
Abstract
The hypothalamic arcuate–median eminence complex (Arc-ME) controls energy balance, fertility and growth through molecularly distinct cell types, many of which remain unknown. To catalog cell types in an unbiased way, we profiled gene expression in 20,921 individual cells in and around the adult mouse Arc-ME using Drop-seq. We identify 50 transcriptionally distinct Arc-ME cell populations, including a rare tanycyte population at the Arc-ME diffusion barrier, a new leptin-sensing neuron population, multiple agouti-related peptide (AgRP) and pro-opiomelanocortin (POMC) subtypes, and an orexigenic somatostatin neuron population. We extended Drop-seq to detect dynamic expression changes across relevant physiological perturbations, revealing cell type–specific responses to energy status, including distinct responses in AgRP and POMC neuron subtypes. Finally, integrating our data with human genome-wide association study data implicates two previously unknown neuron populations in the genetic control of obesity. This resource will accelerate biological discovery by providing insights into molecular and cell type diversity from which function can be inferred.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Accession codes
Primary accessions
Gene Expression Omnibus
Referenced accessions
NCBI Reference Sequence
References
Rodríguez, E.M., Blázquez, J.L. & Guerra, M. The design of barriers in the hypothalamus allows the median eminence and the arcuate nucleus to enjoy private milieus: the former opens to the portal blood and the latter to the cerebrospinal fluid. Peptides 31, 757–776 (2010).
Langlet, F. et al. Tanycytic VEGF-A boosts blood-hypothalamus barrier plasticity and access of metabolic signals to the arcuate nucleus in response to fasting. Cell Metab. 17, 607–617 (2013).
Aponte, Y., Atasoy, D. & Sternson, S.M. AGRP neurons are sufficient to orchestrate feeding behavior rapidly and without training. Nat. Neurosci. 14, 351–355 (2011).
Krashes, M.J. et al. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J. Clin. Invest. 121, 1424–1428 (2011).
Han, S.Y., McLennan, T., Czieselsky, K. & Herbison, A.E. Selective optogenetic activation of arcuate kisspeptin neurons generates pulsatile luteinizing hormone secretion. Proc. Natl. Acad. Sci. USA 112, 13109–13114 (2015).
Vong, L. et al. Leptin action on GABAergic neurons prevents obesity and reduces inhibitory tone to POMC neurons. Neuron 71, 142–154 (2011).
Kong, D. et al. GABAergic RIP-Cre neurons in the arcuate nucleus selectively regulate energy expenditure. Cell 151, 645–657 (2012).
Macosko, E.Z. et al. Highly parallel genome-wide expression profiling of individual cells using nanoliter droplets. Cell 161, 1202–1214 (2015).
Marques, S. et al. Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system. Science 352, 1326–1329 (2016).
Usoskin, D. et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 18, 145–153 (2015).
Vijayan, E., Samson, W.K. & McCann, S.M. In vivo and in vitro effects of cholecystokinin on gonadotropin, prolactin, growth hormone and thyrotropin release in the rat. Brain Res. 172, 295–302 (1979).
Morgan, P.J. & Williams, L.M. The pars tuberalis of the pituitary: a gateway for neuroendocrine output. Rev. Reprod. 1, 153–161 (1996).
Goodman, T. & Hajihosseini, M.K. Hypothalamic tanycytes-masters and servants of metabolic, neuroendocrine, and neurogenic functions. Front. Neurosci. 9, 387 (2015).
Lein, E.S. et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 445, 168–176 (2007).
Tesfaigzi, J. & Carlson, D.M. Expression, regulation, and function of the SPR family of proteins. A review. Cell Biochem. Biophys. 30, 243–265 (1999).
Everitt, B.J. et al. The hypothalamic arcuate nucleus-median eminence complex: immunohistochemistry of transmitters, peptides and DARPP-32 with special reference to coexistence in dopamine neurons. Brain Res. 396, 97–155 (1986).
Choudhury, A.I. et al. The role of insulin receptor substrate 2 in hypothalamic and beta cell function. J. Clin. Invest. 115, 940–950 (2005).
Garfield, A.S. et al. Dynamic GABAergic afferent modulation of AgRP neurons. Nat. Neurosci. 19, 1628–1635 (2016).
Lyons, D.J., Hellysaz, A. & Broberger, C. Prolactin regulates tuberoinfundibular dopamine neuron discharge pattern: novel feedback control mechanisms in the lactotrophic axis. J. Neurosci. 32, 8074–8083 (2012).
Demaria, J.E. et al. Dopamine transporters participate in the physiological regulation of prolactin. Endocrinology 141, 366–374 (2000).
Henry, F.E., Sugino, K., Tozer, A., Branco, T. & Sternson, S.M. Cell type-specific transcriptomics of hypothalamic energy-sensing neuron responses to weight-loss. Elife 4, 09800 (2015).
Sohn, J.W. et al. Serotonin 2C receptor activates a distinct population of arcuate pro-opiomelanocortin neurons via TRPC channels. Neuron 71, 488–497 (2011).
