
Abstract
We have previously shown that impulsivity in rats predicts the emergence of compulsive cocaine seeking and taking, and is coupled to decreased D2/3 receptor availability in the ventral striatum. As withdrawal from cocaine normalises high impulsivity in rats, we investigated, using positron emission tomography (PET), the effects of response-contingent cocaine administration on D2/3 receptor availability in the striatum. Rats were screened for impulsive behavior on the five-choice serial reaction time task. After a baseline PET scan with the D2/3 ligand [18F]fallypride, rats were trained to self-administer cocaine for 15 days under a long-access schedule. As a follow-up, rats were assessed for impulsivity and underwent a second [18F]fallypride PET scan. At baseline, we found that D2/3 receptor availability was significantly lower in the left, but not right, ventral striatum of high-impulsive rats compared with low-impulsive rats. While the number of self-administered cocaine infusions was not different between the two impulsivity groups, impulsivity selectively decreased in high-impulsive rats withdrawn from cocaine. This effect was accompanied by a significant increase in D2/3 receptor availability in the left, but not right, ventral striatum. We further report that D2/3 receptor availability was inversely related to baseline D2/3 receptor availability in the ventral striatum of high-impulsive rats, as well as to the left and right dorsal striatum of both low-impulsive and high-impulsive rats. These findings indicate that the reduction in impulsivity in high-impulsive rats by prior cocaine exposure may be mediated by a selective correction of deficient D2/3 receptor availability in the ventral striatum. A similar baseline-dependent mechanism may account for the therapeutic effects of stimulant drugs in clinical disorders such as ADHD.
Similar content being viewed by others

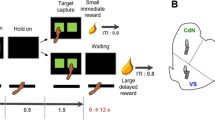
INTRODUCTION
Extreme and persistent forms of impulsive behavior are often present in addiction, hypothetically emerging from chronic substance abuse and/or as a pre-existing vulnerability trait marker (Dalley et al, 2011; Jentsch and Taylor, 1999; Verdejo-Garcia et al, 2008). The neural basis of increased impulsivity in drug addicts is poorly understood but concordant evidence implicates abnormalities in fronto-striatal ganglia circuitry, together with impaired dopamine (DA) neurotransmission in fronto-striatal networks (Koob and Volkow, 2010; Swanson et al, 2007). The most robust neural consequence of individuals being exposed to the stimulant drug cocaine, is a reduction in DA D2/3 receptors in the striatum (Lee et al, 2009; Volkow et al, 2001; Volkow et al, 1993), an effect arising most parsimoniously as a direct response to chronic drug exposure itself (Dalley et al, 2008; Groman et al, 2012; Nader et al, 2006). However, dysfunction of D2/3 receptors is also present before drug exposure in both experimental animals and humans that express high levels of impulsive behavior (Buckholtz et al, 2010; Dalley et al, 2007a). Thus, impaired D2/3 receptor signalling may underlie impulsive behavior and be a susceptibility marker that is further compromised by chronic drug abuse.
A clear prediction arising from the work above is that chronic drug exposure in impulsive subjects will produce an additional decrement in D2/3 receptor availability in the striatum, notably in the nucleus accumbens, a key brain area involved in regulating impulse control (Basar et al, 2010; Dalley et al, 2011). As D2/3 receptor availability is decreased in the ventral striatum of high-impulsive (HI) rats (Dalley et al, 2007a), exposure of impulsive animals to cocaine should further diminish D2/3 receptor availability in the ventral striatum and thereby exacerbate impulsivity. Previous research, however, found that impulsivity was strongly decreased in HI rats withdrawn from intravenous cocaine self-administration (Dalley et al, 2007a; Everitt et al, 2008) and this intervention either had no effect (Dalley et al, 2005a) or transiently increased impulsivity (Winstanley et al, 2009) in low-impulsive (LI) rats. These findings suggest that baseline differences in impulsive behavior and D2/3 receptor availability in the ventral striatum may be critical variables in shaping the ensuing effects of cocaine on response inhibitory control mechanisms. Elucidating the mechanism underlying this interaction may also be relevant to the clinical efficacy of stimulant drugs in clinical disorders of impulse control such as attention-deficit/hyperactivity disorder (ADHD). Furthermore, because ADHD represents a significant risk factor for substance use disorder, (Lee et al, 2011; van Emmerik-van Oortmerssen et al, 2012) this study has relevance for elucidating the etiological basis of vulnerability mechanisms underlying addiction. In the present study, we therefore investigated the consequences of cocaine exposure on D2/3 receptor availability in the ventral striatum in relation to behavioral impulsivity on the five-choice serial reaction time task (five-CSRTT). High impulsivity on this paradigm is measured by the number of anticipatory responses to an imminent visual signal and is analogous to false alarms on the analogous continuous performance test in humans (Robbins, 2002). We used positron emission tomography (PET) and the selective high-affinity D2/3 receptor antagonist [18F]fallypride (Mukherjee et al, 1995) to assess D2/3 receptor availability in the dorsal and ventral striatum, both before, and following exposure of rats to intravenous cocaine self-administration. In parallel, we investigated the relationship between behavioral impulsivity in selected LI and HI rats and changes in D2/3 receptor availability in the ventral striatum following the discontinuation of cocaine self-administration.
MATERIALS AND METHODS
Subjects
Ninety-six adult male Lister-hooded rats (Charles River, Margate, UK), weighing 250–275 g and 2–3 months of age at the beginning of behavioral training, were used in this study. These were housed in groups of four in enclosed ventilation chambers during the training and selection of HI and LI rats. Upon completion of the screening and for the remaining period of the study, rats were singly housed (n=10 HI; n=12 LI). One cohort of rats was assigned to the cocaine self-administration experiment (n=6 HI; n=8 LI), while a second age-matched group of rats (n=4 HI; n=4 LI) was maintained for an equivalent period of time as the first group but did not receive cocaine. The holding room was humidity- and temperature-controlled (22 °C), and rats were maintained under a reversed 12-h light/dark cycle (white lights off/red lights on at 07:00 hours). Food was restricted to maintain body weights at 85–90% of free-feeding weights. Water was available ad libitum. All experiments conformed to the UK Animals (Scientific Procedures) Act of 1986 and local ethical guidelines. A timeline of experimental procedures is shown in Figure 1.

Timeline of experimental procedures in rats expressing differential levels of impulsive behavior on the five-choice serial reaction time task (five-CSRTT). Rats were trained for ∼3 months before impulsivity screening. Values shown are in weeks.
