of forms (Fig. 2; Table 1).

Commonly used delivery methods and carrier molecules for mRNA vaccines along with typical diameters for particulate complexes are shown: naked mRNA (part a); naked mRNA with in vivo electroporation (part b); protamine (cationic peptide)-complexed mRNA (part c); mRNA associated with a positively charged oil-in-water cationic nanoemulsion (part d); mRNA associated with a chemically modified dendrimer and complexed with polyethylene glycol (PEG)-lipid (part e); protamine-complexed mRNA in a PEG-lipid nanoparticle (part f); mRNA associated with a cationic polymer such as polyethylenimine (PEI) (part g); mRNA associated with a cationic polymer such as PEI and a lipid component (part h); mRNA associated with a polysaccharide (for example, chitosan) particle or gel (part i); mRNA in a cationic lipid nanoparticle (for example, 1,2-dioleoyloxy-3-trimethylammoniumpropane (DOTAP) or dioleoylphosphatidylethanolamine (DOPE) lipids) (part j); mRNA complexed with cationic lipids and cholesterol (part k); and mRNA complexed with cationic lipids, cholesterol and PEG-lipid (part l).
Ex vivo loading of DCs. DCs are the most potent antigen-presenting cells of the immune system. They initiate the adaptive immune response by internalizing and proteolytically processing antigens and presenting them to CD8+ and CD4+ T cells on major histocompatibility complexes (MHCs), namely, MHC class I and MHC class II, respectively. Additionally, DCs may present intact antigen to B cells to provoke an antibody response60. DCs are also highly amenable to mRNA transfection. For these reasons, DCs represent an attractive target for transfection by mRNA vaccines, both in vivo and ex vivo.
Although DCs have been shown to internalize naked mRNA through a variety of endocytic pathways61,62,63, ex vivo transfection efficiency is commonly increased using electroporation; in this case, mRNA molecules pass through membrane pores formed by a high-voltage pulse and directly enter the cytoplasm (reviewed in Ref. 64). This mRNA delivery approach has been favoured for its ability to generate high transfection efficiency without the need for a carrier molecule. DCs that are loaded with mRNA ex vivo are then re-infused into the autologous vaccine recipient to initiate the immune response. Most ex vivo-loaded DC vaccines elicit a predominantly cell-mediated immune response; thus, they have been used primarily to treat cancer (reviewed in Ref. 58).
Injection of naked mRNA in vivo. Naked mRNA has been used successfully for in vivo immunizations, particularly in formats that preferentially target antigen-presenting cells, as in intradermal61,65 and intranodal injections66,67,68. Notably, a recent report showed that repeated intranodal immunizations with naked, unmodified mRNA encoding tumour-associated neoantigens generated robust T cell responses and increased progression-free survival68 (discussed further in Box 2).
Physical delivery methods in vivo. To increase the efficiency of mRNA uptake in vivo, physical methods have occasionally been used to penetrate the cell membrane. An early report showed that mRNA complexed with gold particles could be expressed in tissues using a gene gun, a microprojectile method69. The gene gun was shown to be an efficient RNA delivery and vaccination method in mouse models70,71,72,73, but no efficacy data in large animals or humans are available. In vivo electroporation has also been used to increase uptake of therapeutic RNA74,75,76; however, in one study, electroporation increased the immunogenicity of only a self-amplifying RNA and not a non-replicating mRNA-based vaccine74. Physical methods can be limited by increased cell death and restricted access to target cells or tissues. Recently, the field has instead favoured the use of lipid or polymer-based nanoparticles as potent and versatile delivery vehicles.
Protamine. The cationic peptide protamine has been shown to protect mRNA from degradation by serum RNases77; however, protamine-complexed mRNA alone demonstrated limited protein expression and efficacy in a cancer vaccine model, possibly owing to an overly tight association between protamine and mRNA36,78. This issue was resolved by developing the RNActive vaccine platform, in which protamine-formulated RNA serves only as an immune activator and not as an expression vector52.
Cationic lipid and polymer-based delivery. Highly efficient mRNA transfection reagents based on cationic lipids or polymers, such as TransIT-mRNA (Mirus Bio LLC) or Lipofectamine (Invitrogen), are commercially available and work well in many primary cells and cancer cell lines9,13, but they often show limited in vivo efficacy or a high level of toxicity (N.P. and D.W., unpublished observations). Great progress has been made in developing similarly designed complexing reagents for safe and effective in vivo use, and these are discussed in detail in several recent reviews10,11,79,80. Cationic lipids and polymers, including dendrimers, have become widely used tools for mRNA administration in the past few years. The mRNA field has clearly benefited from the substantial investment in in vivo small interfering RNA (siRNA) administration, where these delivery vehicles have been used for over a decade. Lipid nanoparticles (LNPs) have become one of the most appealing and commonly used mRNA delivery tools. LNPs often consist of four components: an ionizable cationic lipid, which promotes self-assembly into virus-sized (~100 nm) particles and allows endosomal release of mRNA to the cytoplasm; lipid-linked polyethylene glycol (PEG), which increases the half-life of formulations; cholesterol, a stabilizing agent; and naturally occurring phospholipids, which support lipid bilayer structure. Numerous studies have demonstrated efficient in vivo siRNA delivery by LNPs (reviewed in Ref. 81), but it has only recently been shown that LNPs are potent tools for in vivo delivery of self-amplifying RNA19 and conventional, non-replicating mRNA21. Systemically delivered mRNA–LNP complexes mainly target the liver owing to binding of apolipoprotein E and subsequent receptor-mediated uptake by hepatocytes82, and intradermal, intramuscular and subcutaneous administration have been shown to produce prolonged protein expression at the site of the injection21,22. The mechanisms of mRNA escape into the cytoplasm are incompletely understood, not only for artificial liposomes but also for naturally occurring exosomes83. Further research into this area will likely be of great benefit to the field of therapeutic RNA delivery.
The magnitude and duration of in vivo protein production from mRNA–LNP vaccines can be controlled in part by varying the route of administration. Intramuscular and intradermal delivery of mRNA–LNPs has been shown to result in more persistent protein expression than systemic delivery routes: in one experiment, the half-life of mRNA-encoded firefly luciferase was roughly threefold longer after intradermal injection than after intravenous delivery21. These kinetics of mRNA–LNP expression may be favourable for inducing immune responses. A recent study demonstrated that sustained antigen availability during vaccination was a driver of high antibody titres and germinal centre (GC) B cell and T follicular helper (TFH) cell responses84. This process was potentially a contributing factor to the potency of recently described nucleoside-modified mRNA–LNP vaccines delivered by the intramuscular and intradermal routes20,22,85. Indeed, TFH cells have been identified as a critical population of immune cells that vaccines must activate in order to generate potent and long-lived neutralizing antibody responses, particularly against viruses that evade humoral immunity86. The dynamics of the GC reaction and the differentiation of TFH cells are incompletely understood, and progress in these areas would undoubtedly be fruitful for future vaccine design (Box 3).
mRNA vaccines against infectious diseases
Development of prophylactic or therapeutic vaccines against infectious pathogens is the most efficient means to contain and prevent epidemics. However, conventional vaccine approaches have largely failed to produce effective vaccines against challenging viruses that cause chronic or repeated infections, such as HIV-1, herpes simplex virus and respiratory syncytial virus (RSV). Additionally, the slow pace of commercial vaccine development and approval is inadequate to respond to the rapid emergence of acute viral diseases, as illustrated by the 2014–2016 outbreaks of the Ebola and Zika viruses. Therefore, the development of more potent and versatile vaccine platforms is crucial.
Preclinical studies have created hope that mRNA vaccines will fulfil many aspects of an ideal clinical vaccine: they have shown a favourable safety profile in animals, are versatile and rapid to design for emerging infectious diseases, and are amenable to scalable good manufacturing practice (GMP) production (already under way by several companies). Unlike protein immunization, several formats of mRNA vaccines induce strong CD8+ T cell responses, likely owing to the efficient presentation of endogenously produced antigens on MHC class I molecules, in addition to potent CD4+ T cell responses56,87,88. Additionally, unlike DNA immunization, mRNA vaccines have shown the ability to generate potent neutralizing antibody responses in animals with only one or two low-dose immunizations20,22,85. As a result, mRNA vaccines have elicited protective immunity against a variety of infectious agents in animal models19,20,22,56,89,90 and have therefore generated substantial optimism. However, recently published results from two clinical trials of mRNA vaccines for infectious diseases were somewhat modest, leading to more cautious expectations about the translation of preclinical success to the clinic22,91 (discussed further below).
Two major types of RNA vaccine have been utilized against infectious pathogens: self-amplifying or replicon RNA vaccines and non-replicating mRNA vaccines. Non-replicating mRNA vaccines can be further distinguished by their delivery method: ex vivo loading of DCs or direct in vivo injection into a variety of anatomical sites. As discussed below, a rapidly increasing number of preclinical studies in these areas have been published recently, and several have entered human clinical trials (Table 2).
Self-amplifying mRNA vaccines
Most currently used self-amplifying mRNA (SAM) vaccines are based on an alphavirus genome92, where the genes encoding the RNA replication machinery are intact but the genes encoding the structural proteins are replaced with the antigen of interest. The full-length RNA is ~9 kb long and can be easily produced by IVT from a DNA template. The SAM platform enables a large amount of antigen production from an extremely small dose of vaccine owing to intracellular replication of the antigen-encoding RNA. An early study reported that immunization with 10 μg of naked SAM vaccine encoding RSV fusion (F), influenza virus haemagglutinin (HA) or louping ill virus pre-membrane and envelope (prM-E) proteins resulted in antibody responses and partial protection from lethal viral challenges in mice93. The development of RNA complexing agents brought remarkable improvement to the efficacy of SAM vaccines. As little as 100 ng of an RNA replicon vaccine encoding RSV F, complexed to LNP, resulted in potent T and B cell immune responses in mice, and 1 μg elicited protective immune responses against RSV infection in a cotton rat intranasal challenge system19. SAM vaccines encoding influenza virus antigens in LNPs or an oil-in-water cationic nanoemulsion induced potent immune responses in ferrets and conferred protection from homologous and heterologous viral challenge in mice94,95,96. Further studies demonstrated the immunogenicity of this vaccine platform against diverse viruses in multiple species, including human cytomegalovirus (CMV), hepatitis C virus and rabies virus in mice, HIV-1 in rabbits, and HIV-1 and human CMV in rhesus macaques50,87,97. Replicon RNA encoding influenza antigens, complexed with chitosan-containing LNPs or polyethylenimine (PEI), has elicited T and B cell immune responses in mice after subcutaneous delivery98,99. Chahal and colleagues developed a delivery platform consisting of a chemically modified, ionizable dendrimer complexed into LNPs89. Using this platform, they demonstrated that intramuscular delivery of RNA replicons encoding influenza virus, Ebola virus or Toxoplasma gondii antigens protected mice against lethal infection89. The same group recently demonstrated that vaccination with an RNA replicon encoding Zika virus prM-E formulated in the same manner elicited antigen-specific antibody and CD8+ T cell responses in mice88. Another recent study reported immunogenicity and moderate protective efficacy of SAM vaccines against bacterial pathogens, namely Streptococcus (groups A and B) spp., further demonstrating the versatility of this platform100.
