
Abstract
This study was set to investigate whether motor effects of nociceptin/orphanin FQ (N/OFQ) can be related to changes in primary motor cortex output. N/OFQ injected i.c.v. biphasically modulated motor performance, low doses being facilitating and higher ones inhibitory. These effects were counteracted by the N/OFQ receptor antagonist [Nphe1 Arg14,Lys15]N/OFQ-NH2 (UFP-101) confirming the specificity of N/OFQ action. However, UFP-101 alone facilitated motor performance, suggesting that endogenous N/OFQ inhibits motor function. N/OFQ and UFP-101 injected into the substantia nigra reticulata but not motor cortex replicated these effects, suggesting motor responses were mediated by subcortical circuits involving the basal ganglia. Intracortical microstimulation technique showed that i.c.v. N/OFQ also biphasically modulated motor cortex excitability and movement representation. Low N/OFQ doses caused a leftward shift of threshold distribution curve in the forelimb area without affecting the number of effective sites. Conversely, high N/OFQ doses increased unresponsive and reduced excitable (movement) sites in vibrissa but not forelimb area. However, increased threshold currents and rightward shift of threshold distribution curve were observed in both areas, suggesting an overall inhibitory effect on cortical motor output. UFP-101 alone evoked effects similar to low N/OFQ doses, suggesting tonic inhibitory control over forelimb movement by endogenous N/OFQ. As shown in behavioral experiments, these effects were replicated by intranigral, but not intracortical, N/OFQ or UFP-101 injections. We conclude that N/OFQ receptors located in the substantia nigra reticulata mediate N/OFQ biphasic control over motor behavior, possibly through changes of primary motor cortex output.
Similar content being viewed by others


INTRODUCTION
Nociceptin/orphanin FQ (N/OFQ) is an endogenous neuropeptide that activates a G-protein-coupled receptor termed NOP. NOP receptors are widely represented in cortical and subcortical motor areas (Darland et al, 1998; Neal et al, 1999) and are involved in motor control. Both NOP receptor stimulation and blockade affects motor function. In particular, i.c.v. injections of N/OFQ or systemic administration of Ro 64–6198 (a synthetic NOP receptor agonist) facilitated spontaneous locomotion at low doses (Florin et al, 1996; Jenck et al, 1997; Higgins et al, 2001; Kuzmin et al, 2004) and inhibited it at higher ones (Reinscheid et al, 1995; Devine et al, 1996; Rizzi et al, 2001; Higgins et al, 2001; Kuzmin et al, 2004). NOP receptor agonists also inhibited exercise-induced locomotion (as in the rotarod test) although in a monophasic way (Jenck et al, 2000; Higgins et al, 2001; Marti et al, 2004a). In contrast, pharmacological blockade (or genetic deletion) of the NOP receptor did not affect spontaneous locomotion but increased exercise-induced motor activity (Marti et al, 2004a). Therefore, it has been proposed that endogenous N/OFQ acts as a physiological constraint of motor function, being its action more relevant under conditions of motor activation rather than at rest (Marti et al, 2004a). The neurobiological substrate(s) underlying motor actions of exogenous and endogenous N/OFQ have been investigated. The N/OFQ-induced hypolocomotion has been related to inhibition of mesencephalic mesoaccumbal (Murphy and Maidment, 1999) and/or nigrostriatal (Marti et al, 2004a) DA neurons. Consistently, injections of the NOP receptor antagonist [Nphe1 Arg14,Lys15]N/OFQ-NH2 (UFP-101) in substantia nigra reticulata (SNr) elevated rotarod performance and striatal DA release (Marti et al, 2004a). Evidence that also the hyperlocomotive response induced by i.c.v. N/OFQ is DA-dependent has been presented (Florin et al, 1996; Kuzmin et al, 2004), although the area involved has not been identified. Endogenous N/OFQ, however, appears to cause motor depressant responses also via non-DAergic mechanisms. Indeed, SNr injections of UFP-101 or systemic administration of 1-[(3R,4R)-1-cyclooctylmethyl-3-hydroxymethyl-4-piperidyl]-3-ethyl-1,3-dihydro-2H benzimidazol-2-one (J-113397) improved motor performance not only in naive but also DA-depleted (6-OHDA hemilesioned) or haloperidol-treated rats (Marti et al, 2004b, 2005). In particular, the antiakinetic effect of J-113397 in 6-OHDA hemilesioned rats was associated with reduction of nigrothalamic GABAergic transmission and, possibly, thalamic disinhibition (Marti et al, 2007). SNr is the major output nucleus of the basal ganglia. It conveys the motor information generated in the cerebral cortex and processed in the striatum to the thalamus and then back to the motor cortex, forming a functional loop which regulates movement initiation and execution (ie the so-called ‘cortico-basal ganglia-thalamo-cortical’ loop; Albin et al, 1989; Alexander and Crutcher, 1990). On this basis, pharmacological treatments (Marti et al, 2004b) or pathological conditions (Marti et al, 2005) that alter N/OFQergic transmission in SNr are likely to interfere with the activity of the ‘cortico-basal ganglia-thalamo-cortical’ loop and, as a consequence, with processing of motor information at the cortical level.
The present study was therefore undertaken to test whether changes in motor behavior produced by activation of central NOP receptors could be associated with changes in motor cortex output. Motor activity has been evaluated in awake rats by means of a battery of behavioral tests involving different motor parameters (ie time to initiate and execute a movement, coordination, and equilibrium). Primary motor cortex (M1) excitability and movement representation (defining motor output) has been investigated in anesthetized rats by intracortical microstimulation (ICMS). This technique allows to excite corticofugal neurons and produce repetitive neuronal discharges, which result in the summation of excitatory synaptic potentials in motoneurons and muscle activity. The role of central NOP receptors in modulation of motor behavior and M1 output has been investigated first by injecting N/OFQ in lateral cerebral ventricle (LCV). UFP-101 has been used to test the specificity of the N/OFQ action and to investigate the influence of endogenous N/OFQ. Finally, the role exerted by NOP receptors located in M1 and SNr has been elucidated by intracortical (layer V) or intranigral injections of N/OFQ and UFP-101.
MATERIALS AND METHODS
Male Sprague–Dawley rats (300–350 g; Stefano Morini, Reggio Emilia, Italy) were kept under regular lighting conditions (12 h light/dark cycle) and given food and water ad libitum. The experimental protocols performed in the present study were approved by Ethical Committee of the University of Ferrara and adequate measures were taken to minimize animal pain and discomfort.
Microinjection Technique
A guide cannula (outer diameter 0.55 mm, inner diameter 0.35 mm) was stereotaxically implanted under isoflurane anesthesia (1.4% in air delivered at 1.2 ml/min) 1 mm above the right or left LCV, M1 or SNr, according to the following coordinates from bregma: LCV, AP −0.9, ML ±1.4, VD −2; M1, AP +2, ML ±2, VD −0.5; SNr, AP −5.5, ML ±2.2, VD −7.3; nose bar positioned at –2.5 (Paxinos and Watson, 1982). The cannula was secured to the skull by acrylic dental cement and metallic screws. A stainless steel obturator (outer diameter 0.30 mm) was left in place inside the guide. After a 7-day recovery period, each rat was opportunely handled and trained before behavioral tests. The day of the experiment, the obturator was removed and saline or pharmacological treatments were injected (volume 0.5 μl) through a stainless-steel injector (outer diameter 0.30 mm; inner diameter 0.15 mm) protruding 1 mm from the cannula tip. At the end of each experiment the placement of the probes was verified by microscopic examination and the rats in which the probes were not correctly positioned were discarded from the study.
Behavioral Studies
Different behavioral tests were used to collect complementary information on different motor parameters in rats.
Bar test
Originally developed to quantify morphine-induced catalepsy (Kuschinsky and Hornykiewicz, 1972), this test measures the ability of the rat to respond to an externally imposed static posture. Also known as the catalepsy test (for a review see Sanberg et al, 1988), it can be used to quantify akinesia (ie time to initiate a movement) also under conditions that are not characterized by increased muscle tone (ie rigidity) as in the cataleptic/catatonic state. The rat was placed gently on a table and forepaws were placed alternatively on blocks of increasing heights (3, 6 and 9 cm). The time (in sec) that each paw spent on the block (ie the immobility time) was recorded (cutoff time of 20 s). Akinesia was calculated as total time spent on the different blocks.
Drag test
Modification of the ‘wheelbarrow test’ (Schallert et al, 1979), this test measures the ability of the rat to balance its body posture with forelimbs in response to an externally imposed dynamic stimulus (backward dragging; Marti et al, 2005). It gives information regarding the time to initiate and execute (bradykinesia) a movement. The rat was gently handled from the tail leaving the forepaws on the table, and was dragged backwards at a constant speed (about 20 cm/s) for a fixed distance (100 cm). The number of steps made by each paw were recorded. Five to seven determinations were collected for each rat.
Speed test
These tests essentially measure rat speed in an open field. The rat was allowed to habituate in a square arena (150 × 150 cm) for 5 min then elevated 3 cm about the ground (by holding its tail) and finally positioned in the centre of the arena. When the animal touched the floor it started running. Behavior was scored online using the ‘correct walking’ criteria (see Bouwman et al, 2005). Data acquisition was stopped when the rat changed its acceleration, velocity or direction. Run speed was calculated as distance traveled (cm/s).