Five-Choice Serial Reaction Time Task
The apparatus consisted of 12 five-choice chambers (25 × 25 × 25 cm; Med Associates, St Albans, VT), each enclosed in a ventilated wooden sound-attenuating cubicle and illuminated by a house-light. The front wall of the chamber was curved with five equally-spaced 2.5 cm square apertures equipped with infrared detectors and a 3W LED located at the rear. A food magazine with infrared detectors at the entrance was set in the rear wall, into which 45 mg reward pellets were delivered (Noyes dustless pellets, Research Diets, UK). The chambers were controlled by a PC using WhiskerServer software (version 2.8) and a Five-Choice client (Cardinal and Aitken, 2010).
Rats were trained on the five-CSRTT over ∼50 daily sessions (six sessions per week) to detect the location of a brief visual stimulus (0.7 s) presented on a random basis in one of the five recesses. Each session consisted of 100 discrete trials and lasted for ∼30 min. Rats were advanced through 10 stages by steadily reducing the stimulus duration (Bari et al, 2008). Training was considered complete when rats responded to the target stimuli of duration 0.7 s with an accuracy of 75% and omissions on fewer than 20% of trials. Trials were initiated by subjects entering the magazine. After a fixed inter-trial interval (ITI) of 5 s, a visual stimulus was presented in a single aperture. Rats were rewarded with a single pellet if they correctly located the position of the target stimulus (a ‘correct’ response). A failure to respond within a limited hold period of 5 s was deemed an ‘omission’ and was signalled by a 5 s time-out period and a loss of food reward on that trial. Similar feedback was given on trials where rats responded in an adjacent aperture (an ‘incorrect’ response) or before the onset of the light stimulus (a ‘premature’ response). Behavioral performance was assessed by choice accuracy (% correct responses/(correct+incorrect trials); premature responding (% premature responses/(correct+incorrect+omission trials); omissions (% omission trials/(correct+incorrect+omission trials); latency to collect food (time from nose-poke response to entering the magazine, ms); correct response latency (time to make a response in the correct aperture after the onset of the light stimulus).
Rats were selected for high impulsivity using three long-ITI challenge sessions spaced 1 week apart (Dalley et al, 2007a). Challenge sessions ended after the completion of 100 discrete trials or when 60 min had elapsed. Note that all animals included in the present study completed 100 trials within the allotted time period (both before and after cocaine). HI rats were defined as those exhibiting a level of premature responding >50% on all three long-ITI sessions. LI rats were selected from the remaining rats and typically responded prematurely in fewer than 30% of trials during the long-ITI sessions.
Intravenous Cocaine Self-Administration
Intravenous cocaine self-administration was carried out using six operant chambers (31.8 × 25.4 × 26.7 cm; Med Associates, St Albans, VT), each enclosed within a ventilated wooden sound attenuating cubicle. The chambers were equipped with two, 4 cm wide retractable levers, positioned 12 cm apart and 8 cm from the grid floor (designated ‘active’ and ‘inactive’ levers). A stimulus light (2.5 W, 24 V) was positioned 2.5 cm above each lever. The chamber was illuminated by a house light (2.5 W, 24 V) positioned on the opposite wall to the levers. The chambers were controlled by a PC using WhiskerServer software (version 2.8) and a Second Order client (Cardinal et al, 2010).
Rats were implanted with a chronic indwelling catheter (CamCaths, Cambridge, UK) in the right jugular vein under isoflurane-induced anaesthesia. The catheter was externalized through a small incision on the dorsum between the scapulae. Rats were housed singly in their home-cages for 1 week to recover from surgery. Following MRI and a baseline [18F]fallypride PET scan (see below, and Figure 1), rats were trained to acquire intravenous cocaine self-administration over 5 consecutive days under a fixed-ratio-1 schedule of reinforcement. During this phase every response on the active lever resulted in a 0.1 ml infusion of sterile saline containing 0.25 mg cocaine hydrochloride (McFarlan-Smith, Edinburgh, UK). Following each cocaine infusion the active lever was retracted for 20 s, the cue light above the lever was illuminated, and the house light was extinguished. After 20 s had elapsed, the active lever was extended into the chamber, responding on which resulted in a further infusion of cocaine. Responses on the inactive lever had no effect but were recorded. During this phase, access to cocaine was restricted to 50 infusions within each 6-h training period. Thereafter, access to cocaine was increased to 150 infusions during each 6-h session on days 6–15.
Post-Cocaine Behavioral Assessment
As a follow-up, HI and LI rats were retested on the five-CSRTT 2 weeks after the final cocaine self-administration session. This was carefully staggered to ensure that the behavioral assessment was carried out at the same time for all the rats after their last exposure to cocaine. Previous studies in our laboratory have established that behavioral performance on the five-CSRTT is severely disrupted for a period of about 7 days after the withdrawal from cocaine self-administration (Dalley et al, 2005a; Dalley et al, 2005b). We therefore assessed behavioral performance and D2/3 receptor availability 2 weeks after the last cocaine self-administration session when the acutely disruptive effects of cocaine withdrawal had dissipated thus revealing more stable and potentially longer-lasting effects on behavior and dopamine receptor function (eg, as demonstrated by (Dalley et al, 2007b). Rats were initially run for two consecutive daily sessions with a fixed ITI of 5 s. They were then challenged with a single long-ITI session (ITI=7 s) involving 100 discrete trials and a maximum duration of 60 min. Within 24 h of this assessment, rats underwent a second [18F]fallypride PET scan (see Figure 1).
Magnetic Resonance Imaging (MRI)
HI and LI rats were anesthetised with 5% isoflurane and transferred to the MRI suite where they were placed on a plastic sliding cradle equipped with atraumatic ear bars. General anesthesia was maintained via the delivery of 1.5% isoflurane through a tube in the incisor bar. Body temperature was maintained at 37 °C using a rectal thermometer. Blood oxygen saturation, heart rate and breathing rate were monitored and maintained within normal limits using a non-invasive mouseOX pulse-oximeter sensor (Starr Life Science Corp, Oakmont, PA) attached to the foot.
Rats were scanned in vivo using a 4.7T Bruker BioSpec 47/40 system (Bruker Corporation, Ettlingen, Germany) over 90 min. Parameters were chosen for optimal contrast between grey and white matter in the brain (TR/TEeff 3500/45ms, ETL 16, NEX 1, 256 × 256 × 96 FOV 38.4 × 38.4 × 1.08 mm3, isotropic resolution 150 μm3). A 72 mm birdcage resonator was used for transmission and signals were detected with a 20 mm diameter surface coil.
MRI images were segmented into grey and white matter using SPM5 (Wellcome Trust Centre of Neuroimaging, UCL, London, UK) and the SPMMouse toolbox (www.fil.ion.ucl.ac.uk/spm/). DARTEL (Ashburner and Friston, 2005) was used to create standard templates of grey and white matter and the resulting transformations were applied to the bias-corrected MR images. These transformed images were averaged to provide a high-resolution anatomical template. Regions of interest (ROI) were delineated using Analyze 8.1 (Mayo Clinic, MN). The striatum was divided into three bilateral regions of interest: posterior dorsal striatum, anterior dorsal striatum, and ventral striatum (see Figure 4a).