One of the advantages of SAM vaccines is that they create their own adjuvants in the form of dsRNA structures, replication intermediates and other motifs that may contribute to their high potency. However, the intrinsic nature of these PAMPs may make it difficult to modulate the inflammatory profile or reactogenicity of SAM vaccines. Additionally, size constraints of the insert are greater for SAM vaccines than for mRNAs that do not encode replicon genes, and the immunogenicity of the replication proteins may theoretically limit repeated use.
Dendritic cell mRNA vaccines
As described above, ex vivo DC loading is a heavily pursued method to generate cell-mediated immunity against cancer. Development of infectious disease vaccines using this approach has been mainly limited to a therapeutic vaccine for HIV-1: HIV-1-infected individuals on highly active antiretroviral therapy were treated with autologous DCs electroporated with mRNA encoding various HIV-1 antigens, and cellular immune responses were evaluated101,102,103,104,105,106. This intervention proved to be safe and elicited antigen-specific CD4+ and CD8+ T cell responses, but no clinical benefit was observed. Another study in humans evaluated a CMV pp65 mRNA-loaded DC vaccination in healthy human volunteers and allogeneic stem cell recipients and reported induction or expansion of CMV-specific cellular immune responses107.
Direct injection of non-replicating mRNA vaccines
Directly injectable, non-replicating mRNA vaccines are an appealing vaccine format owing to their simple and economical administration, particularly in resource-limited settings. Although an early report demonstrated that immunization with liposome-complexed mRNA encoding influenza virus nucleoproteins elicited CTL responses in mice108, the first demonstration of protective immune responses by mRNA vaccines against infectious pathogens was published only a few years ago18. This seminal work demonstrated that intradermally administered uncomplexed mRNA encoding various influenza virus antigens combined with a protamine-complexed RNA adjuvant was immunogenic in multiple animal models and protected mice from lethal viral challenge.
Immunization with the protamine-based RNActive platform encoding rabies virus glycoprotein has also induced protective immunity against a lethal intracerebral virus challenge in mice and potent neutralizing antibody responses in pigs56. In a recently published seminal work, Alberer and colleagues evaluated the safety and immunogenicity of this vaccine in 101 healthy human volunteers91. Subjects received 80–640 μg of mRNA vaccine three times by needle-syringe or needle-free devices, either intradermally or intramuscularly. Seven days after vaccination, nearly all participants reported mild to moderate injection site reactions, and 78% experienced a systemic reaction (for example, fever, headache and chills). There was one serious adverse event that was possibly related to the vaccine: a transient and moderate case of Bell palsy. Surprisingly, the needle-syringe injections did not generate detectable neutralizing antibodies in 98% of recipients. By contrast, needle-free delivery induced variable levels of neutralizing antibodies, the majority of which peaked above the expected protective threshold but then largely waned after 1 year in subjects who were followed up long term. Elucidating the basis of the disparate immunogenicity between the animals and humans who received this vaccine and between the two routes of delivery will be informative for future vaccine design using this platform.
Other infectious disease vaccines have successfully utilized lipid- or polymer-based delivery systems. Cationic 1,2-dioleoyloxy-3-trimethylammoniumpropane (DOTAP) and dioleoylphosphatidylethanolamine (DOPE) lipid-complexed mRNA encoding HIV-1 gag generated antigen-specific CD4+ and CD8+ T cell responses after subcutaneous delivery in mice109. Two other studies demonstrated that PEI-complexed mRNAs could be efficiently delivered to mice to induce HIV-1-specific immune responses: subcutaneously delivered mRNA encoding HIV-1 gag elicited CD4+ and CD8+ T cell responses, and intranasally administered mRNA encoding the HIV-1 envelope gp120 subunit crossed the nasal epithelium and generated antigen-specific immune responses in the nasal cavity110,111. Kranz and colleagues also performed intravenous immunizations in mice using lipid-complexed mRNA encoding influenza virus HA and showed evidence of T cell activation after a single dose59.
Nucleoside-modified mRNA vaccines represent a new and highly efficacious category of mRNA vaccines. Owing to the novelty of this immunization platform, our knowledge of efficacy is limited to the results of four recent publications that demonstrated the potency of such vaccines in small and large animals. The first published report demonstrated that a single intradermal injection of LNP-formulated mRNA encoding Zika virus prM-E, modified with 1-methylpseudouridine and FPLC purification, elicited protective immune responses in mice and rhesus macaques with the use of as little as 50 μg (0.02 mg kg−1) of vaccine in macaques20. A subsequent study by a different group tested a similarly designed vaccine against Zika virus in mice and found that a single intramuscular immunization elicited moderate immune responses, and a booster vaccination resulted in potent and protective immune responses85. This vaccine also incorporated the modified nucleoside 1-methylpseudouridine, but FPLC purification or other methods of removing dsRNA contaminants were not reported. Notably, this report showed that antibody-dependent enhancement of secondary infection with a heterologous flavivirus, a major concern for dengue and Zika virus vaccines, could be diminished by removing a cross-reactive epitope in the E protein. A recent follow-up study evaluated the same vaccine in a model of maternal vaccination and fetal infection112. Two immunizations reduced Zika virus infection in fetal mice by several orders of magnitude and completely rescued a defect in fetal viability.
Another recent report evaluated the immunogenicity of LNP-complexed, nucleoside-modified, non-FPLC-purified mRNA vaccines against influenza HA 10 neuraminidase 8 (H10N8) and H7N9 influenza viruses in mice, ferrets, non-human primates and, for the first time, humans22. A single intradermal or intramuscular immunization with low doses (0.4–10 μg) of LNP-complexed mRNA encoding influenza virus HA elicited protective immune responses against homologous influenza virus challenge in mice. Similar results were obtained in ferrets and cynomolgus monkeys after immunization with one or two doses of 50–400 μg of a vaccine containing LNP-complexed mRNA encoding HA, corroborating that the potency of mRNA–LNP vaccines translates to larger animals, including non-human primates.
On the basis of encouraging preclinical data, two phase I clinical trials have recently been initiated to evaluate the immunogenicity and safety of nucleoside-modified mRNA–LNP vaccines in humans for the first time. The mRNA vaccine encoding H10N8 HA is currently undergoing clinical testing (NCT03076385), and interim findings for 23 vaccinated individuals have been reported22. Participants received a small amount (100 μg) of vaccine intramuscularly, and immunogenicity was measured 43 days after vaccination. The vaccine proved to be immunogenic in all subjects, as measured by haemagglutination inhibition and microneutralization antibody assays. Promisingly, antibody titres were above the expected protective threshold, but they were moderately lower than in the animal models. Similarly to the study by Alberer et al.91, most vaccinated subjects reported mild to moderate reactogenicity (injection site pain, myalgia, headache, fatigue and chills), and three subjects reported severe injection site reactions or a systemic common cold-like response. This level of reactogenicity appears to be similar to that of more traditional vaccine formats113,114. Finally, the Zika virus vaccine described by Richner et al.85,112 is also entering clinical evaluation in a combined phase I/II trial (NCT03014089). Future studies that apply nucleoside-modified mRNA–LNP vaccines against a greater diversity of antigens will reveal the extent to which this strategy is broadly applicable to infectious disease vaccines.
mRNA cancer vaccines
mRNA-based cancer vaccines have been recently and extensively reviewed115,116,117,118,119. Below, the most recent advances and directions are highlighted. Cancer vaccines and other immunotherapies represent promising alternative strategies to treat malignancies. Cancer vaccines can be designed to target tumour-associated antigens that are preferentially expressed in cancerous cells, for example, growth-associated factors, or antigens that are unique to malignant cells owing to somatic mutation120. These neoantigens, or the neoepitopes within them, have been deployed as mRNA vaccine targets in humans121 (Box 2). Most cancer vaccines are therapeutic, rather than prophylactic, and seek to stimulate cell-mediated responses, such as those from CTLs, that are capable of clearing or reducing tumour burden122. The first proof-of-concept studies that not only proposed the idea of RNA cancer vaccines but also provided evidence of the feasibility of this approach were published more than two decades ago123,124. Since then, numerous preclinical and clinical studies have demonstrated the viability of mRNA vaccines to combat cancer (Table 3).
DC mRNA cancer vaccines
As DCs are central players in initiating antigen-specific immune responses, it seemed logical to utilize them for cancer immunotherapy. The first demonstration that DCs electroporated with mRNA could elicit potent immune responses against tumour antigens was reported by Boczkowski and colleagues in 1996 (Ref. 124). In this study, DCs pulsed with ovalbumin (OVA)-encoding mRNA or tumour-derived RNAs elicited a tumour-reducing immune response in OVA-expressing and other melanoma models in mice. A variety of immune regulatory proteins have been identified in the form of mRNA-encoded adjuvants that can increase the potency of DC cancer vaccines. Several studies demonstrated that electroporation of DCs with mRNAs encoding co-stimulatory molecules such as CD83, tumour necrosis factor receptor superfamily member 4 (TNFRSF4; also known as OX40) and 4-1BB ligand (4-1BBL) resulted in a substantial increase in the immune stimulatory activity of DCs125,126,127,128. DC functions can also be modulated through the use of mRNA-encoded pro-inflammatory cytokines, such as IL-12, or trafficking-associated molecules129,130,131. As introduced above, TriMix is a cocktail of mRNA-encoded adjuvants (CD70, CD40L and constitutively active TLR4) that can be electroporated in combination with antigen-encoding mRNA or mRNAs132. This formulation proved efficacious in multiple preclinical studies by increasing DC activation and shifting the CD4+ T cell phenotype from T regulatory cells to T helper 1 (TH1)-like cells132,133,134,135,136. Notably, the immunization of patients with stage III or stage IV melanoma using DCs loaded with mRNA encoding melanoma-associated antigens and TriMix adjuvant resulted in tumour regression in 27% of treated individuals137. Multiple clinical trials have now been conducted using DC vaccines targeting various cancer types, such as metastatic prostate cancer, metastatic lung cancer, renal cell carcinoma, brain cancers, melanoma, acute myeloid leukaemia, pancreatic cancer and others138,139 (reviewed in Refs 51,58).
A new line of research combines mRNA electroporation of DCs with traditional chemotherapy agents or immune checkpoint inhibitors. In one trial, patients with stage III or IV melanoma were treated with ipilimumab, a monoclonal antibody against CTL antigen 4 (CTLA4), and DCs loaded with mRNA encoding melanoma-associated antigens plus TriMix. This intervention resulted in durable tumour reduction in a proportion of individuals with recurrent or refractory melanoma140.