Rotarod test
The fixed-speed rotarod test (Rozas et al, 1997) measures different motor parameters such as motor coordination, gait ability, balance, muscle tone and motivation to run. It was employed according to a previously described protocol (Marti et al, 2004a) which allowed to detect both facilitating and inhibitory drug effects. Briefly, rats were tested in a control session at 4 increasing speeds (30, 35, 40 and 45 r.p.m.; 180 s each), causing a progressive decrement of performance to ∼40% of the maximal response (ie the experimental cutoff time). Other two sessions were repeated 10 and 60 min after drug injection, and drug effect expressed as percent of control performance (total time spent on the rod).
Since pharmacological treatment may induce turning in rats, rotational behavior was measured after saline, N/OFQ or UFP-101 treatment. Rats were left to habituate in circular bowls for 20 min before the beginning of the test. Contralateral or ipsilateral turns (ie turns in the same or opposite direction to the injection side) were counted every 5 min, from 15 min before to 90 min after injection.
ICMS
Anesthesia was induced by ketamine hydrochloride (50 mg/kg i.p.) and maintained by supplementary ketamine injections (i.m.) throughout the experiment, such that long-latency, sluggish hindlimb withdrawal was achieved with severe pinching of the hind-foot. The body temperature was maintained at 36–38°C with a heat lamp. The animal was placed in a Kopf stereotaxic apparatus and a large craniotomy was performed over the frontal cortex of one side. The dura remained intact, and was kept moist with a 0.9% saline solution. Drug injections (0.01 or 10 nmol N/OFQ, 10 nmol UFP-101, dissolved in 0.5 μl of saline) were performed in the left or right LCV (AP=−0.8 mm; ML=±1.5 mm; VD=−3.5 mm below the pial surface), layer V of central M1 (AP=2–3 mm and ML=2.5–3 mm; VD=−1.5 mm) and SNr (AP=−5.5 mm; ML=±2.2 mm; VD=−7.6 mm). Drugs were slowly injected and, to prevent the substance from refluxing, the needle was withdrawn from the cortex 120 s later.
In each animal, the movements evoked by ICMS in the frontal agranular cortex were mapped starting 10 min after injection. When drugs were injected in LCV and SNr, the mapping procedure was similar to that described by Donoghue and Wise (1982) and detailed elsewhere (Franchi, 2000a). Briefly, the electrode penetrations were regularly spaced out over a 500 μm grid. Glass-insulated tungsten microelectrodes (0.6–1.2 MΩ impedance at 1 kHz), were used for stimulation. The electrode was lowered perpendicularly to the cortical surface down to layer V of the frontal agranular cortex (−1.5 mm; Franchi, 2000a). Monophasic cathodal pulses (200 μs duration, 30 ms trains at 300 Hz) of a maximum of 60 μA were passed through the electrode with a minimum interval of 2.5 s. Two observers were required for movement detection and threshold determination. Starting with a current of 60 μA, intensity was decreased in 5 μA steps until the movement was no longer evoked; then the intensity was increased to a level at which ∼50% of the stimulation elicited movement. This level defined current threshold. If no movements or twitches were evoked with 60 μA, the site was recorded as negative. When a movement was observed in two or more body parts, current thresholds were determined for each component. Body parts activated by ICMS were identified by visual inspection and/or muscle palpation. The terms ‘forelimb movement’ and ‘hindlimb movement’ refer collectively to proximal and distal joint movements. When drugs were injected in M1 the procedure was as above, but electrode penetrations were orthogonally spaced out over 200, 400 and 800 μm from injection site. Although the direct current spread is confined to 250 μm from the stimulating electrode, the current can transynaptically activate a wider area of cortex (Jankowska et al, 1975). Then, the ICMS applied at different distances to the site of injection ensures a thorough analysis of drug effect. For each set of experiments (injections in LCV, M1, SNr) appropriate controls were run in parallel: five rats were mapped with the cortex untouched (control group) and the other five received a 0.5 μl injection of saline on the correspondent site (sham group).
At the end of the experimental procedures, animals were perfused transcardially with saline and then with 4% paraformaldehyde solution. Brains were removed, postfixed, sectioned coronally into 50 μm thick slices and then stained with thionine to verify needle and microelectrode positions.
Data Presentation and Statistical Analysis
Motor performance in the bar, drag and speed tests was expressed in absolute values (sec, number of steps, cm/s, respectively) whereas motor performance in the rotarod test as percentage±SEM of the control session. Statistical analysis was performed by one-way repeated measure (RM) ANOVA followed by contrast analysis to determine group differences. In case a significant time × treatment interaction was found, the sequentially rejective Bonferroni test was used to determine specific differences (ie at the single time-point level) between groups. Drug interaction was studied experimentally according to a 2 × 2 factorial design and data analyzed with conventional two-way ANOVA, factor one being N/OFQ and factor two UFP-101. ICMS data were presented as mean±SEM. Inter-group comparisons were determined using one-way ANOVA and χ2 test presented in a two-way contingency table (2 × 2). P-values <0.05 were considered to be statistically significant.
Drugs
N/OFQ and UFP-101 were prepared at the University of Ferrara, as previously described (Guerrini et al, 2000).
RESULTS
Behavioral Studies
I.c.v. injections of nop receptor ligands
To investigate the role of central NOP receptors in modulation of motor activity, N/OFQ was injected i.c.v. (in LCV). UFP-101 was also administered to test the specificity of N/OFQ action and to investigate the role of endogenous N/OFQ. Saline injections did not affect motor activity. Indeed, immobility time, number of steps, speed and rotarod performance were similar in saline-injected (0.6±0.2 s, 13.2±0.3, 67.4±2.1 cm/s and 1021±27 s, respectively) and control (0.8±0.3 s, 12.9±0.8, 68.6±6.3 cm/s and 1064±18 s, respectively) rats. As i.c.v. injection of saline, N/OFQ and UFP-101 did not induce forepaw motor asymmetry, results obtained at the contralateral and the ipsilateral forepaw in the bar and drag test were pooled together.
N/OFQ
-
i)
Bar test: RM ANOVA on the immobility time in the bar test (Figure 1a ) showed a significant effect of treatment (F4,28=503.91, p<0.0001) time (F1,4=24.85, p<0.0001) and a significant time × treatment interaction (F4,29=11.86, p<0.0001). Post hoc analysis revealed that 0.1 nmol N/OFQ evoked a modest and transient elevation of immobility time (ie caused akinesia) compared to saline while 1 and 10 nmol N/OFQ evoked a more robust and prolonged response, detectable 60 min after injection. No change in immobility time was elicited by 0.01 nmol N/OFQ.
-
ii)
Drag test: RM ANOVA on the number of steps in the drag test (Figure 1b) showed a significant effect of treatment (F4,28=102.07, p<0.0001), time (F1,4=59.13, p<0.0001) and a significant time × treatment interaction (F4,29=23.60, p<0.0001). Post hoc analysis revealed that N/OFQ evoked a biphasic response, namely facilitation at 0.01 nmol and inhibition at higher doses (0.1–10 nmol). Both facilitation and inhibition were detected after 60 min.
Figure 1 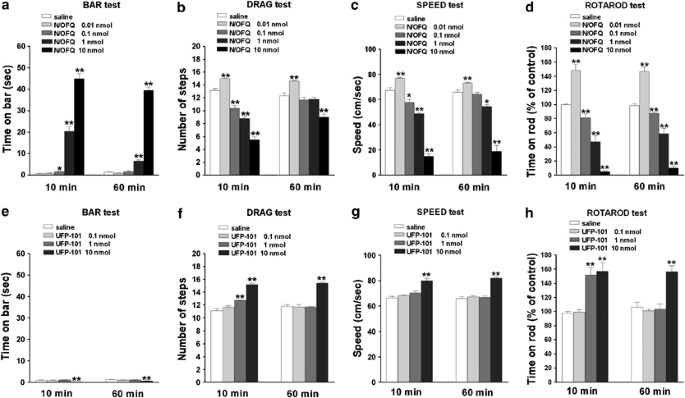
Effect of i.c.v. injection of N/OFQ or UFP-101 on motor activity. N/OFQ (0.01–10 nmol) or UFP-101 (0.1–10 nmol) were injected in the lateral cerebral ventricle and motor activity evaluated in the bar (panels a, e), drag (panels b, f), speed (panels c, g) and rotarod (panels d, h) tests. Each experiment consisted of three different sessions: a control session followed by other two sessions performed 10 and 60 min after saline, N/OFQ or UFP-101 injection (see Materials and Methods). In the bar, drag and speed test data are expressed as absolute values (sec, steps, and cm/s, respectively) whereas in the rotarod test as percentages of motor activity in the control session. Data are means±SEM of 7–9 determinations per group. *p<0.05 and **p<0.01 significantly different from saline.
-
iii)
Speed test: RM ANOVA on speed values (Figure 1c) showed a significant effect of treatment (F4,28=182.51, p<0.0001), time (F1,4=7.15, p=0.012) and a significant time × treatment interaction (F4,29=5.18, p=0.002).Post hoc analysis revealed that, as in the drag test, N/OFQ evoked a biphasic response, improving speed at 0.01 nmol and inhibiting it at higher doses (0.1–10 nmol). Both effects were long-lasting.