Kahn, D., Abrams, G.M., Zimmerman, E.A., Carraway, R. & Leeman, S.E. Neurotensin neurons in the rat hypothalamus: an immunocytochemical study. Endocrinology 107, 47–54 (1980).
Nishiyama, T. et al. Hypothalamic thyrotropin-releasing hormone (TRH)-containing neurons involved in the hypothalamic-hypophysial-thyroid axis. Light microscopic immunohistochemistry. Brain Res. 345, 205–218 (1985).
Liu, J. et al. Tbx19, a tissue-selective regulator of POMC gene expression. Proc. Natl. Acad. Sci. USA 98, 8674–8679 (2001).
Takayasu, S. et al. A neuropeptide ligand of the G protein-coupled receptor GPR103 regulates feeding, behavioral arousal, and blood pressure in mice. Proc. Natl. Acad. Sci. USA 103, 7438–7443 (2006).
Chitravanshi, V.C., Kawabe, K. & Sapru, H.N. Mechanisms of cardiovascular actions of urocortins in the hypothalamic arcuate nucleus of the rat. Am. J. Physiol. Heart Circ. Physiol. 305, H182–H191 (2013).
Reyes, T.M. et al. Urocortin II: a member of the corticotropin-releasing factor (CRF) neuropeptide family that is selectively bound by type 2 CRF receptors. Proc. Natl. Acad. Sci. USA 98, 2843–2848 (2001).
Vaisse, C. et al. Leptin activation of Stat3 in the hypothalamus of wild-type and ob/ob mice but not db/db mice. Nat. Genet. 14, 95–97 (1996).
Kong, W. et al. A role for arcuate cocaine and amphetamine-regulated transcript in hyperphagia, thermogenesis, and cold adaptation. FASEB J. 17, 1688–1690 (2003).
Asnicar, M.A. et al. Vasoactive intestinal polypeptide/pituitary adenylate cyclase-activating peptide receptor 2 deficiency in mice results in growth retardation and increased basal metabolic rate. Endocrinology 143, 3994–4006 (2002).
Novak, C.M., Zhang, M. & Levine, J.A. Neuromedin U in the paraventricular and arcuate hypothalamic nuclei increases non-exercise activity thermogenesis. J. Neuroendocrinol. 18, 594–601 (2006).
Tong, Q., Ye, C.P., Jones, J.E., Elmquist, J.K. & Lowell, B.B. Synaptic release of GABA by AgRP neurons is required for normal regulation of energy balance. Nat. Neurosci. 11, 998–1000 (2008).
Kuperman, Y. et al. CRFR1 in AgRP neurons modulates sympathetic nervous system activity to adapt to cold stress and fasting. Cell Metab. 23, 1185–1199 (2016).
Moga, M.M. & Saper, C.B. Neuropeptide-immunoreactive neurons projecting to the paraventricular hypothalamic nucleus in the rat. J. Comp. Neurol. 346, 137–150 (1994).
Betley, J.N., Cao, Z.F., Ritola, K.D. & Sternson, S.M. Parallel, redundant circuit organization for homeostatic control of feeding behavior. Cell 155, 1337–1350 (2013).
Atasoy, D., Betley, J.N., Su, H.H. & Sternson, S.M. Deconstruction of a neural circuit for hunger. Nature 488, 172–177 (2012).
Garfield, A.S. et al. A neural basis for melanocortin-4 receptor-regulated appetite. Nat. Neurosci. 18, 863–871 (2015).
Petreanu, L., Huber, D., Sobczyk, A. & Svoboda, K. Channelrhodopsin-2-assisted circuit mapping of long-range callosal projections. Nat. Neurosci. 10, 663–668 (2007).
Armbruster, B.N., Li, X., Pausch, M.H., Herlitze, S. & Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 104, 5163–5168 (2007).
Wu, Q., Boyle, M.P. & Palmiter, R.D. Loss of GABAergic signaling by AgRP neurons to the parabrachial nucleus leads to starvation. Cell 137, 1225–1234 (2009).
Hahn, T.M., Breininger, J.F., Baskin, D.G. & Schwartz, M.W. Coexpression of Agrp and NPY in fasting-activated hypothalamic neurons. Nat. Neurosci. 1, 271–272 (1998).
Hsueh, Y.C., Cheng, S.M. & Pan, J.T. Fasting stimulates tuberoinfundibular dopaminergic neuronal activity and inhibits prolactin secretion in oestrogen-primed ovariectomized rats: involvement of orexin A and neuropeptide Y. J. Neuroendocrinol. 14, 745–752 (2002).
Pers, T.H. et al. Biological interpretation of genome-wide association studies using predicted gene functions. Nat. Commun. 6, 5890 (2015).
Locke, A.E. et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 518, 197–206 (2015).
Shungin, D. et al. New genetic loci link adipose and insulin biology to body fat distribution. Nature 518, 187–196 (2015).
Vimaleswaran, K.S. et al. Candidate genes for obesity-susceptibility show enriched association within a large genome-wide association study for BMI. Hum. Mol. Genet. 21, 4537–4542 (2012).