PET
HI and LI rats were scanned using [18F]fallypride on two occasions; before intravenous cocaine self-administration and 2 weeks after the last cocaine self-administration session, which coincided within 24 h of the assessment of impulsivity on the five-CSRTT (see Figure 1). One HI and one LI rat was scanned on each day with the order counterbalanced across the pre- and post-cocaine scanning sessions.
Rats were imaged for 3 h using a microPET Focus-220 scanner (Concorde Microsystems, Knoxville, TN). The rats were placed prone on the scanner bed and the head fixed in a custom-made plastic frame using ear bars and a bite bar. Anaesthesia and physiological measures (body temperature, heart rate, blood oxygen saturation, breathing rate) were maintained as described above. Before the injection of tracer, singles-mode transmission data were acquired for 515 s using a rotating 68Ge point source (∼20 MBq). An attenuation correction sinogram was produced from this scan and a blank scan of the same duration, with scatter correction applied. For all scans, [18F]fallypride was injected intravenously over 30 s, followed by a 15 s heparin-saline flush. The injected [18F]fallypride activity (4 to 70 MBq) was adjusted so that the total mass of labelled and unlabelled fallypride injected was 0.5 nmol/kg. Dynamic data were acquired in list-mode for a 350–650 keV energy window and a 6 ns timing window. Data were subsequently binned into sinograms for the following time frames: 6 × 10 s, 3 × 20 s, 6 × 30 s, 10 × 60 s, 10 × 120 s, 29 × 5 min. Corrections were applied for randoms, dead time, normalization, attenuation, and decay. Fourier rebinning (Defrise et al, 1997) was used to compress the 4D sinograms to 3D before reconstruction with 2D filtered back projection with a Hann window cutoff at the Nyquist frequency. The image voxel size was 0.95 × 0.95 × 0.80 mm, with an array size of 128 × 128 × 95. The reconstructed images were converted to kBq/ml using global and slice factors determined from imaging a uniform phantom filled with a [18F]fluoride solution.
For each scan, a mean PET image was manually and rigidly registered to its own MR image, and each MR image was spatially normalized to the aforementioned MR template using SyN (Avants et al, 2008; Klein et al, 2009), part of the Advanced Normalization Tools (ANTS) package. By combining the rigid and non-rigid (spatial normalization) transforms, the PET images of each scan (n=64) were aligned to the MR template. We verified that the ROI for each subject did not span the intended brain regions; thus our receptor availability measurements were not affected by individual differences in brain structure.
D2/3 receptor availability was quantified using non-displaceable binding potential (BPND) (Innis et al, 2007), determined from reference tissue modeling with the cerebellum acting as the reference region. The borders of the reference region drawn on the MR template excluded the outermost lamina of the cerebellar cortex in order to avoid any partial volume error from uptake in the Purkinje cell layer. Regional BPND was estimated using the simplified reference tissue model (SRTM) (Lammertsma and Hume, 1996), while maps of BPND (used solely for illustrative purposes) were produced using implementation of the SRTM (Gunn et al, 1997).
Statistical Analysis
Behavioral data were subjected to analysis of variance (SPSS, version 17.0, Chicago, IL) using a general linear model with significance at α=0.05. Homogeneity of variance was verified using Levene's test. For repeated-measures analyses, Mauchly’s test of sphericity was applied and the degrees of freedom corrected to more conservative values using the Huynh–Feldt epsilon for any terms involving factors, in which the sphericity assumption was violated. Differences in BPND between HI and LI rats were tested with repeated measures of analysis of variance. As previous research in our laboratory revealed asymmetry in D2/3 receptor availability in the ventral striatum of HI rats (Jupp et al, 2013) we did not correct for multiple comparisons with hemispheric side. Thus, using ex-vivo autoradiography, we observed that [3H]raclopride binding was decreased collectively in the left, but not right nucleus accumbens core and shell of HI rats compared with LI rats. Pearson product moment correlations were used to assess the strength of the association between the change in BPND ((post-cocaine−pre-cocaine)/(pre-cocaine) × 100) and baseline BPND (pre-cocaine scan). All figures show group means±1 SEM. A William’s test was used to evaluate differences between the two independent rho values in HI and LI, calculated separately for the left and right ventral striatum.
RESULTS
Impulsivity Phenotypes
The behavioral profiles of LI and HI rats on the five-CSRTT are shown in Figure 2. The percentage of premature responses for HI (n=6) and LI (n=8) rats, averaged across the three long-ITI challenge sessions, each spaced 1 week apart, was 73.8±3.1% (mean±SEM) and 23.4±2.1%, respectively. HI rats were more impulsive than LI rats regardless of the ITI being set to the training interval of 5 s (‘b1’, ‘b2’, ‘b3’, ‘b4’, P<0.001) or the challenge interval of 7 s (‘LITI’, P<0.001). With the exception of attentional accuracy, which showed a significant decrease in HI rats relative to LI rats during the long ITI sessions (main effect of group: F(1,12)=13.1, P=0.004) but not during the shorter ITI sessions (Table 1), no other behavioral variable was significantly altered in HI rats.

Screening for high impulsivity on the five-CSRTT. HI rats show an increased level of anticipatory responding compared with LI rats when the ITI is fixed at 5 s (baseline sessions: b1, b2, b3, b4) or 7 s (LITI) (LI n=8; HI n=6). HI rats are also less accurate than LI rats during long ITI sessions but show no differences in the number of omissions or latency to collect food reward. Selection of HI and LI rats involved 5 consecutive days of testing (b1, b2, LITI, b3, b4), repeated three times. The data shown (mean±SEM) are averages of three consecutive test sessions in HI and LI rats (**P<0.01, ***P<0.001 HI vs LI).
Intravenous Cocaine Self-Administration
Following MRI and baseline [18F]fallypride PET scans (see Figure 1), both groups of rats were trained to self-administer cocaine by the intravenous route (Figure 3a). Two HI rats in the present study developed excessive rates of cocaine self-administration with subsequent adverse health effects that led to their removal from the study. HI and LI rats reliably self-administered cocaine during the first 5 days, when the daily number of infusions was restricted to 50 (ie, acquisition), and during the next 10 days, when access was increased to 150 daily infusions. No significant differences were observed in cocaine self-administration between LI and HI rats. The mean±SEM number of cocaine infusions during the acquisition phase was: LI=47.02±1.2; HI=48.5±1.0, and during the long-access phase: LI=134.8±8.4; HI=140.9±3.3.