Direct injection of mRNA cancer vaccines
The route of administration and delivery format of mRNA vaccines can greatly influence outcomes. A variety of mRNA cancer vaccine formats have been developed using common delivery routes (intradermal, intramuscular, subcutaneous or intranasal) and some unconventional routes of vaccination (intranodal, intravenous, intrasplenic or intratumoural).
Intranodal administration of naked mRNA is an unconventional but efficient means of vaccine delivery. Direct mRNA injection into secondary lymphoid tissue offers the advantage of targeted antigen delivery to antigen-presenting cells at the site of T cell activation, obviating the need for DC migration. Several studies have demonstrated that intranodally injected naked mRNA can be selectively taken up by DCs and can elicit potent prophylactic or therapeutic antitumour T cell responses62,66; an early study also demonstrated similar findings with intrasplenic delivery141. Coadministration of the DC-activating protein FMS-related tyrosine kinase 3 ligand (FLT3L) was shown in some cases to further improve immune responses to intranodal mRNA vaccination142,143. Incorporation of the TriMix adjuvant into intranodal injections of mice with mRNAs encoding tumour-associated antigens resulted in potent antigen-specific CTL responses and tumour control in multiple tumour models133. A more recent study demonstrated that intranodal injection of mRNA encoding the E7 protein of human papillomavirus (HPV) 16 with TriMix increased the number of tumour-infiltrating CD8+ T cells and inhibited the growth of an E7-expressing tumour model in mice67.
The success of preclinical studies has led to the initiation of clinical trials using intranodally injected naked mRNA encoding tumour-associated antigens into patients with advanced melanoma (NCT01684241) and patients with hepatocellular carcinoma (EudraCT: 2012-005572-34). In one published trial, patients with metastatic melanoma were treated with intranodally administered DCs electroporated with mRNA encoding the melanoma-associated antigens tyrosinase or gp100 and TriMix, which induced limited antitumour responses144.
Intranasal vaccine administration is a needle-free, noninvasive manner of delivery that enables rapid antigen uptake by DCs. Intranasally delivered mRNA complexed with Stemfect (Stemgent) LNPs resulted in delayed tumour onset and increased survival in prophylactic and therapeutic mouse tumour models using the OVA-expressing E.G7-OVA T lymphoblastic cell line145.
Intratumoural mRNA vaccination is a useful approach that offers the advantage of rapid and specific activation of tumour-resident T cells. Often, these vaccines do not introduce mRNAs encoding tumour-associated antigens but simply aim to activate tumour-specific immunity in situ using immune stimulatory molecules. An early study demonstrated that naked mRNA or protamine-stabilized mRNA encoding a non-tumour related gene (GLB1) impaired tumour growth and provided protection in a glioblastoma mouse model, taking advantage of the intrinsic immunogenic properties of mRNA146. A more recent study showed that intratumoural delivery of mRNA encoding an engineered cytokine based on interferon-β (IFNβ) fused to a transforming growth factor-β (TGFβ) antagonist increased the cytolytic capacity of CD8+ T cells and modestly delayed tumour growth in OVA-expressing lymphoma or lung carcinoma mouse models147. It has also been shown that intratumoural administration of TriMix mRNA that does not encode tumour-associated antigens results in activation of CD8α+ DCs and tumour-specific T cells, leading to delayed tumour growth in various mouse models148.
Systemic administration of mRNA vaccines is not common owing to concerns about aggregation with serum proteins and rapid extracellular mRNA degradation; thus, formulating mRNAs into carrier molecules is essential. As discussed above, numerous delivery formulations have been developed to facilitate mRNA uptake, increase protein translation and protect mRNA from RNases10,11,79,80. Another important issue is the biodistribution of mRNA vaccines after systemic delivery. Certain cationic LNP-based complexing agents delivered intravenously traffic mainly to the liver21, which may not be ideal for DC activation. An effective strategy for DC targeting of mRNA vaccines after systemic delivery has recently been described59. An mRNA–lipoplex (mRNA–liposome complex) delivery platform was generated using cationic lipids and neutral helper lipids formulated with mRNA, and it was discovered that the lipid-to-mRNA ratio, and thus the net charge of the particles, has a profound impact on the biodistribution of the vaccine. While a positively charged lipid particle primarily targeted the lung, a negatively charged particle targeted DCs in secondary lymphoid tissues and bone marrow. The negatively charged particle induced potent immune responses against tumour-specific antigens that were associated with impressive tumour reduction in various mouse models59. As no toxic effects were observed in mice or non-human primates, clinical trials using this approach to treat patients with advanced melanoma or triple-negative breast cancer have been initiated (NCT02410733 and NCT02316457).
A variety of antigen-presenting cells reside in the skin149, making it an ideal site for immunogen delivery during vaccination (Fig. 3). Thus, the intradermal route of delivery has been widely used for mRNA cancer vaccines. An early seminal study demonstrated that intradermal administration of total tumour RNA delayed tumour growth in a fibrosarcoma mouse model65. Intradermal injection of mRNA encoding tumour antigens in the protamine-based RNActive platform proved efficacious in various mouse models of cancer36 and in multiple prophylactic and therapeutic clinical settings (Table 3). One such study demonstrated that mRNAs encoding survivin and various melanoma tumour antigens resulted in increased numbers of antigen-specific T cells in a subset of patients with melanoma150. In humans with castration-resistant prostate cancer, an RNActive vaccine expressing multiple prostate cancer-associated proteins elicited antigen-specific T cell responses in the majority of recipients151. Lipid-based carriers have also contributed to the efficacy of intradermally delivered mRNA cancer vaccines. The delivery of OVA-encoding mRNA in DOTAP and/or DOPE liposomes resulted in antigen-specific CTL activity and inhibited growth of OVA-expressing tumours in mice152. In the same study, coadministration of mRNA encoding granulocyte–macrophage colony-stimulating factor (GM-CSF) improved OVA-specific cytolytic responses. Another report showed that subcutaneous delivery of LNP-formulated mRNA encoding two melanoma-associated antigens delayed tumour growth in mice, and co-delivery of lipopolysaccharide (LPS) in LNPs increased both CTL and antitumour activity153. In general, mRNA cancer vaccines have proved immunogenic in humans, but further refinement of vaccination methods, as informed by basic immunological research, will likely be necessary to achieve greater clinical benefits.

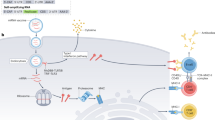
For an injected mRNA vaccine, major considerations for effectiveness include the following: the level of antigen expression in professional antigen-presenting cells (APCs), which is influenced by the efficiency of the carrier, by the presence of pathogen-associated molecular patterns (PAMPs) in the form of double-stranded RNA (dsRNA) or unmodified nucleosides and by the level of optimization of the RNA sequence (codon usage, G:C content, 5′ and 3′ untranslated regions (UTRs) and so on); dendritic cell (DC) maturation and migration to secondary lymphoid tissue, which is increased by PAMPs; and the ability of the vaccine to activate robust T follicular helper (TFH) cell and germinal centre (GC) B cell responses — an area that remains poorly understood. An intradermal injection is shown as an example. EC, extracellular.
The combination of mRNA vaccination with adjunctive therapies, such as traditional chemotherapy, radiotherapy and immune checkpoint inhibitors, has increased the beneficial outcome of vaccination in some preclinical studies154,155. For example, cisplatin treatment significantly increased the therapeutic effect of immunizing with mRNA encoding the HPV16 E7 oncoprotein and TriMix, leading to the complete rejection of female genital tract tumours in a mouse model67. Notably, it has also been suggested that treatment with antibodies against programmed cell death protein 1 (PD1) increased the efficacy of a neoepitope mRNA-based vaccine against metastatic melanoma in humans, but more data are required to explore this hypothesis68.
Therapeutic considerations and challenges
Good manufacturing practice production
mRNA is produced by in vitro reactions with recombinant enzymes, ribonucleotide triphosphates (NTPs) and a DNA template; thus, it is rapid and relatively simple to produce in comparison with traditional protein subunit and live or inactivated virus vaccine production platforms. Its reaction yield and simplicity make rapid mRNA production possible in a small GMP facility footprint. The manufacturing process is sequence-independent and is primarily dictated by the length of the RNA, the nucleotide and capping chemistry and the purification of the product; however, it is possible that certain sequence properties such as extreme length may present difficulties (D.W., unpublished observations). According to current experience, the process can be standardized to produce nearly any encoded protein immunogen, making it particularly suitable for rapid response to emerging infectious diseases.
All enzymes and reaction components required for the GMP production of mRNA can be obtained from commercial suppliers as synthesized chemicals or bacterially expressed, animal component-free reagents, thereby avoiding safety concerns surrounding the adventitious agents that plague cell-culture-based vaccine manufacture. All the components, such as plasmid DNA, phage polymerases, capping enzymes and NTPs, are readily available as GMP-grade traceable components; however, some of these are currently available at only limited scale or high cost. As mRNA therapeutics move towards commercialization and the scale of production increases, more economical options may become accessible for GMP source materials.
GMP production of mRNA begins with DNA template production followed by enzymatic IVT and follows the same multistep protocol that is used for research scale synthesis, with added controls to ensure the safety and potency of the product16. Depending on the specific mRNA construct and chemistry, the protocol may be modified slightly from what is described here to accommodate modified nucleosides, capping strategies or template removal. To initiate the production process, template plasmid DNA produced in Escherichia coli is linearized using a restriction enzyme to allow synthesis of runoff transcripts with a poly(A) tract at the 3′ end. Next, the mRNA is synthesized from NTPs by a DNA-dependent RNA polymerase from bacteriophage (such as T7, SP6, or T3). The template DNA is then degraded by incubation with DNase. Finally, the mRNA is enzymatically or chemically capped to enable efficient translation in vivo. mRNA synthesis is highly productive, yielding in excess of 2 g l−1 of full-length mRNA in multi-gram scale reactions under optimized conditions.
Once the mRNA is synthesized, it is processed though several purification steps to remove reaction components, including enzymes, free nucleotides, residual DNA and truncated RNA fragments. While LiCl precipitation is routinely used for laboratory-scale preparation, purification at the clinical scale utilizes derivatized microbeads in batch or column formats, which are easier to utilize at large scale156,157. For some mRNA platforms, removal of dsRNA and other contaminants is critical for the potency of the final product, as it is a potent inducer of interferon-dependent translation inhibition. This has been accomplished by reverse-phase FPLC at the laboratory scale158, and scalable aqueous purification approaches are being investigated. After mRNA is purified, it is exchanged into a final storage buffer and sterile-filtered for subsequent filling into vials for clinical use. RNA is susceptible to degradation by both enzymatic and chemical pathways157. Formulation buffers are tested to ensure that they are free of contaminating RNases and may contain buffer components, such as antioxidants and chelators, which minimize the effects of reactive oxygen species and divalent metal ions that lead to mRNA instability159.