-
iv)
Rotarod test: RM ANOVA on rotarod values (Figure 1d) showed a significant effect of treatment (F4,28=4980.92, p<0.0001) but not time (F1,4=0.005, p=0.94) and a non significant time × treatment interaction (F4,29=0.61, p=0.66). Post hoc analysis revealed that N/OFQ improved rotarod performance at 0.01 nmol and impaired it at higher doses.

UFP-101
-
i)
Bar test: RM ANOVA showed a significant effect of treatment (F3,18=8.94, p=0.0007), time (F1,3=6.96, p=0.014) and a non significant time × treatment interaction (F3,24=0.94, p=0.43). UFP-101 caused a reduction of immobility time at 10 nmol (Figure 1e).
-
ii)
Drag test: RM ANOVA showed a significant effect of treatment (F3,18=69.29, p<0.0001) but not time (F1,3=0.01, p=0.93) and a significant time × treatment interaction (F3,24=4.69, p=0.01). UFP-101 elevated the number of steps at 10 nmol (Figure 1f). The effect of 10 nmol UFP-110 was also detected 60 min after injection.
-
iii)
Speed test: RM ANOVA showed a significant effect of treatment (F3,18=81.03, p<0.0001), but not time (F1,3=0.69, p=0.41) and a non significant time × treatment interaction (F3,24=1.63, p=0.20). UFP-101 induced long-lasting increase in speed at 10 nmol (Figure 1g).
-
iv)
Rotarod test: RM ANOVA showed a significant effect of treatment (F3,18=18.70, p<0.0001), time (F1,3=4.80, p=0.022) and a significant time × treatment interaction (F3,24=11.71, p<0.0001). UFP-101 improved the rotarod performance at 1 and 10 nmol (Figure 1h). The effect of 1 nmol UFP-101 was transient whereas that produced by the higher dose was prolonged.
Co-injection of N/OFQ and UFP-101
To investigate the selectivity of N/OFQ action, co-injections of low and high N/OFQ and UFP-101 doses (1:10 ratio) were performed. We first tested the specificity of 0.01 nmol N/OFQ by challenging it with 0.1 nmol UFP-101 (Figure 2).

Effect of i.c.v. co-injection of low doses of N/OFQ and UFP-101 on motor activity. N/OFQ (0.01 nmol) and UFP-101 (0.1 nmol) were co-injected in the lateral cerebral ventricle and motor activity evaluated in the drag (panel a), speed (panel b) and rotarod (panel c) test. Each experiment consisted of three different sessions: a control session followed by other two sessions performed 10 and 60 min after saline, N/OFQ or UFP-101 injection (see Materials and Methods). In the drag and speed test data are expressed as absolute values (steps, and cm/s, respectively) whereas in the rotarod test as percentages of motor activity in the control session. Data are means±SEM of seven determinations per group. **p<0.01 significantly different from saline. §§p<0.01 significantly different from N/OFQ.
RM ANOVA in the drag test (Figure 2a), showed a significant effect of treatment (F3,18=57.87, p<0.0001), but not time (F1,3=1.15, p=0.29) and a non significant time × treatment interaction (F3,24=0.04, p=0.98). N/OFQ elevated the number of steps and UFP-101, ineffective alone, prevented this effect. RM ANOVA on speed values (Figure 2b) revealed a significant effect of treatment (F3,18=19.34, p<0.0001) and time (F1,3=6.45, p=0.017) but not a significant interaction between the two (F3,24=1.44, p=0.25). N/OFQ increased rat speed and UFP-101, ineffective alone, prevented this effect. RM ANOVA on rotarod values (Figure 2c) revealed a significant effect of treatment (F3,18=66.09, p<0.0001), but not time (F1,3=0.02, p=0.88) and a non significant time × treatment interaction (F3,24=0.22, p=0.87). N/OFQ elevated rotarod performance while UFP-101, ineffective alone, prevented N/OFQ action.
We then tested the specificity of high N/OFQ doses by challenging 1 nmol N/OFQ with 10 nmol UFP-101 (Figure 3). RM ANOVA on the bar test (Figure 3a) revealed a significant effect of treatment (F3,18=276.13, p<0.0001), time (F1,3=27.19, p<0.0001) and a significant time × treatment interaction (F3,24=34.45, p<0.0001). N/OFQ elevated immobility time while UFP-101, ineffective alone, prevented this effect. In the drag test (Figure 3b), a significant effect of treatment (F3,18=107,13, p<0.0001), time (F1,3=12.33, p<0.0017) and a significant time × treatment interaction (F3,24=9.14, p=0.0003) were found. N/OFQ reduced the number of steps while UFP-101 increased it. The combination of the two was the sum of their effects, that is, no change with respect to saline-treated animals. In the speed test (Figure 3c), a significant effect of treatment (F3,18=94.71, p<0.0001), but not time (F1,3=1.47, p=0.23) and a non significant time × treatment interaction (F3,24=1.77, p=0.18) were observed. N/OFQ reduced, whereas UFP-101 increased speed. Again, combination of the two caused no change in speed when compared to saline-treated animals. Finally, RM ANOVA on the rotarod (Figure 3d) showed a significant effect of treatment (F3,18=164.02, p<0.0001), but not time (F1,3=0.14, p=0.70) and a non significant time × treatment interaction (F3,24=0.04, p=0.97). N/OFQ reduced rotarod performance and UFP-101 improved it. Co-application of N/OFQ and UFP-101 caused a slight increase in performance compared to saline-treated rats.

Effect of i.c.v. co-injection of high doses of N/OFQ and UFP-101 on motor activity. N/OFQ (1 nmol) and UFP-101 (10 nmol) were co-injected in the lateral cerebral ventricle and motor activity evaluated in the bar (panel a), drag (panel b), speed (panel c) and rotarod (panel d) test. Each experiment consisted of three different sessions: a control session followed by other two sessions performed 10 and 60 min after saline or UFP-101 injection (see Materials and Methods). In the bar, drag and speed test data are expressed as absolute values (sec, steps, and cm/s, respectively) whereas in the rotarod test as percentages of motor activity in the first session. Data are means±SEM of seven determinations per group. *p<0.05, **p<0.01 significantly different from saline. §p<0.05, §§p<0.01 significantly different from N/OFQ. #p<0.05, ##p<0.01 significantly different from UFP-101.
Turning behavior: LCV injection of N/OFQ or UFP-101 did not induce turning behavior in the range of doses tested.
M1 injections of NOP receptor ligands
To investigate the localization of NOP receptors involved in motor actions elicited by i.c.v. N/OFQ and UFP-101, intracortical injections (layer V of M1) were first made. Saline, N/OFQ (0.01–10 nmol) or UFP-101 (10 nmol) failed to affect rat performance in the bar, drag, speed and rotarod test (Figure S1).
SNr injections of nop receptor ligands
On the basis of our previous finding that NOP receptors located in the SNr modulate rotarod performance in rats (Marti et al, 2004a) we investigated whether motor effects induced by i.c.v. N/OFQ and UFP-101 could be reproduced by SNr injections. Since injections were made unilaterally, motor activity was evaluated separately at the ipsilateral and contralateral paw.
N/OFQ
-
i)
Bar test: Saline did not affect the immobility time at the contralateral (0.9±0.3 s) and ipsilateral (1.0±0.3 s) forepaw compared to control rats (1.1±0.3 s). RM ANOVA on the immobility time at the contralateral paw in the bar test (Figure 4a and b ) showed a significant effect of treatment (F4,24=80.13, p<0.0001), time (F1,4=11.36, p=0.002) and a significant time × treatment interaction (F4,30=3.36, p=0.021). N/OFQ increased the immobility time dose-dependently and in a prolonged way, being active yet at 0.1 nmol. Qualitatively similar data were obtained at the ipsilateral paw.
-
ii)
Drag test: Saline did not modify the number of steps made by the contralateral (11.9±0.4) and ipsilateral (12.3±0.6) paw compared to control (11.6±0.5) rats. In the drag test (Figure 4c and d), RM ANOVA at the contralateral paw disclosed a significant effect of treatment (F4,24=92.03, p<0.0001), time (F1,4=11.77, p=0.0017) and a significant time × treatment interaction (F4,30=5.16, p=0.0027). N/OFQ elevated the number of steps at 0.01 nmol but reduced them in the 0.1–10 nmol range. Conversely, RM ANOVA at the ipsilateral paw showed a significant effect of treatment (F4,24=26.35, p<0.0001), but not time (F1,4=0.27, p=0.60) and a non significant time × treatment interaction (F4,30=0.48, p=0.88). N/OFQ dose-dependently reduced the number of steps, the threshold inhibitory dose being 1 nmol.