Selective correction of impulsivity in HI rats following withdrawal from intravenous cocaine self-administration. (a) Response-contingent cocaine administration in LI and HI rats under a continuous reinforcement schedule. Access to cocaine was restricted to 50 infusions during the first 5 days and 150 infusions during the subsequent 10 days (0.25 mg/infusion). Under this schedule there was no significant difference between LI and HI rats in the acquisition or maintenance of cocaine self-administration. (b) Behavioral effects of prior cocaine self-administration in LI (white bars) and HI (black bars) on the five-CSRTT. Pre-cocaine values are averaged across three weekly-spaced long ITI sessions (HI vs LI, ***P<0.001). Post-cocaine data are derived from a single long-ITI session administered 2 weeks after the last drug exposure day. It can be seen that HI rats withdrawn from cocaine show a selective reduction in premature responding compared with LI rats (†††P<0.001). There were no significant effects of prior cocaine exposure on attentional accuracy, omissions or magazine latencies in HI and LI rats. The insert graph shows a separate group of age-matched but surgically- and cocaine-naive LI and HI rats (each n=4), which were tested and retested on the five-CSRTT at precisely the same interval as those rats in the main study. It can be seen that HI rats remain impulsive on this task throughout the duration of the study (*P<0.05; **P<0.01).
Post-Cocaine Behavioral Assessment
As a follow-up, HI and LI rats were reassessed for impulsivity and attentional performance on the five-CSRTT. It can be seen in Figure 3b that high impulsivity was profoundly reduced in HI rats withdrawn from cocaine self-administration (group × cocaine interaction: F(1,12)=24.8, P<0.001, post-hoc t-test, P<0.001). This effect was specific to premature responding in HI rats with no significant effects of cocaine exposure on omissions or the speed and accuracy of responding on the five-CSRTT (Figure 3b and Table 1). Furthermore, in a separate group of age-matched cocaine-naive HI rats (n=4), we found no significant decline in impulsivity scores when rats where maintained for an equivalent period of time as those rats in the main study above (see Figure 3b insert and Table 1).
D2/3 Receptor Availability In The Ventral Striatum
Next we investigated D2/3 receptor availability in the ventral striatum of LI and HI rats, both before cocaine self-administration (‘pre-cocaine’) and again 2 weeks after the last cocaine self-administration session, (ie, beyond the acute withdrawal period) when impulsivity had significantly decreased in HI rats (‘post-cocaine’). We found a significantly lower D2/3 receptor availability in the left (t(12)=3.393, P=0.005) but not right (t(12)=0.843, P=0.416) ventral striatum of drug-naive HI rats compared with LI rats (group × hemisphere interaction: F(1,12)=4.9, P=0.04, Figure 4c). Following cocaine self-administration, this difference in D2/3 receptor availability between LI and HI rats in the left ventral striatum was no longer evident (t(12)=−0.242, P=0.813), and remained non-significant in the right hemisphere (t(12)=0.149, P=0.884). Thus, following cocaine exposure, only in the HI group did we observe a significant increase in D2/3 receptor availability in the ventral striatum (pre-/post-cocaine × −side × group interaction: F(1,12)=7.49, P<0.05; post-hoc t-test, P<0.05). The mean±SEM [18F]fallypride BPND values for the left ventral striatum in LI and HI rats, respectively, were 6.25±0.18 and 5.36±0.18 (pre-cocaine), and 6.18±0.24 and 6.28±0.39 (post-cocaine).

Selective remediation of deficient D2/3 receptor availability in the left ventral striatum of HI rats by prior exposure to intravenous cocaine self-administration. (a) 3D depiction of regions of interest showing the ventral striatum (blue), anterior dorsal striatum (green), and posterior dorsal striatum (red). (b) Horizontal section through [18F]fallypride BPND maps for HI and LI rats overlaid on the coregistered MR template (left (L) and right (R)). The images are 7 mm below the dorsal brain surface (BPND threshold=14). (c) Binding potential (BPND) of [18F]fallypride in the left and right ventral striatum of LI (square symbols, n=8) and HI (circle symbols, n=6) rats before (‘pre-cocaine’) and after (‘post-cocaine’) cocaine self-administration. It can be seen that [18F]fallypride BPND is significantly reduced in the left ventral striatum of HI rats compared with LI rats before cocaine exposure (**P<0.01) and that cocaine selectively normalises [18F]fallypride BPND in HI rats in this brain region (*P<0.05).
Baseline-Dependent Effects Of Cocaine On Striatal D2/3 Receptors
Figure 5 shows individual [18F]fallypride BPND values in the anterior and posterior dorsal striatum of LI and HI rats before and after cocaine self-administration. We found no significant group differences in D2/3 receptor availability between LI and HI rats in either region, irrespective of cocaine exposure status. However, when we compared the change in [18F]fallypride BPND before and after cocaine exposure we found that cocaine both increased and decreased [18F]fallypride BPND depending on the baseline availability of D2/3 receptors in the ventral and dorsal striatum (Figure 6). In the ventral striatum this effect was restricted to HI rats with a strong inverse relationship between the percentage change in [18F]fallypride BPND and baseline [18F]fallypride BPND in both the left (rleft=−0.88, P=0.021) and right (rright= −0.90, P=0.014) hemisphere. Visual inspection of the data indicated the presence of an obvious outlier (figure 6a, right panel). However, when this subject was removed from the regression analysis we still observed a significant inverse relationship between the percentage change in [18F]fallypride BPND and baseline [18F]fallypride BPND in the ventral striatum regardless of impulsivity group (rleft=−0.52, P=0.07; rright= −0.65, P=0.016). Moreover, individual changes in D2/3 receptors availability did not significantly correlate with individual changes in impulsive behaviour (LI rats: rleft=0.53, P=0.18; rright=0.49, P=0.21) (HI: rleft=0.31, P=0.54; rright=0.36, P=0.48). In addition, there was no significant difference between the two independent correlation coefficients for the left and right ventral striatum (William’s testleft z=1.1, P=0.86; 1.0; William’s testright z=1.0, P=0.84).

Binding potentials of [18F]fallypride in the left (a, c) and right (b, d) anterior and posterior dorsal striatum of LI (square symbols, n=8) and HI (circle symbols, n=6) rats before (‘pre-cocaine’) and after (‘post-cocaine’) cocaine self-administration. There were no significant baseline differences in [18F]fallypride BPND in either brain region between LI and HI rats. Prior cocaine exposure also had no significant effect on [18F]fallypride BPND in the anterior and posterior dorsal striatum of LI and HI rats.

Relationship between the percentage change in [18F]fallypride BPND in ventral and dorsal striatum before and after the exposure of LI and HI rats to cocaine as a function of baseline (ie, pre-cocaine) [18F]fallypride BPND. With the exception of [18F]fallypride BPND in the ventral striatum of LI rats, the results show that the effects of cocaine on D2/3 receptor availability depend in an inverse manner on baseline [18F]fallypride BPND. The horizontal dotted line depicts no net effect of cocaine on [18F]fallypride BPND. Pearson product moment correlation coefficients and P-values are given in each panel.