Pharmaceutical formulation of mRNAs is an active area of development. Although most products for early phase studies are stored frozen (−70 °C), efforts to develop formulations that are stable at higher temperatures more suitable for vaccine distribution are continuing. Published reports suggest that stable refrigerated or room temperature formulations can be made. The RNActive platform was reported to be active after lyophilization and storage at 5–25 °C for 3 years and at 40 °C for 6 months91. Another report demonstrated that freeze-dried naked mRNA is stable for at least 10 months under refrigerated conditions160. The stability of mRNA products might also be improved by packaging within nanoparticles or by co-formulation with RNase inhibitors161. For lipid-encapsulated mRNA, at least 6 months of stability has been observed (Arbutus Biopharma, personal communication), but longer-term storage of such mRNA–lipid complexes in an unfrozen form has not yet been reported.
Regulatory aspects
There is no specific guidance from the FDA or European Medicines Agency (EMA) for mRNA vaccine products. However, the increasing number of clinical trials conducted under EMA and FDA oversight indicate that regulators have accepted the approaches proposed by various organizations to demonstrate that products are safe and acceptable for testing in humans. Because mRNA falls into the broad vaccine category of genetic immunogens, many of the guiding principles that have been defined for DNA vaccines162 and gene therapy vectors163,164 can likely be applied to mRNA with some adaptations to reflect the unique features of mRNA. A detailed review of EMA regulations for RNA vaccines by Hinz and colleagues highlights the different regulatory paths stipulated for prophylactic infectious disease versus therapeutic applications165. Regardless of the specific classification within existing guidelines, some themes can be observed in what is stated in these guidance documents and in what has been reported for recently published clinical studies. In particular, the recent report of an mRNA vaccine against influenza virus highlights preclinical and clinical data demonstrating biodistribution and persistence in mice, disease protection in a relevant animal model (ferrets), and immunogenicity, local reactogenicity and toxicity in humans22. As mRNA products become more prominent in the vaccine field, it is likely that specific guidance will be developed that will delineate requirements to produce and evaluate new mRNA vaccines.
Safety
The requirement for safety in modern prophylactic vaccines is extremely stringent because the vaccines are administered to healthy individuals. Because the manufacturing process for mRNA does not require toxic chemicals or cell cultures that could be contaminated with adventitious viruses, mRNA production avoids the common risks associated with other vaccine platforms, including live virus, viral vectors, inactivated virus and subunit protein vaccines. Furthermore, the short manufacturing time for mRNA presents few opportunities to introduce contaminating microorganisms. In vaccinated people, the theoretical risks of infection or integration of the vector into host cell DNA are not a concern for mRNA. For the above reasons, mRNA vaccines have been considered a relatively safe vaccine format.
Several different mRNA vaccines have now been tested from phase I to IIb clinical studies and have been shown to be safe and reasonably well tolerated (Tables 2, 3). However, recent human trials have demonstrated moderate and in rare cases severe injection site or systemic reactions for different mRNA platforms22,91. Potential safety concerns that are likely to be evaluated in future preclinical and clinical studies include local and systemic inflammation, the biodistribution and persistence of expressed immunogen, stimulation of auto-reactive antibodies and potential toxic effects of any non-native nucleotides and delivery system components. A possible concern could be that some mRNA-based vaccine platforms54,166 induce potent type I interferon responses, which have been associated not only with inflammation but also potentially with autoimmunity167,168. Thus, identification of individuals at an increased risk of autoimmune reactions before mRNA vaccination may allow reasonable precautions to be taken. Another potential safety issue could derive from the presence of extracellular RNA during mRNA vaccination. Extracellular naked RNA has been shown to increase the permeability of tightly packed endothelial cells and may thus contribute to oedema169. Another study showed that extracellular RNA promoted blood coagulation and pathological thrombus formation170. Safety will therefore need continued evaluation as different mRNA modalities and delivery systems are utilized for the first time in humans and are tested in larger patient populations.
Conclusions and future directions
Currently, mRNA vaccines are experiencing a burst in basic and clinical research. The past 2 years alone have witnessed the publication of dozens of preclinical and clinical reports showing the efficacy of these platforms. Whereas the majority of early work in mRNA vaccines focused on cancer applications, a number of recent reports have demonstrated the potency and versatility of mRNA to protect against a wide variety of infectious pathogens, including influenza virus, Ebola virus, Zika virus, Streptococcus spp. and T. gondii (Tables 1,2).
While preclinical studies have generated great optimism about the prospects and advantages of mRNA-based vaccines, two recent clinical reports have led to more tempered expectations22,91. In both trials, immunogenicity was more modest in humans than was expected based on animal models, a phenomenon also observed with DNA-based vaccines171, and the side effects were not trivial. We caution that these trials represent only two variations of mRNA vaccine platforms, and there may be substantial differences when the expression and immunostimulatory profiles of the vaccine are changed. Further research is needed to determine how different animal species respond to mRNA vaccine components and inflammatory signals and which pathways of immune signalling are most effective in humans.
Recent advances in understanding and reducing the innate immune sensing of mRNA have aided efforts not only in active vaccination but also in several applications of passive immunization or passive immunotherapy for infectious diseases and cancer (Box 4). Direct comparisons between mRNA expression platforms should clarify which systems are most appropriate for both passive and active immunization. Given the large number of promising mRNA platforms, further head-to-head comparisons would be of utmost value to the vaccine field because this would allow investigators to focus resources on those best suited for each application.
The fast pace of progress in mRNA vaccines would not have been possible without major recent advances in the areas of innate immune sensing of RNA and in vivo delivery methods. Extensive basic research into RNA and lipid and polymer biochemistry has made it possible to translate mRNA vaccines into clinical trials and has led to an astonishing level of investment in mRNA vaccine companies (Table 4). Moderna Therapeutics, founded in 2010, has raised almost US$2 billion in capital with a plan to commercialize mRNA-based vaccines and therapies172,173. The US Biomedical Advanced Research and Development Authority (BARDA) has committed support for Moderna's clinical evaluation of a promising nucleoside-modified mRNA vaccine for Zika virus (NCT03014089). In Germany, CureVac AG has an expanding portfolio of therapeutic targets174, including both cancer and infectious diseases, and BioNTech is developing an innovative approach to personalized cancer medicine using mRNA vaccines121 (Box 2). The translation of basic research into clinical testing is also made more expedient by the commercialization of custom GMP products by companies such as New England Biolabs and Aldevron175. Finally, the recent launch of the Coalition for Epidemic Preparedness Innovations (CEPI) provides great optimism for future responses to emerging viral epidemics. This multinational public and private partnership aims to raise $1 billion to develop platform-based vaccines, such as mRNA, to rapidly contain emerging outbreaks before they spread out of control.
The future of mRNA vaccines is therefore extremely bright, and the clinical data and resources provided by these companies and other institutions are likely to substantially build on and invigorate basic research into mRNA-based therapeutics.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
World Health Organization. Immunization coverage. World Health Organization http://www.who.int/mediacentre/factsheets/fs378/en (2017).
Younger, D. S., Younger, A. P. & Guttmacher, S. Childhood vaccination: implications for global and domestic public health. Neurol. Clin. 34, 1035–1047 (2016).
Plotkin, S. A. Vaccines: the fourth century. Clin. Vaccine Immunol. 16, 1709–1719 (2009).
Rodrigues, C. M. C., Pinto, M. V., Sadarangani, M. & Plotkin, S. A. Whither vaccines? J. Infect. 74 (Suppl. 1), S2–S9 (2017).
Wolff, J. A. et al. Direct gene transfer into mouse muscle in vivo. Science 247, 1465–1468 (1990). This study demonstrates protein production from RNA administered in vivo.
Jirikowski, G. F., Sanna, P. P., Maciejewski-Lenoir, D. & Bloom, F. E. Reversal of diabetes insipidus in Brattleboro rats: intrahypothalamic injection of vasopressin mRNA. Science 255, 996–998 (1992).
Suschak, J. J., Williams, J. A. & Schmaljohn, C. S. Advancements in DNA vaccine vectors, non-mechanical delivery methods, and molecular adjuvants to increase immunogenicity. Hum. Vaccin. Immunother. 13, 2837–2848 (2017).
Tandrup Schmidt, S., Foged, C., Korsholm, K. S., Rades, T. & Christensen, D. Liposome-based adjuvants for subunit vaccines: formulation strategies for subunit antigens and immunostimulators. Pharmaceutics 8, E7 (2016).
Kariko, K. et al. Incorporation of pseudouridine into mRNA yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol. Ther. 16, 1833–1840 (2008).
Kauffman, K. J., Webber, M. J. & Anderson, D. G. Materials for non-viral intracellular delivery of messenger RNA therapeutics. J. Control. Release 240, 227–234 (2016).
Guan, S. & Rosenecker, J. Nanotechnologies in delivery of mRNA therapeutics using nonviral vector-based delivery systems. Gene Ther. 24, 133–143 (2017).
Thess, A. et al. Sequence-engineered mRNA without chemical nucleoside modifications enables an effective protein therapy in large animals. Mol. Ther. 23, 1456–1464 (2015).
Kariko, K., Muramatsu, H., Ludwig, J. & Weissman, D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 39, e142 (2011). This study demonstrates the importance of purification of IVT mRNA in achieving potent protein translation and in suppressing inflammatory responses.
Weissman, D. mRNA transcript therapy. Expert Rev. Vaccines 14, 265–281 (2015).
Sahin, U., Kariko, K. & Tureci, O. mRNA-based therapeutics — developing a new class of drugs. Nat. Rev. Drug Discov. 13, 759–780 (2014). This is a useful Review covering vaccine and non-vaccine forms of mRNA therapeutics.
Pardi, N., Muramatsu, H., Weissman, D. & Kariko, K. In vitro transcription of long RNA containing modified nucleosides. Methods Mol. Biol. 969, 29–42 (2013).
Tsui, N. B., Ng, E. K. & Lo, Y. M. Stability of endogenous and added RNA in blood specimens, serum, and plasma. Clin. Chem. 48, 1647–1653 (2002).
Petsch, B. et al. Protective efficacy of in vitro synthesized, specific mRNA vaccines against influenza A virus infection. Nat. Biotechnol. 30, 1210–1216 (2012). This study demonstrates that directly injected, non-replicating mRNA can induce protective immune responses against an infectious pathogen.
Geall, A. J. et al. Nonviral delivery of self-amplifying RNA vaccines. Proc. Natl Acad. Sci. USA 109, 14604–14609 (2012). This important study demonstrates that the duration of in vivo protein production from RNA replicons can be greatly improved by packaging them into lipid nanoparticles.
Pardi, N. et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature 543, 248–251 (2017).