Figure 4 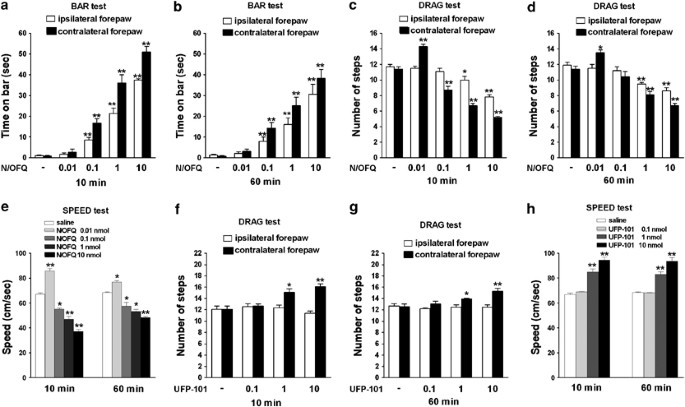
Effect of intranigral injections of N/OFQ or UFP-101 on motor activity. N/OFQ (0.01–10 nmol) or UFP-101 (0.1–10 nmol) were injected in SNr and motor activity evaluated in the bar (panels a–b), drag (panels c–d, f–g) and speed (panels e–h) test. Motor activity in the bar and drag test was evaluated separately at the paws ipsilateral and contralateral to the injection side. Motor activity in the speed test was calculated as distance traveled. Each experiment consisted of three different sessions: a control session followed by other two sessions performed 10 and 60 min after saline, N/OFQ or UFP-101 injection (see Materials and Methods). In the bar, drag and speed test data are expressed as absolute values (sec, steps, and cm/s, respectively) whereas in the rotarod test as percentages of motor activity in the control session. Data are means±SEM of seven determinations per group. *p<0.05 and **p<0.01 significantly different from saline.
-
iii)
Speed test: Saline did not affect rat speed (66.7±1.1 and 68.6±6.3 cm/s, respectively, for the saline-treated and control rats). Analysis of speed values (Figure 4e) revealed a significant effect of treatment (F4,24=136.15, p<0.0001), time (F1,4=6.35, p=0.017) and a significant time × treatment interaction (F4,30=12.37, p<0.0001). N/OFQ biphasically modulated rat speed, low doses (0.01 nmol) being facilitatory and higher ones (0.1–10 nmol) inhibitory.

UFP-101
-
i)
Bar test: RM ANOVA on the immobility time did not reveal significant effects of UFP-101 at the contralateral and ipsilateral paws (Figure S2).
-
ii)
Drag test: RM ANOVA at the contralateral paw (Figure 4f and g) revealed a significant effect of treatment (F3,18=30.22, p<0.0001) but not time (F1,3=1.38, p=0.25) and a non significant time × treatment interaction (F3,24=0.66, p=0.58). UFP-101 (1 and 10 nmol) elevated the number of steps at both 10 and 60 min post injection time. Conversely, RM ANOVA did not reveal any effect of UFP-101 on stepping activity at the ipsilateral paw.
-
iii)
Speed test: RM ANOVA on speed values (Figure 4h) showed a significant effect of treatment (F3,18=58.87, p<0.0001), but not time (F1,4=0.48, p=0.49) and a non significant time × treatment interaction (F4,30=0.95, p=0.43). UFP-101 (1 and 10 nmol) consistently elevated speed at 10 and 60 min after injection.
Co-injection of N/OFQ and UFP-101
To investigate the selectivity of N/OFQ action in SNr, co-injections of low and high N/OFQ and UFP-101 doses (1:10 ratio) were performed (Figure 5).

Effect of intranigral co-injection of N/OFQ and UFP-101 on motor activity. N/OFQ and UFP-101 were co-injected at low (0.01 and 0.1 nmol, respectively; panels a–b) and high (1 and 10 nmol, respectively; panels c–e) doses in substantia nigra pars reticulata (SNr) and motor activity evaluated, at 10 min post-injection time, in the drag (panel a, d), speed (panels b, e) and bar (panel c) test. Motor activity in the bar and drag test was evaluated separately at the paws ipsilateral and contralateral to the injection side. Motor activity in the speed test was calculated as distance traveled. In the bar, drag and speed test data are expressed as absolute values (sec, steps, and cm/s, respectively) and are means±SEM of seven determinations per group. *p<0.05, **p<0.01 significantly different from saline. §§p<0.01 significantly different from N/OFQ.
In the drag test (Figure 5a), conventional two-way ANOVA showed a main effect of N/OFQ (F1,24=14.72, p=0.0008), UFP-101 (F1,24=22.00, p=0.0001) and a significant N/OFQ × UFP-101 interaction (F1,24=11.63, p=0.0023). N/OFQ (0.01 nmol) elevated the number of steps and UFP-101 (0.1 nmol), ineffective alone, prevented this increase. Likewise, in the speed test (Figure 5b), ANOVA showed a main effect of N/OFQ (F1,24=32.45, p<0.0001), UFP-101 (F1,24=21.72, p<0.0001) and a significant N/OFQ × UFP-101 interaction (F1,24=33.71, p<0.0001). N/OFQ elevated rat speed, and UFP-101, ineffective alone, prevented its effect.
UFP-101 (10 nmol) was also challenged against N/OFQ (1 nmol). In the bar test (Figure 5c), ANOVA showed a main effect of N/OFQ (F1,24=82.79, p<0.0001), UFP-101 (F1,24=83.78, p<0.0001) and a significant N/OFQ × UFP-101 interaction (F1,24=80.45, p<0.0001) at the contralateral paw. N/OFQ increased immobility time and UFP-101, ineffective alone, prevented this effect. Similar results were obtained at the ipsilateral paw. In the drag test (Figure 5d), ANOVA revealed a main effect of N/OFQ (F1,24=140.84, p<0.0001), UFP-101 (F1,24=128.00, p<0.0001) and a significant N/OFQ × UFP-101 interaction (F1,24=11.57, p=0.0023) at the contralateral paw. N/OFQ reduced the number of steps, UFP-101 increased it and the combination of the two did not result in significant changes compared to saline-treated animals. At the ipsilateral paw, a main effect of N/OFQ (F1,24=7.91, p=0.0096) but not UFP-101 (F1,24=1.01, p=0.32) and a non significant N/OFQ × UFP-101 interaction (F1,24=2.59, p=0.12) were found. N/OFQ reduced the number of steps and UFP-101, ineffective alone, prevented this effect. Finally, ANOVA on speed values (Figure 5e) showed a main effect of N/OFQ (F1,24=22.63, p<0.0001), UFP-101 (F1,24=68.86, p=0.0001) and a non significant N/OFQ × UFP-101 interaction (F1,24=4.03, p=0.056). N/OFQ reduced speed, UFP-101 increased it and their combination resulted in a stimulation not different from that evoked by UFP-101 alone.
ICMS
Since NOP receptor stimulation or blockade affected motor activity, the hypothesis was tested that manipulation of central NOP receptors could change output from M1.
I.c.v. injections of NOP receptor ligands
Examination of M1 maps (examples are given in Figure 6) revealed several changes in movement representation in the 10 nmol N/OFQ group (Figure 6e). Contiguous unresponsive sites were consistently observed within M1 after i.c.v. injection of 10 nmol N/OFQ. To quantitatively assess these changes, the percentage of both unresponsive and responsive sites (movement sites in the vibrissa and forelimb areas) was calculated within the total site population (Figure 7a). ANOVA revealed changes in movement representation after injection of NOP receptor ligands (F14,74=10.45, p<0.0001). N/OFQ 10 nmol doubled the percentage of unresponsive sites. This effect was associated with a significant decrease (∼49%) in movement sites in the vibrissa representation and no change in excitable sites in forelimb representation. ANOVA revealed that NOP receptor ligands significantly affected movement thresholds (F9,49=31.40, p<0.0001; Figure 7b). N/OFQ (10 nmol) increased threshold currents in both vibrissa and forelimb representations (∼55 and ∼47%, respectively) whereas both N/OFQ (0.01 nmol) and UFP-101 (10 nmol) reduced them (∼17 and ∼33%), although only in forelimb representation. The differences in excitability appeared in more detail by looking at the distribution of vibrissa and forelimb movement thresholds. N/OFQ (10 nmol) caused a significant increase in the percentage of those sites where higher currents were necessary to evoke vibrissa (Figure 7c; χ2=59.37 p<0.01) and forelimb (Figure 7d; χ2=40.84, p<0.01) movements (2 × 2, 35 μA as dividing point). Conversely, N/OFQ (0.01 nmol) and UFP-101 (10 nmol) did not change the distribution of thresholds in the vibrissa but caused a significant leftward shift of the distribution curve in the forelimb representation (N/OFQ 0.01 nmol: χ2=21.26, p<0.01; UFP-101 10 nmol: χ2=41.67 p<0.01, 2 × 2, 20 μA as a dividing point). In ∼40% of sites, currents lower than 20 μA were usually necessary to evoke forelimb movement.