Post-cocaine baseline-dependent effects on D2/3 receptor availability were also observed in the anterior and posterior dorsal striatum, which in these regions extended to LI rats as well. For all regions the relationship was strongly inversely related to baseline D2/3 receptor availability (anterior dorsal striatum LI rats: rleft=−0.78, P=0.02; rright= −0.75, P=0.03, HI rats: rleft=−0.93, P=0.008; rright=−0.91, P=0.01; posterior dorsal striatum LI rats: rleft =−0.86, P=0.006, rright=−0.85, P=0.008, HI rats: rleft =−0.87, P=0.024, rright=−0.91, P=0.01).
DISCUSSION
Here we replicate and extend our previous finding of lower D2/3 receptor availability in the ventral striatum of high-impulsive rats (Dalley et al, 2007a) by demonstrating that high impulsivity in rats is selectively reduced by prior exposure to response-contingent cocaine administration and that this effect is accompanied by a selective increase in D2/3 receptor availability in the ventral striatum of high-impulsive rats compared with low-impulsive rats. Furthermore the observed decrease or increase in D2/3 receptor availability in the striatum was baseline-dependent. Thus, when the pre-exposure binding potential of [18F]fallypride was low, as was the case in the ventral striatum of high-impulsive rats, prior cocaine exposure and withdrawal had the effect of increasing D2/3 receptor availability. In contrast, when baseline binding potential of [18F]fallypride was high, we observed reduced D2/3 receptor availability, an effect that was present throughout the ventral and dorsal striatum. As high impulsivity was selectively reduced in rats with a recent prior history of cocaine self-administration, increased D2/3 receptor availability in the ventral striatum may be a contributory mechanism underlying the reduction of impulsivity in high impulsive rats.
High-Impulsivity is Associated With Lower D2/3 Receptors Availability in the Ventral Striatum and is Selectively Ameliorated by Prior Exposure to Cocaine
In line with previous results we show that low D2/3 receptor availability in the ventral striatum is a latent phenotype of high impulsivity in rats. However, in our original study (Dalley et al, 2007a), this deficit was bilaterally expressed, whereas in the present study D2/3 receptor availability was lower only in the left ventral striatum of HI rats compared with LI rats. The reason for this divergent result is unclear but may be due to methodological improvements made in the present study. Firstly, we rigidly registered PET images to individual MR images and non-rigidly registered these to a template MR image, whereas the previous approach involved rigid registration of PET to a template MR image. We also used an anatomical outline of the ventral striatum (Figure 4a) rather than a spherical region of interest and imaged the rats using a microPET Focus-220 scanner, which has a higher spatial resolution than the microPET P4 scanner used previously. Finally, we injected a higher radiotracer activity in the present study (mean 28 MBq vs 13 MBq), which coupled with the higher sensitivity of the Focus-220 compared with the P4, will have resulted in higher image signal-to-noise ratios. Collectively, these technical refinements would be expected to improve the precision and accuracy of [18F]fallypride BPND estimation in the ventral striatum, and thereby confirm that low D2/3 receptor availability in the ventral striatum is an endophenotype associated with high-impulsivity in rats.
Based on prior findings (Dalley et al, 2009; Groman et al, 2012; Nader et al, 2006), we predicted that prior cocaine exposure would further decrease D2/3 receptors in the ventral striatum and exacerbate impulsivity. However, our results clearly show that withdrawal from cocaine self-administration both improves impulse control and restores D2/3 receptor availability in this region, an effect that was strongly baseline-dependent. Although there was one obvious outlier in the high-impulsive group where baseline [18F]fallypride BPND was low (∼4), the general inverse relationship between baseline [18F]fallypride BPND and the subsequent effects of cocaine on this index held in all striatal regions-of-interest, except the ventral striatum for LI rats. Thus, the baseline-dependent effects on D2/3 receptor availability evident during withdrawal from cocaine were regionally-indiscriminate and probably therefore mediated by common global underlying mechanisms rather than by mechanisms that were specific to the ventral striatum. In considering the nature of this mechanism, it is unlikely that aging had any major bearing on these results (ie, the time elapsed between the first and second PET scan) since aging studies in both human and non-human subjects invariably report a decrease in D2/3 receptors in the striatum (Giardino, 1996; Morris et al, 1999; Volkow et al, 1996; Wong et al, 1984), not an increase as found in the present study.
Several explanations are possible to account for the observed increase in D2/3 receptor availability in the dorsal and ventral striatum of rats exposed to cocaine. Firstly, a consistent effect associated with cocaine withdrawal is an upregulation of striatal D3 receptors (Conrad et al, 2010; Neisewander et al, 2004). As [18F]fallypride binds to both D2 and D3 receptors (Mukherjee et al, 1995) it is possible that the cocaine-induced increase in [18F]fallypride BPND was caused by a selective upregulation of D3 receptors. Indeed, binding of the radiotracer [11C]-(+)-PHNO, which has preferential affinity for D3 receptors, is reportedly increased in the striatum of methamphetamine poly-drug users (Boileau et al, 2012). However, it is unclear whether the low expression of D3 receptors in the dorsal striatum compared with the ventral striatum (Sokoloff et al, 1990) would be sufficient to generate the broadly similar increases in [18F]fallypride BPND observed in both striatal regions in the present study. Secondly, withdrawal from stimulant drugs (<3 weeks) is associated with behavioral sensitization to direct-acting D2 receptor agonists (De Vries et al, 2002; Edwards et al, 2007; Robinson and Berridge, 2008; Ujike et al, 1990). This effect may reflect the reported increase in high-affinity D2 receptors in the striatum of rats withdrawn from cocaine self-administration (Briand et al, 2008). However, as fallypride is unlikely to distinguish between low- and high-affinity D2 receptor states (Seeman et al, 2003) this mechanism would appear unlikely in the present context. Thirdly, the observed increase in [18F]fallypride BPND in rats withdrawn from cocaine may reflect a reduction in DA release in the striatum.