Pardi, N. et al. Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. J. Control. Release 217, 345–351 (2015).
Bahl, K. et al. Preclinical and clinical demonstration of immunogenicity by mRNA vaccines against H10N8 and H7N9 influenza viruses. Mol. Ther. 25, 1316–1327 (2017). This is a report of a clinical vaccine trial using directly injected, non-replicating, nucleoside-modified mRNA against an infectious pathogen.
Ross, J. & Sullivan, T. D. Half-lives of beta and gamma globin messenger RNAs and of protein synthetic capacity in cultured human reticulocytes. Blood 66, 1149–1154 (1985).
Holtkamp, S. et al. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood 108, 4009–4017 (2006).
Gallie, D. R. The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 5, 2108–2116 (1991).
Martin, S. A., Paoletti, E. & Moss, B. Purification of mRNA guanylyltransferase and mRNA (guanine-7-) methyltransferase from vaccinia virions. J. Biol. Chem. 250, 9322–9329 (1975).
Stepinski, J., Waddell, C., Stolarski, R., Darzynkiewicz, E. & Rhoads, R. E. Synthesis and properties of mRNAs containing the novel “anti-reverse” cap analogs 7-methyl(3′-O-methyl)GpppG and 7-methyl (3′-deoxy)GpppG. RNA 7, 1486–1495 (2001).
Malone, R. W., Felgner, P. L. & Verma, I. M. Cationic liposome-mediated RNA transfection. Proc. Natl Acad. Sci. USA 86, 6077–6081 (1989).
Gustafsson, C., Govindarajan, S. & Minshull, J. Codon bias and heterologous protein expression. Trends Biotechnol. 22, 346–353 (2004).
Mauro, V. P. & Chappell, S. A. A critical analysis of codon optimization in human therapeutics. Trends Mol. Med. 20, 604–613 (2014).
Kudla, G., Lipinski, L., Caffin, F., Helwak, A. & Zylicz, M. High guanine and cytosine content increases mRNA levels in mammalian cells. PLoS Biol. 4, e180 (2006).
Kudla, G., Murray, A. W., Tollervey, D. & Plotkin, J. B. Coding-sequence determinants of gene expression in Escherichia coli. Science 324, 255–258 (2009).
Buhr, F. et al. Synonymous codons direct cotranslational folding toward different protein conformations. Mol. Cell 61, 341–351 (2016).
Yu, C. H. et al. Codon usage influences the local rate of translation elongation to regulate co-translational protein folding. Mol. Cell 59, 744–754 (2015).
Chen, N. et al. RNA sensors of the innate immune system and their detection of pathogens. IUBMB Life 69, 297–304 (2017).
Fotin-Mleczek, M. et al. Messenger RNA-based vaccines with dual activity induce balanced TLR-7 dependent adaptive immune responses and provide antitumor activity. J. Immunother. 34, 1–15 (2011).
Rettig, L. et al. Particle size and activation threshold: a new dimension of danger signaling. Blood 115, 4533–4541 (2010).
de Haro, C., Mendez, R. & Santoyo, J. The eIF-2α kinases and the control of protein synthesis. FASEB J. 10, 1378–1387 (1996).
Liang, S. L., Quirk, D. & Zhou, A. RNase L: its biological roles and regulation. IUBMB Life 58, 508–514 (2006).
Zhang, Z. et al. Structural analysis reveals that Toll-like receptor 7 is a dual receptor for guanosine and single-stranded RNA. Immunity 45, 737–748 (2016).
Tanji, H. et al. Toll-like receptor 8 senses degradation products of single-stranded RNA. Nat. Struct. Mol. Biol. 22, 109–115 (2015).
Isaacs, A., Cox, R. A. & Rotem, Z. Foreign nucleic acids as the stimulus to make interferon. Lancet 2, 113–116 (1963).
Schwartz, S. et al. Transcriptome-wide mapping reveals widespread dynamic-regulated pseudouridylation of ncRNA and mRNA. Cell 159, 148–162 (2014).
Carlile, T. M. et al. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 515, 143–146 (2014).
Andries, O. et al. N1-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J. Control. Release 217, 337–344 (2015).
Anderson, B. R. et al. Incorporation of pseudouridine into mRNA enhances translation by diminishing PKR activation. Nucleic Acids Res. 38, 5884–5892 (2010).
Anderson, B. R. et al. Nucleoside modifications in RNA limit activation of 2′-5′-oligoadenylate synthetase and increase resistance to cleavage by RNase L. Nucleic Acids Res. 39, 9329–9338 (2011).
Kariko, K., Buckstein, M., Ni, H. & Weissman, D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity 23, 165–175 (2005). This report demonstrates that nucleoside modification of mRNA decreases inflammatory responses.
Kauffman, K. J. et al. Efficacy and immunogenicity of unmodified and pseudouridine-modified mRNA delivered systemically with lipid nanoparticles in vivo. Biomaterials 109, 78–87 (2016).
Brito, L. A. et al. A cationic nanoemulsion for the delivery of next-generation RNA vaccines. Mol. Ther. 22, 2118–2129 (2014).
Van Lint, S. et al. The ReNAissanCe of mRNA-based cancer therapy. Expert Rev. Vaccines 14, 235–251 (2015).
Kallen, K. J. et al. A novel, disruptive vaccination technology: self-adjuvanted RNActive® vaccines. Hum. Vaccin Immunother. 9, 2263–2276 (2013).
Rauch, S., Lutz, J., Kowalczyk, A., Schlake, T. & Heidenreich, R. RNActive® technology: generation and testing of stable and immunogenic mRNA vaccines. Methods Mol. Biol. 1499, 89–107 (2017).
Edwards, D. K. et al. Adjuvant effects of a sequence-engineered mRNA vaccine: translational profiling demonstrates similar human and murine innate response. J. Transl Med. 15, 1 (2017).
Kowalczyk, A. et al. Self-adjuvanted mRNA vaccines induce local innate immune responses that lead to a potent and boostable adaptive immunity. Vaccine 34, 3882–3893 (2016).
Schnee, M. et al. An mRNA vaccine encoding rabies virus glycoprotein induces protection against lethal infection in mice and correlates of protection in adult and newborn pigs. PLoS Negl. Trop. Dis. 10, e0004746 (2016).
Ziegler, A. et al. A new RNA-based adjuvant enhances virus-specific vaccine responses by locally triggering TLR- and RLH-dependent effects. J. Immunol. 198, 1595–1605 (2017).
Benteyn, D., Heirman, C., Bonehill, A., Thielemans, K. & Breckpot, K. mRNA-based dendritic cell vaccines. Expert Rev. Vaccines 14, 161–176 (2015).
Kranz, L. M. et al. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 534, 396–401 (2016).
Wykes, M., Pombo, A., Jenkins, C. & MacPherson, G. G. Dendritic cells interact directly with naive B lymphocytes to transfer antigen and initiate class switching in a primary T-dependent response. J. Immunol. 161, 1313–1319 (1998).
Selmi, A. et al. Uptake of synthetic naked RNA by skin-resident dendritic cells via macropinocytosis allows antigen expression and induction of T-cell responses in mice. Cancer Immunol. Immunother. 65, 1075–1083 (2016).
Diken, M. et al. Selective uptake of naked vaccine RNA by dendritic cells is driven by macropinocytosis and abrogated upon DC maturation. Gene Ther. 18, 702–708 (2011).
Lorenz, C. et al. Protein expression from exogenous mRNA: uptake by receptor-mediated endocytosis and trafficking via the lysosomal pathway. RNA Biol. 8, 627–636 (2011).
Gehl, J. Electroporation: theory and methods, perspectives for drug delivery, gene therapy and research. Acta Physiol. Scand. 177, 437–447 (2003).
Granstein, R. D., Ding, W. & Ozawa, H. Induction of anti-tumor immunity with epidermal cells pulsed with tumor-derived RNA or intradermal administration of RNA. J. Invest. Dermatol. 114, 632–636 (2000).
Kreiter, S. et al. Intranodal vaccination with naked antigen-encoding RNA elicits potent prophylactic and therapeutic antitumoral immunity. Cancer Res. 70, 9031–9040 (2010).
Bialkowski, L. et al. Intralymphatic mRNA vaccine induces CD8 T-cell responses that inhibit the growth of mucosally located tumours. Sci. Rep. 6, 22509 (2016).
Sahin, U. et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature 547, 222–226 (2017).
Qiu, P., Ziegelhoffer, P., Sun, J. & Yang, N. S. Gene gun delivery of mRNA in situ results in efficient transgene expression and genetic immunization. Gene Ther. 3, 262–268 (1996).
Steitz, J., Britten, C. M., Wolfel, T. & Tuting, T. Effective induction of anti-melanoma immunity following genetic vaccination with synthetic mRNA coding for the fusion protein EGFP.TRP2. Cancer Immunol. Immunother. 55, 246–253 (2006).
Aberle, J. H., Aberle, S. W., Kofler, R. M. & Mandl, C. W. Humoral and cellular immune response to RNA immunization with flavivirus replicons derived from tick-borne encephalitis virus. J. Virol. 79, 15107–15113 (2005).
Kofler, R. M. et al. Mimicking live flavivirus immunization with a noninfectious RNA vaccine. Proc. Natl Acad. Sci. USA 101, 1951–1956 (2004).
Mandl, C. W. et al. In vitro-synthesized infectious RNA as an attenuated live vaccine in a flavivirus model. Nat. Med. 4, 1438–1440 (1998).
Johansson, D. X., Ljungberg, K., Kakoulidou, M. & Liljestrom, P. Intradermal electroporation of naked replicon RNA elicits strong immune responses. PLoS ONE 7, e29732 (2012).
Piggott, J. M., Sheahan, B. J., Soden, D. M., O'Sullivan, G. C. & Atkins, G. J. Electroporation of RNA stimulates immunity to an encoded reporter gene in mice. Mol. Med. Rep. 2, 753–756 (2009).
Broderick, K. E. & Humeau, L. M. Electroporation-enhanced delivery of nucleic acid vaccines. Expert Rev. Vaccines 14, 195–204 (2015).
Hoerr, I., Obst, R., Rammensee, H. G. & Jung, G. In vivo application of RNA leads to induction of specific cytotoxic T lymphocytes and antibodies. Eur. J. Immunol. 30, 1–7 (2000).
Schlake, T., Thess, A., Fotin-Mleczek, M. & Kallen, K. J. Developing mRNA-vaccine technologies. RNA Biol. 9, 1319–1330 (2012).
Reichmuth, A. M., Oberli, M. A., Jeklenec, A., Langer, R. & Blankschtein, D. mRNA vaccine delivery using lipid nanoparticles. Ther. Deliv. 7, 319–334 (2016).
Midoux, P. & Pichon, C. Lipid-based mRNA vaccine delivery systems. Expert Rev. Vaccines 14, 221–234 (2015).