Effect of i.c.v. injection of N/OFQ and UFP-101 on primary motor cortex output. Representative primary motor cortex maps of movements evoked at threshold current levels in the vibrissa and forelimb areas. A schematic of rat brain showing vibrissa and forelimb areas (right) and reporting a coordinate grid (left) is represented (panel a). The maps relative to control rats (panel b) and rats injected with saline (panel c), N/OFQ (0.01 nmol; panel d), N/OFQ (10 nmol; panel e) and UFP-101 (10 nmol; panel f) in the lateral cerebral ventricle are also shown. The microelectrode was sequentially introduced to a depth of 1500 μm. Interpenetration distances were 500 μm. In these mapping schemes, frontal poles are at the bottom. Zero corresponds to bregma; numbers indicate rostral or caudal distance from the bregma or lateral distance from the mid-line. Movement evoked at one point is indicated by symbols and threshold range by the different grey scale. Absence of symbol (within or at the border of the maps) indicates that penetration was not performed due to presence of a large vessel. In panel e, the presence of dark symbols and unresponsive sites is worth noting.

Effect of i.c.v. injection of N/OFQ and UFP-101 on primary motor cortex output. N/OFQ (0.01 and 10 nmol) and UFP-101 (10 nmol) were injected in the lateral cerebral ventricle, and the percentage of unresponsive and excitable sites in the vibrissa and forelimb areas (panel a) or average thresholds currents required to evoke vibrissa and forelimb movements (panel b) were measured. Threshold current distribution is also shown (panels c–d). The percentage of other movement sites (neck, jaw, eye and hindlimb) are not shown because these movements were not extensively explored. Note that N/OFQ (10 nmol) significantly shifted to the right both vibrissa and forelimb threshold distributions whereas N/OFQ (0.01 nmol) and UFP-101 (10 nmol) significantly shifted to the left the forelimb threshold distribution. Data are means±SEM of five determinations per group. *p<0.05, **p<0.01 significantly different from control.
M1 injections of NOP receptor ligands
To investigate whether NOP receptors located in M1 modulated local excitability, injections of NOP receptor ligands in the layer V of M1 were made (examples are given in Figure 8). ANOVA on threshold currents considered as a whole or at each level away from the injection site (Figure S3) revealed no significant changes in all treated group compared to saline.

Intracortical injections of N/OFQ and UFP-101. Examples of cross-shaped grids showing injection and stimulation sites in control rats (panel b) or rats injected with saline (panel c), N/OFQ (0.01 and 10 nmol; panels d and e), and UFP-101 (10 nmol; panel f) in primary motor cortex. A schematic of rat brain showing vibrissa and forelimb areas (right) and reporting a coordinate grid (left) is also represented (panel a). For each stimulation site, a letter indicates the type of ICMS-evoked movement and the corresponding number the threshold current (in μA) required to evoke it. Note that threshold values in control and saline groups overlap with those of N/OFQ and UFP-101 groups.
SNr injections of NOP receptor ligands
Intranigral injections of NOP receptor ligands were performed to investigate whether NOP receptors located in SNr affected motor excitability. ANOVA on M1 maps derived in SNr (representative examples given in Figure 9) revealed that NOP receptor ligands modulated the numbers of responsive and unresponsive sites (F14,74=12.09, p<0.0001; Figure 10a). N/OFQ (10 nmol) was ineffective in the vibrissa area but increased (∼121%) the number of unresponsive sites and simultaneously reduced (∼60%) the number of excitable sites in the forelimb representation. ANOVA also revealed that NOP receptor ligands modulated threshold currents (F9,49=13.71, p<0.0001; Figure 10b). N/OFQ (10 nmol) enhanced the mean threshold values in the vibrissa (∼29%) and forelimb (∼58%) areas. Moreover, UFP-101 (10 nmol) reduced (∼44%) threshold currents selectively in the forelimb. A slight inhibition (∼15%) was also observed with 0.01 nmol N/OFQ in the forelimb area, which however, did not reach the level of significance. Statistical analysis on threshold distribution showed that N/OFQ 10 nmol shifted to the right the distribution in both vibrissa (Figure 10c, χ2=20.99, p<0.01) and forelimb (Figure 10d, χ2=53.27, p<0.01) evoked-movement (2 × 2, 35 μA as a dividing point). Conversely, N/OFQ (0.01 nmol) and UFP-101 (10 nmol) caused a significant leftward shift in the threshold distribution in forelimb representation (N/OFQ 0.01 nmol χ2=9.89 p<0.01; UFP-101 10 nmol: χ2=62.80 p<0.01, 2 × 2, 20 μA as a dividing point).

Effect of intranigral injection of N/OFQ and UFP-101 on primary motor cortex output. Representative primary motor cortex maps of movements evoked at threshold current levels in the vibrissa and forelimb areas. A schematic of rat brain showing vibrissa and forelimb areas (right) and reporting a coordinate grid (left) is represented (panel a). The maps relative to control rats (panel b) and rats injected with saline (panel c), N/OFQ (0.01 nmol; panel d), N/OFQ (10 nmol; panel e) and UFP-101 (10 nmol; panel f) in substantia nigra reticulata are also shown. The microelectrode was sequentially introduced to a depth of 1500 μm. Interpenetration distances were 500 μm. In these mapping schemes, frontal poles are at the bottom. Zero corresponds to bregma; numbers indicate rostral or caudal distance from the bregma or lateral distance from the mid-line. Movement evoked at one point is indicated by symbols and threshold range by the different grey scale. Absence of symbol (within or at the border of the maps) indicates that penetration was not performed due to presence of a large vessel. In panel e, the presence of dark symbols and unresponsive sites is worth noting.

Effect of intranigral injection of N/OFQ and UFP-101 on primary motor cortex output. N/OFQ (0.01 and 10 nmol) and UFP-101 (10 nmol) were injected in substantia nigra reticulata, and the percentage of unresponsive and excitable sites in the vibrissa and forelimb areas (panel a) or the thresholds currents required to evoke vibrissa and forelimb movements (panel b) were measured. Threshold current distribution is also shown (panels c–d). The percentage of other movement sites (neck, jaw, eye and hindlimb) are not shown because these movements were not extensively explored. Note that N/OFQ (10 nmol) significantly shifted to the right both vibrissa and forelimb distributions whereas N/OFQ (0.01 nmol) and UFP-101 (10 nmol) significantly shifted to the left the forelimb distribution. Data are means±SEM of five determinations per group. *p<0.05, **p<0.01 significantly different from control.
DISCUSSION
Exogenous N/OFQ produced a dose-dependent, biphasic regulation of motor performance in rats. Inhibition was predominant since it was quantitatively larger and detected in a wider dose-range than facilitation. Conversely, UFP-101 monotonically facilitated motor activity suggesting an inhibitory role for endogenous N/OFQ in motor control. NOP receptor ligands produced changes in M1 output, which were consistent with their motor effects. Thus, exogenous N/OFQ biphasically regulated motor cortex excitability, low doses being facilitatory and higher ones inhibitory. Conversely, UFP-101 increased motor cortex excitability (in the forelimb area), suggesting that endogenous N/OFQ tonically inhibits forelimb movement. Both behavioral and electrophysiological effects were evoked by i.c.v. and intranigral, but not intracortical, drug injections, overall suggesting that subcortical NOP receptors regulate motor behavior and motor cortex output via modulation of cortical afferents.
Motor impairment has been consistently reported as one of the main biological effects induced by central NOP receptor stimulation in rodents. N/OFQ given i.c.v. (Reinscheid et al, 1995; Devine et al, 1996; Nishi et al, 1997; Rizzi et al, 2001; Higgins et al, 2001; Kuzmin et al, 2004) or Ro 64–6198 given systemically (Jenck et al, 2000; Higgins et al, 2001; Varty et al, 2005) depressed both spontaneous and exercise-induced locomotion. Fairly high doses of N/OFQ (1–30 nmol) or Ro 64–6198 (10 mg/Kg) were required to depress motor activity. These doses, although selective for the NOP receptor (Nishi et al, 1997; Noda et al, 1998; Higgins et al, 2001), may induce hypolocomotion and catalepsy by affecting not only motor but also vestibular and cardiovascular functions (Sulaiman et al, 1999; Kapusta et al, 1999). The battery of complementary behavioral tests used in the present study provides information on the state of activation of the basal ganglia-thalamo-cortical circuit. Indeed, modulation of the time to initiate and to execute a movement, as in the bar and drag test, primarily engage the basal ganglia (Hauber, 1998). Walking activity (as on the rotarod) also engages the dorsolateral striatum, as shown by biochemical (Brown and Sharp, 1995; Holschneider et al, 2003) and neurochemical (Bergquist et al, 2003; Petzinger et al, 2007) evidence. Although the cerebellar-thalamo-cortical circuit is also activated during rotarod performance, its role in motor control predominates (over the basal ganglia-thalamo-cortical circuit) when training period is prolonged to several weeks (Holschneider et al, 2007).
These tests allowed to clearly demonstrate that lower doses of i.c.v. N/OFQ (0.1–1 nmol) inhibited motor behavior by inducing akinesia and bradykinesia, by slowing the time to initiate and to execute a movement. The specificity of these effects is also confirmed by reports that these doses of N/OFQ did not affect other motor parameters such as righting reflex (Devine et al, 1996) or muscle strength (Jenck et al, 1997) and tone (Devine et al, 1996; Marti et al, 2004a). Moreover, akinesia and bradykinesia, were replicated by N/OFQ injections in SNr, the motor output of basal ganglia. This confirms and extends our previous studies in naive (Marti et al, 2004a) and 6-OHDA hemilesioned (Marti et al, 2005) rats, further endorsing the view that nigral NOP receptors are modulators of specific motor patterns.