Although post-mortem and imaging studies evaluating the dopamine transporter (DAT) in chronic cocaine abusers have been mixed and inconclusive, there has been no compelling evidence that this presynaptic marker is downregulated in cocaine addiction, putatively caused by a loss of dopaminergic terminals and/or a change in the regulation of this primary protein target (Narendran and Martinez, 2008). It is probable therefore that the downregulation of D2/3 receptors in the striatum of abstinent cocaine addicts (Martinez et al, 2004; Volkow et al, 2003) and in animals (Nader et al, 2006) reflects a compensatory response of post-synaptic D2/3 receptors to limit the effects of repeated drug-induced increases in DA release in this region. On the basis of the present findings this may be the predominant effect when the baseline pool of D2/3 receptors in the striatum is high. By contrast, and in keeping with a recent study in humans expressing high levels of impulsivity and low D2/3 receptor availability in the midbrain (Buckholtz et al, 2010), an adaptive consequence of prior cocaine exposure may be to preferentially increase the availability of presynaptic D2/3 receptors in the striatum when their density at baseline is low. If this hypothesis is correct, the present results suggest the novel inference that compensatory changes in presynaptic DA receptors are more likely when baseline D2/3 receptor availability is relatively low in the striatum.
Response-Contingent Cocaine Administration Has Persistent Effects on the Expression of Impulsive Behavior
Exposure of HI rats to cocaine had the dramatic effect of reducing their impulsive behavior on the five-choice task when subsequently tested 2 weeks after the last drug self-administration session. In addition, the accuracy of HI rats was no longer significantly different from LI rats when tested in withdrawal. We believe that these effects were caused by the exposure of rats to cocaine itself, and subsequent withdrawal, as a separate group of age-matched but drug-naive high-impulsive rats continued to show high levels of impulsivity on this task at an identical time of testing. Consistent with this conclusion, we have shown that yoked intravenous infusions of saline over several weeks have no effect on impulsive behavior (Dalley et al, 2007a; Dalley et al, 2005a). In addition, the normalizing effect of prior cocaine exposure on impulsivity was not due to differences in the quantity or rate of cocaine self-administration, which was not different between LI and HI rats. The failure to observe differential rates of cocaine escalation between the impulsivity groups, as reported previously (Dalley et al, 2007a), may be due to the continuous daily exposure of rats to cocaine self-administration in the present study, unlike our earlier study, which repeatedly exposed rats to cycles of cocaine self-administration and periods of forced withdrawal in between to assess longitudinal effects on impulsivity. Nevertheless, two high-impulsive rats in the present study developed excessive rates of cocaine self-administration with subsequent adverse health effects that led to their removal from the study. The exclusion of these animals may have influenced the conclusion of no difference in escalation between the two impulsivity groups.
The reduction in impulsivity in HI rats induced by prior cocaine exposure was behaviorally-selective with no ancillary effects on the speed or motivation to respond on the task. The neural basis of reduced impulsivity in cocaine-exposed rats remains unclear but converging evidence implicates a correction of D2/3 receptor-mediated neurotransmission in the nucleus accumbens, a key region involved in regulating impulse control (Basar et al, 2010; Dalley et al, 2011). In a recent autoradiography study, D2/3 receptors were found to be significantly reduced in the nucleus accumbens shell, but not core, of drug-naive HI rats (Jupp et al, 2013). Consistent with this evidence, impulsivity was exacerbated in HI rats following local infusions of a D2/3 receptor antagonist in the shell (Besson et al, 2010). Thus, our present PET findings suggest that cocaine pre-exposure may attenuate impulsivity in HI rats by producing a long-lasting upregulation of D2/3 receptors in the shell subregion of the nucleus accumbens. This putative mechanism appears to be restricted to HI rats because withdrawal from cocaine had no significant effect of impulsivity in LI rats. Moreover, in LI rats, cocaine did not alter D2/3 receptor availability in the ventral striatum.
Clinical Implications
Deficiencies in behavioral inhibition are present in neuropsychiatric disorders such as ADHD and addiction, and are associated with DA dysfunction (Koob et al, 2010; Monterosso et al, 2005; Swanson et al, 2007). Here we show that impaired inhibition in a rodent test of impulsivity is allied to a relative deficiency in D2/3 receptor availability in the ventral striatum, and that prior response-contingent exposure to cocaine both restores D2/3 receptor availability in this region and improves impulse control. These findings accord with reports that D2/3 receptor agonists improve response inhibitory control in stimulant addicts (Ersche et al, 2011) and that individual variation in D2/3 receptors predicts inhibitory control deficits in methamphetamine-treated monkeys (Groman et al, 2012). Our results are also broadly compatible with the rate-dependency hypothesis of stimulant drug action in ADHD, postulated to depend on baseline differences in behavior (Dews and Wenger, 1977; Robbins and Sahakian, 1979). In this study, we demonstrate evidence for baseline dependency at the neurobiological level in the striatum and this may be relevant to recent findings of reduced D2/3 receptor availability in the nucleus accumbens and caudate nucleus of unmedicated adults with ADHD (Volkow et al, 2009), as well as evidence that treatment response in this disorder is associated with increased DA transmission in the ventral striatum (Volkow et al, 2012). Thus, the clinical efficacy of stimulant drugs such as methylphenidate in ADHD may depend, in part, on restoring D2/3 receptor signalling in the ventral striatum of impulsive individuals.
References
Ashburner J, Friston KJ (2005). Unified segmentation. NeuroImage 26 (3): 839–851.
Avants BB, Epstein CL, Grossman M, Gee JC (2008). Symmetric diffeomorphic image registration with cross-correlation: evaluating automated labeling of elderly and neurodegenerative brain. Medical image analysis 12 (1): 26–41.
Bari A, Dalley JW, Robbins TW (2008). The application of the 5-choice serial reaction time task for the assessment of visual attentional processes and impulse control in rats. Nature protocols 3 (5): 759–767.
Basar K, Sesia T, Groenewegen H, Steinbusch HW, Visser-Vandewalle V, Temel Y (2010). Nucleus accumbens and impulsivity. Prog Neurobiol 92 (4): 533–557.
Besson M, Belin D, McNamara R, Theobald DE, Castel A, Beckett VL et al (2010). Dissociable control of impulsivity in rats by dopamine d2/3 receptors in the core and shell subregions of the nucleus accumbens. Neuropsychopharmacology 35 (2): 560–569.
Boileau I, Payer D, Houle S, Behzadi A, Rusjan PM, Tong J et al (2012). Higher binding of the dopamine D3 receptor-preferring ligand [11C]-(+)-propyl-hexahydro-naphtho-oxazin in methamphetamine polydrug users: a positron emission tomography study. J Neurosci 32 (4): 1353–1359.
Briand LA, Flagel SB, Seeman P, Robinson TE (2008). Cocaine self-administration produces a persistent increase in dopamine D2 High receptors. Eur Neuropsychopharmacol 18 (8): 551–556.
Buckholtz JW, Treadway MT, Cowan RL, Woodward ND, Li R, Ansari MS et al (2010). Dopaminergic network differences in human impulsivity. Science 329 (5991): 532.
Cardinal RN, Aitken MR (2010). Whisker: a client-server high-performance multimedia research control system. Behav Res Methods 42 (4): 1059–1071.