Kanasty, R., Dorkin, J. R., Vegas, A. & Anderson, D. Delivery materials for siRNA therapeutics. Nat. Mater. 12, 967–977 (2013).
Akinc, A. et al. Targeted delivery of RNAi therapeutics with endogenous and exogenous ligand-based mechanisms. Mol. Ther. 18, 1357–1364 (2010).
Ratajczak, M. Z. & Ratajczak, J. Horizontal transfer of RNA and proteins between cells by extracellular microvesicles: 14 years later. Clin. Transl Med. 5, 7 (2016).
Tam, H. H. et al. Sustained antigen availability during germinal center initiation enhances antibody responses to vaccination. Proc. Natl Acad. Sci. USA 113, E6639–E6648 (2016).
Richner, J. M. et al. Modified mRNA Vaccines protect against Zika virus infection. Cell 168, 1114–1125.e10 (2017).
Havenar-Daughton, C., Lee, J. H. & Crotty, S. Tfh cells and HIV bnAbs, an immunodominance model of the HIV neutralizing antibody generation problem. Immunol. Rev. 275, 49–61 (2017).
Brito, L. A. et al. Self-amplifying mRNA vaccines. Adv. Genet. 89, 179–233 (2015).
Chahal, J. S. et al. An RNA nanoparticle vaccine against Zika virus elicits antibody and CD8+ T cell responses in a mouse model. Sci. Rep. 7, 252 (2017).
Chahal, J. S. et al. Dendrimer-RNA nanoparticles generate protective immunity against lethal Ebola, H1N1 influenza, and Toxoplasma gondii challenges with a single dose. Proc. Natl Acad. Sci. USA 113, E4133–E4142 (2016).
Ulmer, J. B. & Geall, A. J. Recent innovations in mRNA vaccines. Curr. Opin. Immunol. 41, 18–22 (2016).
Alberer, M. et al. Safety and immunogenicity of a mRNA rabies vaccine in healthy adults: an open-label, non-randomised, prospective, first-in-human phase 1 clinical trial. Lancet 390, 1511–1520 (2017). This is a report of a clinical vaccine trial using directly injected, non-replicating, unmodified mRNA against an infectious pathogen.
Perri, S. et al. An alphavirus replicon particle chimera derived from venezuelan equine encephalitis and sindbis viruses is a potent gene-based vaccine delivery vector. J. Virol. 77, 10394–10403 (2003).
Fleeton, M. N. et al. Self-replicative RNA vaccines elicit protection against influenza A virus, respiratory syncytial virus, and a tickborne encephalitis virus. J. Infect. Dis. 183, 1395–1398 (2001). This is an early report of the protective efficacy that results from self-amplifying mRNA vaccines against infectious pathogens.
Magini, D. et al. Self-amplifying mRNA vaccines expressing multiple conserved influenza antigens confer protection against homologous and heterosubtypic viral challenge. PLoS ONE 11, e0161193 (2016).
Hekele, A. et al. Rapidly produced SAM® vaccine against H7N9 influenza is immunogenic in mice. Emerg. Microbes Infect. 2, e52 (2013).
Brazzoli, M. et al. Induction of broad-based immunity and protective efficacy by self-amplifying mRNA vaccines encoding influenza virus hemagglutinin. J. Virol. 90, 332–344 (2015).
Bogers, W. M. et al. Potent immune responses in rhesus macaques induced by nonviral delivery of a self-amplifying RNA vaccine expressing HIV type 1 envelope with a cationic nanoemulsion. J. Infect. Dis. 211, 947–955 (2015).
McCullough, K. C. et al. Self-replicating replicon-RNA delivery to dendritic cells by chitosan-nanoparticles for translation in vitro and in vivo. Mol. Ther. Nucleic Acids 3, e173 (2014).
Demoulins, T. et al. Polyethylenimine-based polyplex delivery of self-replicating RNA vaccines. Nanomedicine 12, 711–722 (2016).
Maruggi, G. et al. Immunogenicity and protective efficacy induced by self-amplifying mRNA vaccines encoding bacterial antigens. Vaccine 35, 361–368 (2017).
Van Gulck, E. et al. mRNA-based dendritic cell vaccination induces potent antiviral T-cell responses in HIV-1-infected patients. AIDS 26, F1–F12 (2012).
Routy, J. P. et al. Immunologic activity and safety of autologous HIV RNA-electroporated dendritic cells in HIV-1 infected patients receiving antiretroviral therapy. Clin. Immunol. 134, 140–147 (2010).
Allard, S. D. et al. A phase I/IIa immunotherapy trial of HIV-1-infected patients with Tat, Rev and Nef expressing dendritic cells followed by treatment interruption. Clin. Immunol. 142, 252–268 (2012).
Gandhi, R. T. et al. Immunization of HIV-1-infected persons with autologous dendritic cells transfected with mRNA encoding HIV-1 Gag and Nef: results of a randomized, placebo-controlled clinical trial. J. Acquir. Immune Def. Syndr. 71, 246–253 (2016).
Jacobson, J. M. et al. Dendritic cell immunotherapy for HIV-1 infection using autologous HIV-1 RNA: a randomized, double-blind, placebo-controlled clinical trial. J. Acquir. Immune Def. Syndr. 72, 31–38 (2016).
Gay, C. L. et al. Immunogenicity of AGS-004 dendritic cell therapy in patients treated during acute HIV infection. AIDS Res. Hum. Retroviruses 10.1089/aid.2017.0071 (2017).
Van Craenenbroeck, A. H. et al. Induction of cytomegalovirus-specific T cell responses in healthy volunteers and allogeneic stem cell recipients using vaccination with messenger RNA-transfected dendritic cells. Transplantation 99, 120–127 (2015).
Martinon, F. et al. Induction of virus-specific cytotoxic T lymphocytes in vivo by liposome-entrapped mRNA. Eur. J. Immunol. 23, 1719–1722 (1993). This early study demonstrates that liposome-encapsulated mRNA encoding a viral antigen induces T cell responses.
Pollard, C. et al. Type I IFN counteracts the induction of antigen-specific immune responses by lipid-based delivery of mRNA vaccines. Mol. Ther. 21, 251–259 (2013).
Zhao, M., Li, M., Zhang, Z., Gong, T. & Sun, X. Induction of HIV-1 gag specific immune responses by cationic micelles mediated delivery of gag mRNA. Drug Deliv. 23, 2596–2607 (2016).
Li, M. et al. Enhanced intranasal delivery of mRNA vaccine by overcoming the nasal epithelial barrier via intra- and paracellular pathways. J. Control. Release 228, 9–19 (2016).
Richner, J. M. et al. Vaccine mediated protection against Zika virus-induced congenital disease. Cell 170, 273–283.e12 (2017).
Roman, F., Vaman, T., Kafeja, F., Hanon, E. & Van Damme, P. AS03(A)-adjuvanted influenza A (H1N1) 2009 vaccine for adults up to 85 years of age. Clin. Infect. Dis. 51, 668–677 (2010).
Zarei, S. et al. Immunogenicity and reactogenicity of two diphtheria-tetanus-whole cell pertussis vaccines in Iranian pre-school children, a randomized controlled trial. Hum. Vaccin. Immunother. 9, 1316–1322 (2013).
Diken, M., Kranz, L. M., Kreiter, S. & Sahin, U. mRNA: a versatile molecule for cancer vaccines. Curr. Issues Mol. Biol. 22, 113–128 (2017).
Fiedler, K., Lazzaro, S., Lutz, J., Rauch, S. & Heidenreich, R. mRNA cancer vaccines. Recent Results Cancer Res. 209, 61–85 (2016).
Grunwitz, C. & Kranz, L. M. mRNA cancer vaccines-messages that prevail. Curr. Top. Microbiol. Immunol. 405, 145–164 (2017).
McNamara, M. A., Nair, S. K. & Holl, E. K. RNA-Based Vaccines in Cancer Immunotherapy. J. Immunol. Res. 2015, 794528 (2015).
Sullenger, B. A. & Nair, S. From the RNA world to the clinic. Science 352, 1417–1420 (2016).
Vigneron, N. Human tumor antigens and cancer immunotherapy. Biomed. Res. Int. 2015, 948501 (2015).
Tureci, O. et al. Targeting the heterogeneity of cancer with individualized neoepitope vaccines. Clin. Cancer Res. 22, 1885–1896 (2016).
Coulie, P. G., Van den Eynde, B. J., van der Bruggen, P. & Boon, T. Tumour antigens recognized by T lymphocytes: at the core of cancer immunotherapy. Nat. Rev. Cancer 14, 135–146 (2014).
Conry, R. M. et al. Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res. 55, 1397–1400 (1995).
Boczkowski, D., Nair, S. K., Snyder, D. & Gilboa, E. Dendritic cells pulsed with RNA are potent antigen-presenting cells in vitro and in vivo. J. Exp. Med. 184, 465–472 (1996). This report demonstrates the efficacy of mRNA DC vaccines.
De Keersmaecker, B. et al. The combination of 4-1BBL and CD40L strongly enhances the capacity of dendritic cells to stimulate HIV-specific T cell responses. J. Leukoc. Biol. 89, 989–999 (2011).
Dannull, J. et al. Enhancing the immunostimulatory function of dendritic cells by transfection with mRNA encoding OX40 ligand. Blood 105, 3206–3213 (2005).
Aerts-Toegaert, C. et al. CD83 expression on dendritic cells and T cells: correlation with effective immune responses. Eur. J. Immunol. 37, 686–695 (2007).
Grunebach, F. et al. Cotransfection of dendritic cells with RNA coding for HER-2/neu and 4-1BBL increases the induction of tumor antigen specific cytotoxic T lymphocytes. Cancer Gene Ther. 12, 749–756 (2005).
Bontkes, H. J., Kramer, D., Ruizendaal, J. J., Meijer, C. J. & Hooijberg, E. Tumor associated antigen and interleukin-12 mRNA transfected dendritic cells enhance effector function of natural killer cells and antigen specific T-cells. Clin. Immunol. 127, 375–384 (2008).
Bontkes, H. J. et al. Dendritic cells transfected with interleukin-12 and tumor-associated antigen messenger RNA induce high avidity cytotoxic T cells. Gene Ther. 14, 366–375 (2007).
Dorrie, J. et al. Introduction of functional chimeric E/L-selectin by RNA electroporation to target dendritic cells from blood to lymph nodes. Cancer Immunol. Immunother. 57, 467–477 (2008).
Bonehill, A. et al. Enhancing the T-cell stimulatory capacity of human dendritic cells by co-electroporation with CD40L, CD70 and constitutively active TLR4 encoding mRNA. Mol. Ther. 16, 1170–1180 (2008). This is a description of the TriMix mRNA adjuvant cocktail.
Van Lint, S. et al. Preclinical evaluation of TriMix and antigen mRNA-based antitumor therapy. Cancer Res. 72, 1661–1671 (2012).