Inhibition of locomotion was not the only effect induced by central NOP receptor stimulation since very low doses of N/OFQ (0.01 nmol) produced mild but significant facilitation. In previous studies, i.c.v. injection of 0.01–0.5 nmol N/OFQ (Florin et al, 1996; Jenck et al, 1997; Kuzmin et al, 2004; Sakoori and Murphy, 2004) or systemic administration of intermediate doses of Ro 64–6198 (3–6 mg/Kg, Higgins et al, 2001) facilitated spontaneous locomotion in rodents. This facilitation was previously related to the well-known anxiolytic effect of NOP receptor agonists (Jenck et al, 1997). However, the present study points out that the 0.01 nmol N/OFQ-induced facilitation is a specific motor effect. Indeed, i.c.v. N/OFQ enhanced not only rat speed and rotarod performance but also stepping activity. Moreover, facilitation in the drag and speed test was replicated by stimulation of NOP receptors in the SNr, suggesting activation of motor pathways. To confirm this view, the contralateral limb was selectively affected in the drag test. In fact, we have previously reported that SNr injections of 0.01 nmol N/OFQ did not affect rotarod performance (Marti et al, 2004a). The most parsimonious explanation is that the improvements in stepping activity and run speed induced by unilateral SNr N/OFQ injections are too mild to affect exercise-induced locomotion as in the rotarod test.
Despite the fact that exogenous N/OFQ evoked both motor facilitation and inhibition, the latter effect appeared predominant. This is in line with the finding that endogenous N/OFQ physiologically inhibits movement. Indeed, UFP-101, given i.c.v. or injected in SNr, facilitated stepping activity, run speed and rotarod performance (see Marti et al, 2004a, for the effect of UFP-101 injections in SNr on rotarod performance). Consistently, deletion of the NOP receptor gene resulted in enhanced rotarod performance (Marti et al, 2004a). It is noteworthy that motor activation was induced by doses of UFP-101 (1 nmol i.c.v.) that were found ineffective in modulating spontaneous locomotion (Kuzmin et al, 2004; Gavioli et al, 2003; Rizzi et al, 2007; Sakoori and Murphy, 2008), pain (Calò et al, 2002, 2005; Rizzi et al, 2006) or depression (Gavioli et al, 2003). This finding strengthens the view that exercise-induced movement is the most sensitive biological parameter influenced by endogenous N/OFQ, possibly due to phasic release of N/OFQ under motor activation (Marti et al, 2005). This view is not contradicted by the finding that UFP-101 10 nmol improved motor performance also under static conditions (ie reduced the immobility time in the bar test). Indeed, this facilitation was not consistent across groups. This may possibly be due to experimental reasons. Indeed, changes in immobility time below 1 s (which is about the time required to withdraw the paw from the blocks) approach the limits of sensitivity of the method (ie the reaction time of the operator). It is therefore possible that slight changes in basal activity across different groups of animals may alter the possibility to detect a significant response to UFP-101. Overall, the data obtained with N/OFQ and UFP-101 suggest that exogenous N/OFQ is capable of activating facilitatory and inhibitory motor pathways while endogenous N/OFQ tonically interacts only with the inhibitory ones. Whether these effects are mediated by different receptor subtypes (Marti et al, 2003, Kuzmin et al, 2004) or receptor located along different and functional opposing pathways remains a matter of conjecture.
Changes in motor behavior observed after stimulation and blockade of NOP receptors in awake rats were in line with changes in motor output observed in anesthetized rats by using ICMS in layer V of M1. Indeed, the efferent neurons located in this area are most intensively involved in movement control (Beloozerova et al, 2003). ICMS in layer V elicits movement via direct stimulation of corticofugal and/or intracortical neurons (Jankowska et al, 1975), resulting in summation of excitatory synaptic potentials in motoneurons and muscle activity. Thus, movement representation, as assessed by ICMS, is a measure of the output function of the motor cortex. Moreover, it has proven to be highly sensitive to a variety of neural manipulations that influence the balance between excitatory and inhibitory circuits within M1 (Sanes et al, 1990; Hess and Donoghue, 1994; Huntley, 1997; Nudo and Milliken, 1996; Franchi, 2000b).
NOP receptors are widely expressed in cortical areas (particularly in layers III–VI; Neal et al, 1999), where they can modulate local neurotransmission both at the pre-synaptic (Sbrenna et al, 2000; Marti et al, 2003) and post-synaptic (Siniscalchi et al, 2002; Bianchi et al, 2004) level. However, neither motor output nor behavior was affected by M1 injections of NOP receptor ligands. This indicates that cortical NOP receptors were not involved in local modulation of the normal balance between excitatory and inhibitory circuits or that they did not change the excitability of cortico-fugal neurons belonging to the main subcortical output systems, namely the cortico-spinal, the cortico-pontine, the cortico-striatal and the cortico-thalamic system. Conversely, the fact that SNr injections of N/OFQ and UFP-101 affect motor cortex excitability suggests that the main influence on M1 cortical circuits is due to changes in cortical inputs. M1 receives inputs related to locomotion primarily from the ventrolateral thalamus, and in the absence of this input, the locomotion-related modulation of cortical activity nearly vanishes (Beloozerova and Sirota, 1998). Therefore, a candidate mechanism capable of altering the cortical excitability after injection of NOP receptor ligands might be the modulation of thalamic excitatory input to M1.
Neurobiological Substrates of N/OFQ Actions
Changes in DA transmission may underlie motor effects induced by N/OFQ. Indeed, stimulation of NOP receptors expressed on nigral DA neurons (Norton et al, 2002; Maidment et al, 2002) hyperpolarized DA cells and reduced their firing activity (Marti et al, 2004a). Moreover, motor inhibition induced by intranigral injections of N/OFQ was associated with reduced nigrostriatal DA transmission in vivo (Marti et al, 2004a). Disruption of motor cortex activity was also associated with motor impairment. M1 receives inputs from basal ganglia circuits that are known to be severely disrupted by striatal DA deficiency (Steiner and Kitai, 2000; Orieux et al, 2002; Parr-Brownlie and Hyland, 2005). In haloperidol-treated cats, the activity of the motor thalamus was found to be reduced and the afferent pathways to M1, that influence the segmental apparatus of the spinal cord, inhibited (Voloshin et al, 1994). Moreover, haloperidol-induced motor impairment in rats was associated with reduced baseline firing rate, bursting activity and movement-related firing in cortical neurons (Parr-Brownlie and Hyland, 2005). The finding that high N/OFQ doses injected in SNr inhibited motor behavior and cortical motor output, possibly via inhibition of DA transmission, is consistent with an inhibitory effect of N/OFQ on thalamo-cortical transmission. Indeed, reduction of thalamo-cortical inputs leaves the motor cortex functionally deactivated (Wichmann and DeLong, 1993; Obeso et al, 2000; Boraud et al, 2002; Rolland et al, 2007). Evidence that endogenous N/OFQ in SNr also modulates thalamo-cortical projections, although possibly via non-DA mechanisms, has been obtained in the 6-hydroxydopamine hemilesioned rat model of Parkinson's disease. Indeed, NOP receptor antagonists elevated GABA and reduced GLU release in the lesioned SNr, which was associated with reduced nigro-thalamic GABA transmission and attenuation of akinesia at the parkinsonian limb (Marti et al, 2007). This finding suggests that endogenous N/OFQ in SNr tonically inhibits thalamic activity. Interestingly, the electrophysiological data indicate that this tonic activity affects forelimb but not vibrissa motor representations. The main reason of this difference may be the nature of vibrissa and forelimb motor systems which involve different cortico-basal ganglia motor circuits (Hoover et al, 2003; Miyachi et al, 2006).
It proves more difficult to explain the enhanced motor cortex excitability and motor facilitation induced by low N/OFQ doses. Motor facilitation was prevented by D1 and D2 receptor antagonists (Florin et al, 1996) or catecholamine depletion (Kuzmin et al, 2004). It is possible that a low degree of NOP receptor stimulation reduces dendritic DA release in SN. This would remove the inhibitory feedback mediated by somatodendritic D2 autoreceptors (Cragg and Greenfield, 1997; Bustos et al, 2004) and result in a facilitation of rat locomotion (Bergquist et al, 2003). Alternatively, as shown for classical opioids (Johnson and North, 1992), low N/OFQ doses may preferentially inhibit GABA interneurons leading to disinhibition of nigral DA neurons (Cobb and Abercrombie, 2002).
Concluding Remarks
A careful analysis of motor behavior using a battery of complementary tests, has demonstrated that exogenous N/OFQ dose-dependently facilitates and inhibits motor behavior while endogenous N/OFQ regulates movement in an inhibitory way. Although we cannot exclude the possibility that other areas also mediate N/OFQ motor effects (eg the VTA and spontaneous locomotion), these data indicate that NOP receptors in SNr mediate specific motor programs such as time to initiate and execute a movement. The present study also demonstrates for the first time that exogenous NOP receptor ligands and endogenous N/OFQ regulate motor cortex excitability in a way, which is consistent with their motor actions. Although the neurobiological substrates remain to be investigated, the present study suggests that changes in cortical output and behavior are mainly operated by subcortical NOP receptors located in SNr through modulation of the ‘cortico-basal ganglia-thalamo-cortical’ loop.