Conrad KL, Ford K, Marinelli M, Wolf ME (2010). Dopamine receptor expression and distribution dynamically change in the rat nucleus accumbens after withdrawal from cocaine self-administration. Neuroscience 169 (1): 182–194.
Dalley JW, Everitt BJ, Robbins TW (2011). Impulsivity, compulsivity, and top-down cognitive control. Neuron 69 (4): 680–694.
Dalley JW, Fryer TD, Aigbirhio FI, Brichard L, Richards HK, Hong YT et al (2009). Modelling human drug abuse and addiction with dedicated small animal positron emission tomography. Neuropharmacology 56 (Suppl 1): 9–17.
Dalley JW, Fryer TD, Brichard L, Robinson ES, Theobald DE, Laane K et al (2007a). Nucleus accumbens D2/3 receptors predict trait impulsivity and cocaine reinforcement. Science 315 (5816): 1267–1270.
Dalley JW, Laane K, Pena Y, Theobald DE, Everitt BJ, Robbins TW (2005a). Attentional and motivational deficits in rats withdrawn from intravenous self-administration of cocaine or heroin. Psychopharmacology (Berl) 182 (4): 579–587.
Dalley JW, Laane K, Theobald DE, Pena Y, Bruce CC, Huszar AC et al (2007b). Enduring deficits in sustained visual attention during withdrawal of intravenous methylenedioxymethamphetamine self-administration in rats: results from a comparative study with d-amphetamine and methamphetamine. Neuropsychopharmacology 32 (5): 1195–1206.
Dalley JW, Mar AC, Economidou D, Robbins TW (2008). Neurobehavioral mechanisms of impulsivity: fronto-striatal systems and functional neurochemistry. Pharmacol Biochem Behav 90 (2): 250–260.
Dalley JW, Theobald DE, Berry D, Milstein JA, Laane K, Everitt BJ et al (2005b). Cognitive sequelae of intravenous amphetamine self-administration in rats: evidence for selective effects on attentional performance. Neuropsychopharmacology 30 (3): 525–537.
De Vries TJ, Schoffelmeer AN, Binnekade R, Raaso H, Vanderschuren LJ (2002). Relapse to cocaine- and heroin-seeking behavior mediated by dopamine D2 receptors is time-dependent and associated with behavioral sensitization. Neuropsychopharmacology 26 (1): 18–26.
Defrise M, Kinahan PE, Townsend DW, Michel C, Sibomana M, Newport DF (1997). Exact and approximate rebinning algorithms for 3-D PET data. IEEE Trans Med Imaging 16 (2): 145–158.
Dews PB, Wenger GR (1977). Rate-dependency of the behavioral effects of amphetamine. In: Thompson T, Dews PB (eds). Advances in Behavioral Pharmacology. Academic Press, Vol 1 pp 167–227.
Edwards S, Whisler KN, Fuller DC, Orsulak PJ, Self DW (2007). Addiction-related alterations in D1 and D2 dopamine receptor behavioral responses following chronic cocaine self-administration. Neuropsychopharmacology 32 (2): 354–366.
Ersche KD, Roiser JP, Abbott S, Craig KJ, Muller U, Suckling J et al (2011). Response perseveration in stimulant dependence is associated with striatal dysfunction and can be ameliorated by a D(2/3) receptor agonist. Biol Psychiatry 70 (8): 754–762.
Everitt BJ, Belin D, Economidou D, Pelloux Y, Dalley JW, Robbins TW (2008). Review. Neural mechanisms underlying the vulnerability to develop compulsive drug-seeking habits and addiction. Philos Trans R Soc Lond B Biol Sci 363 (1507): 3125–3135.
Giardino L (1996). Right-left asymmetry of D1- and D2-receptor density is lost in the basal ganglia of old rats. Brain Res 720 (1-2): 235–238.
Groman SM, Lee B, Seu E, James AS, Feiler K, Mandelkern MA et al (2012). Dysregulation of D(2)-mediated dopamine transmission in monkeys after chronic escalating methamphetamine exposure. J Neurosci 32 (17): 5843–5852.
Gunn RN, Lammertsma AA, Hume SP, Cunningham VJ (1997). Parametric imaging of ligand-receptor binding in PET using a simplified reference region model. Neuroimage 6 (4): 279–287.
Innis RB, Cunningham VJ, Delforge J, Fujita M, Gjedde A, Gunn RN et al (2007). Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J Cereb Blood Flow Metab 27 (9): 1533–1539.
Jentsch JD, Taylor JR (1999). Impulsivity resulting from frontostriatal dysfunction in drug abuse: implications for the control of behavior by reward-related stimuli. Psychopharmacology 146 (4): 373–390.
Jupp B, Caprioli D, Saigal N, Reverte I, Shrestha S, Cumming P et al (2013). Dopaminergic and GABA-ergic markers of impulsivity in rats: evidence for anatomical localisation in ventral striatum and prefrontal cortex. Eur J Neurosci (in press).
Klein A, Andersson J, Ardekani BA, Ashburner J, Avants B, Chiang MC et al (2009). Evaluation of 14 nonlinear deformation algorithms applied to human brain MRI registration. NeuroImage 46 (3): 786–802.
Koob GF, Volkow ND (2010). Neurocircuitry of addiction. Neuropsychopharmacology 35 (1): 217–238.
Lammertsma AA, Hume SP (1996). Simplified reference tissue model for PET receptor studies. NeuroImage 4 (3 Pt 1): 153–158.
Lee B, London ED, Poldrack RA, Farahi J, Nacca A, Monterosso JR et al (2009). Striatal dopamine d2/d3 receptor availability is reduced in methamphetamine dependence and is linked to impulsivity. J Neurosci 29 (47): 14734–14740.
Lee SS, Humphreys KL, Flory K, Liu R, Glass K (2011). Prospective association of childhood attention-deficit/hyperactivity disorder (ADHD) and substance use and abuse/dependence: a meta-analytic review. Clin Psychol Rev 31 (3): 328–341.
Martinez D, Broft A, Foltin RW, Slifstein M, Hwang DR, Huang Y et al (2004). Cocaine dependence and d2 receptor availability in the functional subdivisions of the striatum: relationship with cocaine-seeking behavior. Neuropsychopharmacology 29 (6): 1190–1202.
Monterosso JR, Aron AR, Cordova X, Xu J, London ED (2005). Deficits in response inhibition associated with chronic methamphetamine abuse. Drug Alcohol Depend 79 (2): 273–277.
Morris ED, Chefer SI, Lane MA, Muzic RF, Wong DF, Dannals RF et al (1999). Loss of D2 receptor binding with age in rhesus monkeys: importance of correction for differences in striatal size. J Cereb Blood Flow Metab 19 (2): 218–229.