Van Lint, S. et al. Optimized dendritic cell-based immunotherapy for melanoma: the TriMix-formula. Cancer Immunol. Immunother. 63, 959–967 (2014).
Pen, J. J. et al. Modulation of regulatory T cell function by monocyte-derived dendritic cells matured through electroporation with mRNA encoding CD40 ligand, constitutively active TLR4, and CD70. J. Immunol. 191, 1976–1983 (2013).
Wilgenhof, S. et al. Therapeutic vaccination with an autologous mRNA electroporated dendritic cell vaccine in patients with advanced melanoma. J. Immunother. 34, 448–456 (2011).
Wilgenhof, S. et al. A phase IB study on intravenous synthetic mRNA electroporated dendritic cell immunotherapy in pretreated advanced melanoma patients. Ann. Oncol. 24, 2686–2693 (2013).
Mitchell, D. A. et al. Tetanus toxoid and CCL3 improve dendritic cell vaccines in mice and glioblastoma patients. Nature 519, 366–369 (2015).
Batich, K. A. et al. Long-term survival in glioblastoma with cytomegalovirus pp65-targeted vaccination. Clin. Cancer Res. 23, 1898–1909 (2017).
Wilgenhof, S. et al. Phase II study of autologous monocyte-derived mRNA electroporated dendritic cells (TriMixDC-MEL) plus ipilimumab in patients with pretreated advanced melanoma. J. Clin. Oncol. 34, 1330–1338 (2016).
Zhou, W. Z. et al. RNA melanoma vaccine: induction of antitumor immunity by human glycoprotein 100 mRNA immunization. Hum. Gene Ther. 10, 2719–2724 (1999).
Kreiter, S. et al. FLT3 ligand as a molecular adjuvant for naked RNA vaccines. Methods Mol. Biol. 1428, 163–175 (2016).
Kreiter, S. et al. FLT3 ligand enhances the cancer therapeutic potency of naked RNA vaccines. Cancer Res. 71, 6132–6142 (2011).
Bol, K. F. et al. Intranodal vaccination with mRNA-optimized dendritic cells in metastatic melanoma patients. Oncoimmunology 4, e1019197 (2015).
Phua, K. K., Staats, H. F., Leong, K. W. & Nair, S. K. Intranasal mRNA nanoparticle vaccination induces prophylactic and therapeutic anti-tumor immunity. Sci. Rep. 4, 5128 (2014).
Scheel, B. et al. Therapeutic anti-tumor immunity triggered by injections of immunostimulating single-stranded RNA. Eur. J. Immunol. 36, 2807–2816 (2006).
Van der Jeught, K. et al. Intratumoral administration of mRNA encoding a fusokine consisting of IFN-β and the ectodomain of the TGF-β receptor II potentiates antitumor immunity. Oncotarget 5, 10100–10113 (2014).
Van Lint, S. et al. Intratumoral delivery of TriMix mRNA results in T-cell activation by cross-presenting dendritic cells. Cancer Immunol. Res. 4, 146–156 (2016).
Clausen, B. E. & Stoitzner, P. Functional specialization of skin dendritic cell subsets in regulating T Cell responses. Front. Immunol. 6, 534 (2015).
Weide, B. et al. Direct injection of protamine-protected mRNA: results of a phase 1/2 vaccination trial in metastatic melanoma patients. J. Immunother. 32, 498–507 (2009).
Kubler, H. et al. Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: a first-in-man phase I/IIa study. J. Immunother. Cancer 3, 26 (2015).
Hess, P. R., Boczkowski, D., Nair, S. K., Snyder, D. & Gilboa, E. Vaccination with mRNAs encoding tumor-associated antigens and granulocyte-macrophage colony-stimulating factor efficiently primes CTL responses, but is insufficient to overcome tolerance to a model tumor/self antigen. Cancer Immunol. Immunother. 55, 672–683 (2006).
Oberli, M. A. et al. Lipid nanoparticle assisted mRNA delivery for potent cancer immunotherapy. Nano Lett. 17, 1326–1335 (2017).
Fotin-Mleczek, M. et al. Highly potent mRNA based cancer vaccines represent an attractive platform for combination therapies supporting an improved therapeutic effect. J. Gene Med. 14, 428–439 (2012).
Fotin-Mleczek, M. et al. mRNA-based vaccines synergize with radiation therapy to eradicate established tumors. Radiat. Oncol. 9, 180 (2014).
Pascolo, S. Messenger RNA-based vaccines. Expert Opin. Biol. Ther. 4, 1285–1294 (2004).
Geall, A. J., Mandl, C. W. & Ulmer, J. B. RNA: the new revolution in nucleic acid vaccines. Semin. Immunol. 25, 152–159 (2013).
Weissman, D., Pardi, N., Muramatsu, H. & Kariko, K. HPLC purification of in vitro transcribed long RNA. Methods Mol. Biol. 969, 43–54 (2013).
Muralidhara, B. K. et al. Critical considerations for developing nucleic acid macromolecule based drug products. Drug Discov. Today 21, 430–444 (2016).
Jones, K. L., Drane, D. & Gowans, E. J. Long-term storage of DNA-free RNA for use in vaccine studies. Biotechniques 43, 675–681 (2007).
Probst, J. et al. Characterization of the ribonuclease activity on the skin surface. Genet. Vaccines Ther. 4, 4 (2006).
U.S. Food & Drug Administration. Guidance for Industry: Considerations for plasmid DNA vaccines for infectious disease indications. U.S. Food & Drug Administration https://www.fda.gov/downloads/BiologicsBloodVaccines/GuidanceComplianceRegulatoryInformation/Guidances/Vaccines/ucm091968.pdf (2007).
U.S. Food & Drug Administration. Guidance for Industry: Guidance for human somatic cell therapy and gene therapy. U.S. Food & Drug Administration https://www.fda.gov/downloads/BiologicsBloodVaccines/GuidanceComplianceRegulatoryInformation/Guidances/CellularandGeneTherapy/ucm081670.pdf (1998).
European Medicines Agency. Commission Directive 2009/120/EC. European Commission https://ec.europa.eu/health//sites/health/files/files/eudralex/vol-1/dir_2009_120/dir_2009_120_en.pdf (2009).
Hinz, T. et al. The European regulatory environment of RNA-based vaccines. Methods Mol. Biol. 1499, 203–222 (2017).
Pepini, T. et al. Induction of an IFN-mediated antiviral response by a self-amplifying RNA vaccine: implications for vaccine design. J. Immunol. 198, 4012–4024 (2017).
Theofilopoulos, A. N., Baccala, R., Beutler, B. & Kono, D. H. Type I interferons (α/β) in immunity and autoimmunity. Annu. Rev. Immunol. 23, 307–336 (2005).
Nestle, F. O. et al. Plasmacytoid predendritic cells initiate psoriasis through interferon-α production. J. Exp. Med. 202, 135–143 (2005).
Fischer, S. et al. Extracellular RNA mediates endothelial-cell permeability via vascular endothelial growth factor. Blood 110, 2457–2465 (2007).
Kannemeier, C. et al. Extracellular RNA constitutes a natural procoagulant cofactor in blood coagulation. Proc. Natl Acad. Sci. USA 104, 6388–6393 (2007).
Liu, M. A. & Ulmer, J. B. Human clinical trials of plasmid DNA vaccines. Adv. Genet. 55, 25–40 (2005).
DeFrancesco, L. The 'anti-hype' vaccine. Nat. Biotechnol. 35, 193–197 (2017).
Servick, K. On message. Science 355, 446–450 (2017).
CureVac AG. From science to patients — ideas become treatments at CureVac. CureVac http://www.curevac.com/research-development (2017).
Aldevron. Aldevron expands North Dakota biomanufacturing facility. Aldevron http://www.aldevron.com/about-us/news/aldevron-expands-north-dakota-biomanufacturing-facility (2016).
Kreiter, S. et al. Mutant MHC class II epitopes drive therapeutic immune responses to cancer. Nature 520, 692–696 (2015).
Ott, P. A. et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 547, 217–221 (2017).
Jacobson, E. B., Caporale, L. H. & Thorbecke, G. J. Effect of thymus cell injections on germinal center formation in lymphoid tissues of nude (thymusless) mice. Cell. Immunol. 13, 416–430 (1974).
Forster, R., Emrich, T., Kremmer, E. & Lipp, M. Expression of the G-protein-coupled receptor BLR1 defines mature, recirculating B cells and a subset of T-helper memory cells. Blood 84, 830–840 (1994).
Forster, R. et al. A putative chemokine receptor, BLR1, directs B cell migration to defined lymphoid organs and specific anatomic compartments of the spleen. Cell 87, 1037–1047 (1996).
Breitfeld, D. et al. Follicular B helper T cells express CXC chemokine receptor 5, localize to B cell follicles, and support immunoglobulin production. J. Exp. Med. 192, 1545–1552 (2000).
Schaerli, P. et al. CXC chemokine receptor 5 expression defines follicular homing T cells with B cell helper function. J. Exp. Med. 192, 1553–1562 (2000).
Johnston, R. J. et al. Bcl6 and Blimp-1 are reciprocal and antagonistic regulators of T follicular helper cell differentiation. Science 325, 1006–1010 (2009).
Nurieva, R. I. et al. Bcl6 mediates the development of T follicular helper cells. Science 325, 1001–1005 (2009).
Yu, D. et al. The transcriptional repressor Bcl-6 directs T follicular helper cell lineage commitment. Immunity 31, 457–468 (2009).
Crotty, S. A brief history of T cell help to B cells. Nat. Rev. Immunol. 15, 185–189 (2015).
Klein, F. et al. Antibodies in HIV-1 vaccine development and therapy. Science 341, 1199–1204 (2013).
Gils, A., Bertolotto, A., Mulleman, D., Bejan-Angoulvant, T. & Declerck, P. J. Biopharmaceuticals: reference products and biosimilars to treat inflammatory diseases. Ther. Drug Monit. 39, 308–315 (2017).
Sparrow, E., Friede, M., Sheikh, M. & Torvaldsen, S. Therapeutic antibodies for infectious diseases. Bull. World Health Organ. 95, 235–237 (2017).
Henricks, L. M., Schellens, J. H., Huitema, A. D. & Beijnen, J. H. The use of combinations of monoclonal antibodies in clinical oncology. Cancer Treat. Rev. 41, 859–867 (2015).
Lewiecki, E. M. Treatment of osteoporosis with denosumab. Maturitas 66, 182–186 (2010).
Paton, D. M. PCSK9 inhibitors: monoclonal antibodies for the treatment of hypercholesterolemia. Drugs Today 52, 183–192 (2016).
Hollevoet, K. & Declerck, P. J. State of play and clinical prospects of antibody gene transfer. J. Transl Med. 15, 131 (2017).