References
Albin RA, Young AB, Penney B (1989). The functional anatomy of the basal ganglia disorders. Trends Neurosci 12: 366–375.
Alexander GE, Crutcher MD (1990). Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends Neurosci 13: 266–271.
Beloozerova IN, Sirota MG (1998). Cortically controlled gait modifications in the cat. Ann NY Acad Sci 860: 550–554.
Beloozerova IN, Sirota MG, Swadlow HA (2003). Activity of different classes of neurons of the motor cortex during locomotion. J Neurosci 23: 1087–1097.
Bergquist F, Shahabi HN, Nissbrandt H (2003). Somatodendritic dopamine release in rat substantia nigra influences motor performance on the accelerating rod. Brain Res 973: 81–91.
Bianchi C, Marani L, Barbieri M, Marino S, Beani L, Siniscalchi A (2004). Effects of nociceptin/orphanin FQ and endomorphin-1 on glutamate and GABA release, intracellular [Ca2+] and cell excitability in primary cultures of rat cortical neurons. Neuropharmacology 47: 873–883.
Boraud T, Bezard E, Bioulac B, Gross CE (2002). From single extracellular unit recording in experimental and human Parkinsonism to the development of a functional concept of the role played by the basal ganglia in motor control. Prog Neurobiol 66: 265–283.
Bouwman BM, van Lier H, Nitert HE, Drinkenburg WH, Coenen AM, van Rijn CM (2005). The relationship between hippocampal EEG theta activity and locomotor behaviour in freely moving rats: effects of vigabatrin. Brain Res Bull 64: 505–509.
Brown LL, Sharp FR (1995). Metabolic mapping of rat striatum: somatotopic organization of sensorimotor activity. Brain Res 686: 207–222.
Bustos G, Abarca J, Campusano J, Bustos V, Noriega V, Aliaga E (2004). Functional interactions between somatodendritic dopamine release, glutamate receptors and brain-derived neurotrophic factor expression in mesencephalic structures of the brain. Brain Res Brain Res Rev 47: 126–144.
Calò G, Guerrini R, Rizzi A, Salvadori S, Burmeister M, Kapusta DR et al (2005). UFP-101, a peptide antagonist selective for the nociceptin/orphanin FQ receptor. CNS Drug Rev 11: 97–112.
Calò G, Rizzi A, Rizzi D, Bigoni R, Guerrini R, Marzola G et al (2002). [Nphe1,Arg14,Lys15]Nociceptin-NH2, a novel potent and selective antagonist of the nociceptin/orphanin FQ receptor. Br J Pharmacol 136: 303–311.
Cobb WS, Abercrombie ED (2002). Distinct roles for nigral GABA and glutamate receptors in the regulation of dendritic dopamine release under normal conditions and in response to systemic haloperidol. J Neurosci 22: 1407–1413.
Cragg SJ, Greenfield SA (1997). Differential autoreceptor control of somatodendritic and axon terminal dopamine release in substantia nigra, ventral tegmental area, and striatum. J Neurosci 17: 5738–5746.
Darland T, Heinricher MM, Grandy DY (1998). Orphanin FQ/nociceptin: a role in pain and analgesia, but so much more. Trends Neurosci 21: 215–221.
Devine DP, Taylor L, Reinscheid RK, Monsma Jr FJ, Civelli O, Akil H (1996). Rats rapidly develop tolerance to the locomotor-inhibiting effects of the novel neuropeptide orphanin FQ. Neurochem Res 21: 1387–1396.
Donoghue JP, Wise SP (1982). The motor cortex of the rat: cytoarchitecture and microstimulation mapping. J Comp Neurol 212: 76–88.
Florin S, Suaudeau C, Meunier JC, Costentin J (1996). Nociceptin stimulates locomotion and exploratory behaviour in mice. Eur J Pharmacol 317: 9–13.
Franchi G (2000a). Reorganization of vibrissal motor representation following severing and repair of the facial nerve in adult rats. Exp Brain Res 131: 33–43.
Franchi G (2000b). Changes in motor representation related to facial nerve damage and regeneration in adult rats. Exp Brain Res 135: 53–65.
Gavioli EC, Marzola G, Guerrini R, Bertorelli R, Zucchini S, De Lima TC et al (2003). Blockade of nociceptin/orphanin FQ-NOP receptor signalling produces antidepressant-like effects: pharmacological and genetic evidences from the mouse forced swimming test. Eur J Neurosci 17: 1987–1990.
Guerrini R, Calò G, Bigoni R, Rizzi A, Varani K, Toth G et al (2000). Further studies on nociceptin-related peptides: discovery of a new chemical template with antagonist activity on the nociceptin receptor. J Med Chem 43: 2805–2813.
Hauber W (1998). Involvement of basal ganglia transmitter systems in movement initiation. Prog Neurobiol 56: 507–540.
Hess G, Donoghue JP (1994). Long-term potentiation of horizontal connections provides a mechanism to reorganize cortical motor maps. J Neurophysiol 71: 2543–2547.
Higgins G, Grottick AJ, Ballard TM, Richards JG, Messer J, Takeshima H et al (2001). Influence of the selective ORL1 receptor agonist, Ro64–6198, on rodent neurological function. Neuropharmacology 41: 97–107.
Holschneider DP, Maarek JM, Yang J, Harimoto J, Scremin OU (2003). Functional brain mapping in freely moving rats during treadmill walking. J Cereb Blood Flow Metab 23: 925–932.
Holschneider DP, Yang J, Guo Y, Maarek JM (2007). Reorganization of functional brain maps after exercise training: Importance of cerebellar-thalamic-cortical pathway. Brain Res 1184: 96–107.
Hoover JE, Hoffer ZS, Alloway KD (2003). Projections from primary somatosensory cortex to the neostriatum: the role of somatotopic continuity in corticostriatal convergence. J Neurophysiol 89: 1576–1587.
Huntley GW (1997). Correlation between patterns of horizontal connectivity and the extend of short-term representational plasticity in rat motor cortex. Cereb Cortex 7: 143–156.
Jankowska E, Padel Y, Tanaka R (1975). The mode of activation of pyramidal tract cells by intracortical stimuli. J Physiol 249: 617–636.
Jenck F, Moreau J, Martin J, Kilpatrick G, Reinscheid RK, Monsma Jr FJ et al (1997). Orphanin FQ acts as an anxiolytic to attenuate behavioural responses to stress. Proc Natl Acad Sci USA 94: 14854–14858.
Jenck F, Wichmann J, Dautzenberg FM, Moreau JL, Ouagazzal AM, Martin JR et al (2000). A synthetic agonist at the orphanin FQ/nociceptin receptor ORL1: anxiolytic profile in the rat. Proc Natl Acad Sci USA 97: 4938–4943.
Johnson SW, North RA (1992). Two types of neurone in the rat ventral tegmental area and their synaptic inputs. J Physiol 450: 455–468.
Kapusta DR, Chang JK, Kenigs VA (1999). Central administration of [Phe1psi(CH2-NH)Gly2]nociceptin(1-13)-NH2 and orphanin FQ/nociceptin (OFQ/N) produce similar cardiovascular and renal responses in conscious rats. J Pharmacol Exp Ther 289: 173–180.
Kuschinsky K, Hornykiewicz O (1972). Morphine catalepsy in the rat: relation to striatal dopamine metabolism. Eur J Pharmacol 19: 119–122.
Kuzmin A, Sandin J, Terenius L, Ogren SO (2004). Evidence in locomotion test for the functional heterogeneity of ORL-1 receptors. Br J Pharmacol 141: 132–140.
Maidment NT, Chen Y, Tan AM, Murphy NP, Leslie FM (2002). Rat ventral midbrain dopamine neurons express the orphanin FQ/nociceptin receptor ORL-1. Neuroreport 13: 1137–1140.
Marti M, Mela F, Fantin M, Zucchini S, Brown JM, Witta J et al (2005). Blockade of nociceptin/orphanin FQ transmission attenuates symptoms and neurodegeneration associated with Parkinson's disease. J Neurosci 95: 9591–9601.
Marti M, Mela F, Guerrini R, Calò G, Bianchi C, Morari M (2004b). Blockade of nociceptin/orphanin FQ transmission in rat substantia nigra reverses haloperidol-induced akinesia and normalizes nigral glutamate release. J Neurochem 91: 1501–1504.
Marti M, Mela F, Veronesi C, Guerrini R, Salvatori S, Federici M et al (2004a). Blockade of nociceptin/orphanin FQ receptor signalling in rat substantia nigra pars reticulata stimulates nigrostriatal dopaminergic transmission and motor behaviour. J Neurosci 24: 6659–6666.
Marti M, Stocchi S, Paganini F, Mela F, De Risi C, Calò G et al (2003). Pharmacological profiles of presynaptic nociceptin/orphanin FQ receptors modulatine 5-hydroxytryptamine and noradrenaline release in the rat neocortex. Br J Pharmacol 138: 91–98.