Mukherjee J, Yang ZY, Das MK, Brown T (1995). Fluorinated benzamide neuroleptics--III. Development of (S)-N-[(1-allyl-2-pyrrolidinyl)methyl]-5-(3-[18F]fluoropropyl)-2, 3-dimethoxybenzamide as an improved dopamine D-2 receptor tracer. Nucl Med Biol 22 (3): 283–296.
Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N et al (2006). PET imaging of dopamine D2 receptors during chronic cocaine self-administration in monkeys. Nat Neurosci 9 (8): 1050–1056.
Narendran R, Martinez D (2008). Cocaine abuse and sensitization of striatal dopamine transmission: a critical review of the preclinical and clinical imaging literature. Synapse 62 (11): 851–869.
Neisewander JL, Fuchs RA, Tran-Nguyen LT, Weber SM, Coffey GP, Joyce JN (2004). Increases in dopamine D3 receptor binding in rats receiving a cocaine challenge at various time points after cocaine self-administration: implications for cocaine-seeking behavior. Neuropsychopharmacology 29 (8): 1479–1487.
Robbins TW (2002). The 5-choice serial reaction time task: behavioural pharmacology and functional neurochemistry. Psychopharmacology (Berl) 163 (3-4): 362–380.
Robbins TW, Sahakian BJ (1979). ‘Paradoxical’ effects of psychomotor stimulant drugs in hyperactive children from the standpoint of behavioural pharmacology. Neuropharmacology 18 (12): 931–950.
Robinson TE, Berridge KC (2008). Review. The incentive sensitization theory of addiction: some current issues. Philos Trans R Soc Lond B Biol Sci 363 (1507): 3137–3146.
Seeman P, Tallerico T, Ko F (2003). Dopamine displaces [3H]domperidone from high-affinity sites of the dopamine D2 receptor, but not [3H]raclopride or [3H]spiperone in isotonic medium: Implications for human positron emission tomography. Synapse 49 (4): 209–215.
Sokoloff P, Giros B, Martres MP, Bouthenet ML, Schwartz JC (1990). Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 347 (6289): 146–151.
Swanson JM, Kinsbourne M, Nigg J, Lanphear B, Stefanatos GA, Volkow N et al (2007). Etiologic subtypes of attention-deficit/hyperactivity disorder: brain imaging, molecular genetic and environmental factors and the dopamine hypothesis. Neuropsychol Rev 17 (1): 39–59.
Ujike H, Akiyama K, Otsuki S (1990). D-2 but not D-1 dopamine agonists produce augmented behavioral response in rats after subchronic treatment with methamphetamine or cocaine. Psychopharmacology (Berl) 102 (4): 459–464.
van Emmerik-van Oortmerssen K, van de Glind G, van den Brink W, Smit F, Crunelle CL, Swets M et al (2012). Prevalence of attention-deficit hyperactivity disorder in substance use disorder patients: a meta-analysis and meta-regression analysis. Drug Alcohol Depend 122 (1-2): 11–19.
Verdejo-Garcia A, Lawrence AJ, Clark L (2008). Impulsivity as a vulnerability marker for substance-use disorders: review of findings from high-risk research, problem gamblers and genetic association studies. Neurosci Biobehav Rev 32 (4): 777–810.
Volkow ND, Chang L, Wang GJ, Fowler JS, Ding YS, Sedler M et al (2001). Low level of brain dopamine D2 receptors in methamphetamine abusers: association with metabolism in the orbitofrontal cortex. Am J Psychiatry 158 (12): 2015–2021.
Volkow ND, Fowler JS, Wang GJ (2003). The addicted human brain: insights from imaging studies. J Clin Invest 111 (10): 1444–1451.
Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ et al (1993). Decreased dopamine D2 receptor availability is associated with reduced frontal metabolism in cocaine abusers. Synapse 14 (2): 169–177.
Volkow ND, Wang G-J, Kollins SH, Wigal TL, Newcorn JH, Telang F et al (2009). Evaluating dopamine reward pathway in ADHD: clinical implications. Jama 302 (10): 1084–1091.
Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, MacGregor RR et al (1996). Measuring age-related changes in dopamine D2 receptors with 11C-raclopride and 18F-N-methylspiroperidol. Psychiatry Res 67 (1): 11–16.
Volkow ND, Wang GJ, Tomasi D, Kollins SH, Wigal TL, Newcorn JH et al (2012). Methylphenidate-elicited dopamine increases in ventral striatum are associated with long-term symptom improvement in adults with attention deficit hyperactivity disorder. J Neurosci 32 (3): 841–849.
Winstanley CA, Bachtell RK, Theobald DE, Laali S, Green TA, Kumar A et al (2009). Increased impulsivity during withdrawal from cocaine self-administration: role for DeltaFosB in the orbitofrontal cortex. Cereb Cortex 19 (2): 435–444.
Wong DF, Wagner HN, Dannals RF, Links JM, Frost JJ, Ravert HT et al (1984). Effects of age on dopamine and serotonin receptors measured by positron tomography in the living human brain. Science 226 (4681): 1393–1396.
Acknowledgements
This work was funded by the Medical Research Council (G9536855, G0701500) and by a joint award from the Medical Research Council and Wellcome Trust in support of the Behavioural and Clinical Neuroscience Institute at Cambridge University. The authors also acknowledge funding from the Medical Research Council in support of the ICCAM addiction cluster in the UK (G1000018).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
TWR discloses his consultancy with Cambridge Cognition, Lilly, Lundbeck and GlaxoSmithKline, plus research grants with Lilly, Lundbeck and GSK. The remaining authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Caprioli, D., Hong, Y., Sawiak, S. et al. Baseline-Dependent Effects of Cocaine Pre-Exposure on Impulsivity and D2/3 Receptor Availability in the Rat Striatum: Possible Relevance to the Attention-Deficit Hyperactivity Syndrome. Neuropsychopharmacol 38, 1460–1471 (2013). https://doi.org/10.1038/npp.2013.44
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2013.44
Keywords
This article is cited by
-
Serotonin and Dopamine Show Different Response Profiles to Acute Stress in the Nucleus Accumbens and Medial Prefrontal Cortex of Rats with Neuropathic Pain
Neurochemical Research (2023)
-
A sex-dependent role for the prelimbic cortex in impulsive action both before and following early cocaine abstinence
Neuropsychopharmacology (2021)
-
Dissociable effects of cocaine and yohimbine on impulsive action and relapse to cocaine seeking
Psychopharmacology (2017)
-
Attentional performance, impulsivity, and related neurotransmitter systems in apoE2, apoE3, and apoE4 female transgenic mice
Psychopharmacology (2016)
-
Noradrenergic versus dopaminergic modulation of impulsivity, attention and monitoring behaviour in rats performing the stop-signal task
Psychopharmacology (2013)