Fuchs, S. P. & Desrosiers, R. C. Promise and problems associated with the use of recombinant AAV for the delivery of anti-HIV antibodies. Mol. Ther. Methods Clin. Dev. 3, 16068 (2016).
Boczkowski, D., Lee, J., Pruitt, S. & Nair, S. Dendritic cells engineered to secrete anti-GITR antibodies are effective adjuvants to dendritic cell-based immuno-therapy. Cancer Gene Ther. 16, 900–911 (2009).
Pruitt, S. K. et al. Enhancement of anti-tumor immunity through local modulation of CTLA-4 and GITR by dendritic cells. Eur. J. Immunol. 41, 3553–3563 (2011).
Pardi, N. et al. Administration of nucleoside-modified mRNA encoding broadly neutralizing antibody protects humanized mice from HIV-1 challenge. Nat. Commun. 8, 14630 (2017). This is the first study to demonstrate that directly injected, non-replicating mRNA encoding a monoclonal antibody protects animals against an infectious pathogen.
Stadler, C. R. et al. Elimination of large tumors in mice by mRNA-encoded bispecific antibodies. Nat. Med. 23, 815–817 (2017).
Thran, M. et al. mRNA mediates passive vaccination against infectious agents, toxins, and tumors. EMBO Mol. Med. 9, 1434–1447 (2017).
Sebastian, M. et al. Phase Ib study evaluating a self-adjuvanted mRNA cancer vaccine (RNActive®) combined with local radiation as consolidation and maintenance treatment for patients with stage IV non-small cell lung cancer. BMC Cancer 14, 748 (2014).
Wang, Y. et al. Systemic delivery of modified mRNA encoding herpes simplex virus 1 thymidine kinase for targeted cancer gene therapy. Mol. Ther. 21, 358–367 (2013).
Perche, F. et al. Enhancement of dendritic cells transfection in vivo and of vaccination against B16F10 melanoma with mannosylated histidylated lipopolyplexes loaded with tumor antigen messenger RNA. Nanomedicine 7, 445–453 (2011).
Mockey, M. et al. mRNA-based cancer vaccine: prevention of B16 melanoma progression and metastasis by systemic injection of MART1 mRNA histidylated lipopolyplexes. Cancer Gene Ther. 14, 802–814 (2007).
Uchida, S. et al. Systemic delivery of messenger RNA for the treatment of pancreatic cancer using polyplex nanomicelles with a cholesterol moiety. Biomaterials 82, 221–228 (2016).
Lazzaro, S. et al. CD8 T-cell priming upon mRNA vaccination is restricted to bone-marrow-derived antigen-presenting cells and may involve antigen transfer from myocytes. Immunology 146, 312–326 (2015).
Van Driessche, A. et al. Clinical-grade manufacturing of autologous mature mRNA-electroporated dendritic cells and safety testing in acute myeloid leukemia patients in a phase I dose-escalation clinical trial. Cytotherapy 11, 653–668 (2009).
Van Tendeloo, V. F. et al. Induction of complete and molecular remissions in acute myeloid leukemia by Wilms' tumor 1 antigen-targeted dendritic cell vaccination. Proc. Natl Acad. Sci. USA 107, 13824–13829 (2010).
Berneman, Z. N. et al. Dendritic cell vaccination in malignant pleural mesothelioma: a phase I/II study [abstract]. J. Clin. Oncol. 32 (Suppl.), 7583 (2014).
Amin, A. et al. Survival with AGS-003, an autologous dendritic cell-based immunotherapy, in combination with sunitinib in unfavorable risk patients with advanced renal cell carcinoma (RCC): phase 2 study results. J. Immunother. Cancer 3, 14 (2015).
Khoury, H. J. et al. Immune responses and long-term disease recurrence status after telomerase-based dendritic cell immunotherapy in patients with acute myeloid leukemia. Cancer 123, 3061–3072 (2017).
Sebastian, M. et al. Messenger RNA vaccination in NSCLC: findings from a phase I/IIa clinical trial [abstract]. J. Clin. Oncol. 29 (Suppl.), 2584 (2011).
Rausch, S., Schwentner, C., Stenzl, A. & Bedke, J. mRNA vaccine CV9103 and CV9104 for the treatment of prostate cancer. Hum. Vaccin Immunother. 10, 3146–3152 (2014).
Mitchell, D. A. et al. Monoclonal antibody blockade of IL-2 receptor α during lymphopenia selectively depletes regulatory T cells in mice and humans. Blood 118, 3003–3012 (2011).
Borch, T. H. et al. mRNA-transfected dendritic cell vaccine in combination with metronomic cyclophosphamide as treatment for patients with advanced malignant melanoma. Oncoimmunology 5, e1207842 (2016).
Kongsted, P. et al. Dendritic cell vaccination in combination with docetaxel for patients with metastatic castration-resistant prostate cancer: a randomized phase II study. Cytotherapy 19, 500–513 (2017).
Kyte, J. A. et al. Immune response and long-term clinical outcome in advanced melanoma patients vaccinated with tumor-mRNA-transfected dendritic cells. Oncoimmunology 5, e1232237 (2016).
Vik-Mo, E. O. et al. Therapeutic vaccination against autologous cancer stem cells with mRNA-transfected dendritic cells in patients with glioblastoma. Cancer Immunol. Immunother. 62, 1499–1509 (2013).
Lesterhuis, W. J. et al. Immunogenicity of dendritic cells pulsed with CEA peptide or transfected with CEA mRNA for vaccination of colorectal cancer patients. Anticancer Res. 30, 5091–5097 (2010).
Aarntzen, E. H. et al. Vaccination with mRNA-electroporated dendritic cells induces robust tumor antigen-specific CD4+ and CD8+ T cells responses in stage III and IV melanoma patients. Clin. Cancer Res. 18, 5460–5470 (2012).
Bol, K. F. et al. Long overall survival after dendritic cell vaccination in metastatic uveal melanoma patients. Am. J. Ophthalmol. 158, 939–947 (2014).
Bol, K. F. et al. Prophylactic vaccines are potent activators of monocyte-derived dendritic cells and drive effective anti-tumor responses in melanoma patients at the cost of toxicity. Cancer Immunol. Immunother. 65, 327–339 (2016).
Weide, B. et al. Results of the first phase I/II clinical vaccination trial with direct injection of mRNA. J. Immunother. 31, 180–188 (2008).
Acknowledgements
D.W. was supported by the National Institute of Allergy and Infectious Disease (NIAID) of the US National Institutes of Health (NIH) under award numbers CHAVI-ID UM1-AI100645, R01-AI050484, R01-AI124429 and R01-AI084860, by Gates Foundation Collaboration for AIDS Vaccine Discovery (CAVD) grant OPP1033102, by the Defense Advanced Research Projects Agency under grant HR0011-17-2-0069 and by the New Frontier Sciences division of Takeda Pharmaceuticals. F.W.P. was supported by the Center for HIV/AIDS Vaccine Immunology and Immunogen Discovery (CHAVI-ID) grant UM1-AI100645, Defense Advanced Research Projects Agency grant HR0011-17-2-0069 and by Gates Foundation grant OPP1066832. The authors acknowledge K. Karikó for her profoundly helpful advice.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
In accordance with the University of Pennsylvania policies and procedures and our ethical obligations as researchers, we report that Norbert Pardi, Michael J. Hogan and Drew Weissman are named on patents that describe the use of nucleoside-modified mRNA as a platform to deliver therapeutic proteins and vaccines. We have disclosed those interests fully to the University of Pennsylvania, and we have in place an approved plan for managing any potential conflicts arising from licensing of our patents. Frederick Porter reports no competing financial interests.
Related links
FURTHER INFORMATION
Glossary
- Dendritic cell
-
(DC). A professional antigen-presenting cell that can potently activate CD4+ and CD8+ T cells by presenting peptide antigens on major histocompatibility complex (MHC) class I and II molecules, respectively, along with co-stimulatory molecules.
- Pathogen-associated molecular pattern
-
(PAMP). Conserved molecular structure produced by microorganisms and recognized as an inflammatory danger signal by various innate immune receptors.
- Type I interferon
-
A family of proteins, including but not limited to interferon-β (IFNβ) and multiple isoforms of IFNα, released by cells in response to viral infections and pathogen products. Type I IFN sensing results in the upregulation of interferon-stimulated genes and an antiviral cellular state.
- Fast protein liquid chromatography
-
(FPLC). A form of liquid chromatography that can be used to purify proteins or nucleic acids. High-performance liquid chromatography (HPLC) is a similar approach, which uses high pressure to purify materials.
- Nucleoside modification
-
The incorporation of chemically modified nucleosides, such as pseudouridine, 1-methylpseudouridine, 5-methylcytidine and others, into mRNA transcripts, usually to suppress innate immune sensing and/or to improve translation.
- Adjuvant
-
An additive to vaccines that modulates and/or boosts the potency of the immune response, often allowing lower doses of antigen to be used effectively. Adjuvants may be based on pathogen-associated molecular patterns (PAMPs) or on other molecules that activate innate immune sensors.
- MHC class I
-
A polymorphic set of proteins expressed on the surface of all nucleated cells that present antigen to CD8+ (including cytotoxic) T cells in the form of proteolytically processed peptides, typically 8–11 amino acids in length.
- MHC class II
-
A polymorphic set of proteins expressed on professional antigen-presenting cells and certain other cell types, which present antigen to CD4+ (helper) T cells in the form of proteolytically processed peptides, typically 11–30 amino acids in length.
- Good manufacturing practice
-
(GMP). A collection of guidelines and practices designed to guarantee the production of consistently high-quality and safe pharmaceutical products. GMP-grade materials must be used for human clinical trials.
- Passive immunization or passive immunotherapy
-
In contrast to traditional (active) vaccines, these therapies do not generate de novo immune responses but can provide immune-mediated protection through the delivery of antibodies or antibody-encoding genes. Passive vaccination offers the advantage of immediate action but at the disadvantage of high cost.
Rights and permissions
About this article

Cite this article
Pardi, N., Hogan, M., Porter, F. et al. mRNA vaccines — a new era in vaccinology. Nat Rev Drug Discov 17, 261–279 (2018). https://doi.org/10.1038/nrd.2017.243
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrd.2017.243
This article is cited by
-
Screening of novel tumor-associated antigens for lung adenocarcinoma mRNA vaccine development based on pyroptosis phenotype genes
BMC Cancer (2024)
-
Willingness to pay for an mRNA-based anti-cancer treatment: results from a contingent valuation study in Israel
Israel Journal of Health Policy Research (2024)
-
An overview on mRNA-based vaccines to prevent monkeypox infection
Journal of Nanobiotechnology (2024)
-
High-throughput barcoding of nanoparticles identifies cationic, degradable lipid-like materials for mRNA delivery to the lungs in female preclinical models
Nature Communications (2024)
-
An oncolytic circular RNA therapy
Nature Cancer (2024)