Marti M, Trapella C, Viaro R, Morari M (2007). The nociceptin/orphanin FQ receptor antagonist J-113397 and L-DOPA additively attenuate experimental parkinsonism through overinhibition of the nigrothalamic pathway. J Neurosci 27: 1297–1307.
Miyachi S, Lu X, Imanishi M, Sawada K, Nambu A, Takada M (2006). Somatotopically arranged inputs from putamen and subthalamic nucleus to primary motor cortex. Neurosci Res 56: 300–308.
Murphy NP, Maidment NT (1999). Orphanin FQ/nociceptin modulation of mesolimbic dopamine transmission determined by microdialysis. J Neurochem 73: 179–186.
Neal Jr CR, Mansour A, Reinscheid R, Nothacker HP, Civelli O, Akil H et al (1999). Opioid receptor-like (ORL1) receptor distribution in the rat central nervous system: comparison of ORL1 receptor mRNA expression with 125I-[14Tyr]-orphanin FQ binding. J Comp Neurol 412: 563–605.
Nishi M, Houtani T, Noda Y, Mamiya T, Sato K, Doi T et al (1997). Unrestrained nociceptive response and dysregulation of hearing ability in mice lacking the nociceptin/orphaninFQ receptor. EMBO J 16: 1858–1864.
Noda Y, Mamiya T, Nabeshima T, Nishi M, Higashioka M, Takeshima H (1998). Loss of antinociception induced by naloxone benzoylhydrazone in nociceptin receptor-knockout mice. J Biol Chem 273: 18047–18051.
Norton CS, Neal CR, Kumar S, Akil H, Watson SJ (2002). Nociceptin/orphanin FQ and opioid receptor-like receptor mRNA expression in dopamine systems. J Comp Neurol 444: 358–368.
Nudo RJ, Milliken GW (1996). Reorganization of movement representations in primary motor cortex following focal ischemic infarcts in adult squirrel monkeys. J Neurophysiol 75: 2144–2149.
Obeso JA, Rodriguez-Oroz MC, Chana P, Lera G, Rodriguez M, Olanow CW (2000). The evolution and origin of motor complications in Parkinson's disease. Neurology 5: S13–S20.
Orieux G, François C, Féger J, Hirsch EC (2002). Consequences of dopaminergic denervation on the metabolic activity of the cortical neurons projecting to the subthalamic nucleus in the rat. J Neurosci 22: 8762–8770.
Parr-Brownlie LC, Hyland BI (2005). Bradykinesia induced by dopamine D2 receptor blockade is associated with reduced motor cortex activity in the rat. J Neurosci 25: 5700–5709.
Paxinos G, Watson C (1982). The Rat Brain in Stereotaxic Coordinates. Academic: Sydney.
Petzinger GM, Walsh JP, Akopian G, Hogg E, Abernathy A, Arevalo P et al (2007). Effects of treadmill exercise on dopaminergic transmission in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse model of basal ganglia injury. J Neurosci 27: 5291–5300.
Reinscheid RK, Nothacker HP, Bourson A, Ardati A, Henningsen RA, Bunzow JR et al (1995). Orphanin FQ: a neuropeptide that activates an opioid-like G protein-coupled receptor. Science 270: 792–794.
Rizzi A, Bigoni R, Marzola G, Guerrini R, Salvadori S, Regoli D et al (2001). Characterization of the locomotor activity-inhibiting effect of nociceptin/orphanin FQ in mice. Naunyn Schmiedebergs Arch Pharmacol 363: 161–165.
Rizzi A, Nazzaro C, Marzola G, Zucchini S, Trapella C, Guerrini R et al (2006). Endogenous nociceptin/orphanin FQ signalling produces opposite spinal antinociceptive and supraspinal pronociceptive effects in the mouse formalin test: pharmacological and genetic evidences. Pain 124: 100–108.
Rizzi A, Spagnolo B, Wainford RD, Fischetti C, Guerrini R, Marzola G et al (2007). In vitro and in vivo studies on UFP-112, a novel potent and long lasting agonist selective for the nociceptin/orphanin FQ receptor. Peptides 28: 1240–1251.
Rolland AS, Herrero MT, Garcia-Martinez V, Ruberg M, Hirsch EC, François C (2007). Metabolic activity of cerebellar and basal ganglia-thalamic neurons is reduced in parkinsonism. Brain 130: 265–275.
Rozas G, Guerra MJ, Labandeira-Garcia JL (1997). An automated rotarod method for quantitative drug-free evaluation of overall motor deficits in rat models of parkinsonism. Brain Res Brain Res Protoc 2: 75–84.
Sakoori K, Murphy NP (2004). Central administration of nociceptin/orphanin FQ blocks the acquisition of conditioned place preference to morphine and cocaine, but not conditioned place aversion to naloxone in mice. Psychopharmacology 172: 129–136.
Sakoori K, Murphy NP (2008). Endogenous Nociceptin (Orphanin FQ) Suppresses Basal Hedonic State and Acute Reward Responses to Methamphetamine and Ethanol, but Facilitates Chronic Responses. Neuropsychopharmacology 33: 877–891.
Sanberg PR, Bunsey MD, Giordano M, Norman AB (1988). The catalepsy test: its ups and downs. Behav Neurosci 102: 748–759.
Sanes JN, Suner S, Donoghue JP (1990). Dynamic organization of primary motor cortex output to target muscles in adult rats. I. Long-term patterns of reorganization following motor or mixed peripheral nerve lesions. Exp Brain Res 79: 479–491.
Sbrenna S, Marti M, Morari M, Calò G, Guerrini R, Beani L et al (2000). Modulation of 5-hydroxytryptamine efflux from rat cortical synaptosomes by opioids and nociceptin. Br J Pharmacol 130: 425–433.
Schallert T, De Ryck M, Whishaw IQ, Ramirez VD, Teitelbaum P (1979). Excessive bracing reactions and their control by atropine and L-DOPA in an animal analog of Parkinsonism. Exp Neurol 64: 33–43.
Siniscalchi A, Rodi D, Morari M, Marti M, Cavallini S, Marino S et al (2002). Direct and indirect inhibition by nociceptin/orphanin FQ on noradrenaline release from rodent cerebral cortex in vitro. Br J Pharmacol 136: 1178–1184.
Steiner H, Kitai ST (2000). Regulation of rat cortex function by D1 dopamine receptors in the striatum. J Neurosci 20: 5449–5460.
Sulaiman MR, Niklasson M, Tham R, Dutia MB (1999). Modulation of vestibular function by nociceptin/orphanin FQ: an in vivo and in vitro study. Brain Res 828: 74–82.
Varty GB, Hyde LA, Hodgson RA, Lu SX, McCool MF, Kazdoba TM et al (2005). Characterization of the nociceptin receptor (ORL-1) agonist, Ro64-6198, in tests of anxiety across multiple species. Psychopharmacology (Berl) 182: 132–143.
Voloshin MY, Lukhanina EP, Kolomietz BP, Prokopenko VF, Rodionov VA (1994). Electrophysiological investigation of thalamic neuronal mechanisms of motor disorders in parkinsonism: an influence of D2ergic transmission blockade on excitation and inhibition of relay neurons in motor thalamic nuclei of cat. Neuroscience 62: 771–781.
Wichmann T, DeLong MR (1993). Pathophysiology of parkinsonian motor abnormalities. Adv Neurol 60: 53–61.
Author information
Authors and Affiliations
Corresponding author
Additional information
DISCLOSURE/CONFLICT OF INTEREST
We declare that this work has been funded by grants from the Italian Ministry of the University (FIRB Internazionalizzazione). We declare that, except for income received from my primary employer, no financial support or compensation has been received from any individual or corporate entity over the past 3 years for research or professional service and there are no personal financial holdings that could be perceived as constituting a potential conflict of interest.
Matteo Marti, Riccardo Viaro, Remo Guerrini, Gianfranco Franchi, Michele Morari are fully paid by the University of Ferrara.
Supplementary Information accompanies the paper on the Neuropsychopharmacology website (http://www.nature.com/npp)
Supplementary information
Rights and permissions
About this article
Cite this article
Marti, M., Viaro, R., Guerrini, R. et al. Nociceptin/Orphanin FQ Modulates Motor Behavior and Primary Motor Cortex Output Through Receptors Located in Substantia Nigra Reticulata. Neuropsychopharmacol 34, 341–355 (2009). https://doi.org/10.1038/npp.2008.56
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2008.56
Keywords
This article is cited by
-
A systematic review of the role of the nociceptin receptor system in stress, cognition, and reward: relevance to schizophrenia
Translational Psychiatry (2018)
-
Panicolytic-like effects caused by substantia nigra pars reticulata pretreatment with low doses of endomorphin-1 and high doses of CTOP or the NOP receptors antagonist JTC-801 in male Rattus norvegicus
Psychopharmacology (2017)
-
Involvement of Neurotransmitters in the Action of the Nociceptin/Orphanin FQ Peptide-Receptor System on Passive Avoidance Learning in Rats
Neurochemical Research (2014)
-
Gender differences in Nociceptin/Orphanin FQ-induced food intake in strains derived from rats prone (WOKW) and resistant (Dark Agouti) to metabolic syndrome: a possible involvement of the cocaine- and amphetamine-regulated transcript system
Genes & Nutrition (2011)