Fenselau, H. et al. A rapidly acting glutamatergic ARC→PVH satiety circuit postsynaptically regulated by α-MSH. Nat. Neurosci. 20, 42–51 (2017).
Boules, M., Li, Z., Smith, K., Fredrickson, P. & Richelson, E. Diverse roles of neurotensin agonists in the central nervous system. Front. Endocrinol. (Lausanne) 4, 36 (2013).
Wood, A.R. et al. Defining the role of common variation in the genomic and biological architecture of adult human height. Nat. Genet. 46, 1173–1186 (2014).
Balthasar, N. et al. Leptin receptor signaling in POMC neurons is required for normal body weight homeostasis. Neuron 42, 983–991 (2004).
Taniguchi, H. et al. A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron 71, 995–1013 (2011).
Krashes, M.J. et al. An excitatory paraventricular nucleus to AgRP neuron circuit that drives hunger. Nature 507, 238–242 (2014).
van den Pol, A.N. et al. Neuromedin B and gastrin-releasing peptide excite arcuate nucleus neuropeptide Y neurons in a novel transgenic mouse expressing strong Renilla green fluorescent protein in NPY neurons. J. Neurosci. 29, 4622–4639 (2009).
Postic, C. et al. Dual roles for glucokinase in glucose homeostasis as determined by liver and pancreatic beta cell-specific gene knock-outs using Cre recombinase. J. Biol. Chem. 274, 305–315 (1999).
Saxena, A. et al. Trehalose-enhanced isolation of neuronal sub-types from adult mouse brain. Biotechniques 52, 381–385 (2012).
Robinson, M.D., McCarthy, D.J. & Smyth, G.K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Satija, R., Farrell, J.A., Gennert, D., Schier, A.F. & Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 33, 495–502 (2015).
van der Maaten, L. & Hinton, G. Visualizing data using t-SNE. J. Mach. Learn. Res. 9, 2579–2605 (2008).
Esther, M., Kriegel, H.-P., Sander, J. & Xu, X. A density-based algorithm for discovering clusters in large spatial databases with noise. in 2nd International Conference on Knowledge Discovery and Data Mining (KDD-96) (Institute for Computer Science, University of Munich, 1996).
Kharchenko, P.V., Silberstein, L. & Scadden, D.T. Bayesian approach to single-cell differential expression analysis. Nat. Methods 11, 740–742 (2014).
Fresno, C. & Fernández, E.A. RDAVIDWebService: a versatile R interface to DAVID. Bioinformatics 29, 2810–2811 (2013).
Hempel, C.M., Sugino, K. & Nelson, S.B. A manual method for the purification of fluorescently labeled neurons from the mammalian brain. Nat. Protoc. 2, 2924–2929 (2007).
Picelli, S. et al. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protoc. 9, 171–181 (2014).
Mullier, A., Bouret, S.G., Prevot, V. & Dehouck, B. Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain. J. Comp. Neurol. 518, 943–962 (2010).
Bonilla, I.E., Tanabe, K. & Strittmatter, S.M. Small proline-rich repeat protein 1A is expressed by axotomized neurons and promotes axonal outgrowth. J. Neurosci. 22, 1303–1315 (2002).
Morris, A.P. et al. Large-scale association analysis provides insights into the genetic architecture and pathophysiology of type 2 diabetes. Nat. Genet. 44, 981–990 (2012).
Day, F.R. et al. Large-scale genomic analyses link reproductive aging to hypothalamic signaling, breast cancer susceptibility and BRCA1-mediated DNA repair. Nat. Genet. 47, 1294–1303 (2015).
Perry, J.R. et al. Parent-of-origin-specific allelic associations among 106 genomic loci for age at menarche. Nature 514, 92–97 (2014).
Boraska, V. et al. A genome-wide association study of anorexia nervosa. Mol. Psychiatry 19, 1085–1094 (2014).
Chang, C.C. et al. Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience 4, 7 (2015).
The 1000 Genomes Project Consortium. A global reference for human genetic variation. Nature 526, 68–74 (2015).
Yates, A. et al. Ensembl 2016. Nucleic Acids Res. 44 D1: D710–D716 (2016).
Acknowledgements
We gratefully acknowledge Z. Yang, J. Madara and C. Wu for technical assistance, A. Garfield for editorial advice and P. Kharchenko for advice on SCDE. Quantitative PCR and confocal imaging were done at BIDMC's Molecular Medicine Core and Confocal Imaging Core, respectively. Funding was provided by US National Institutes of Health grants to B.B.L. (R01 DK096010, R01 DK089044, R01 DK071051, R01 DK075632, R37 DK053477, BNORC Transgenic Core P30 DK046200, BADERC Transgenic Core P30 DK057521), E.D.R. (R01 DK102170, R01 DK085171, R01 DK102173), E.D.R. and L.T.T. (BNORC Functional Genomics Core P30 DK046200), L.T.T. (BADERC Pilot and Feasibility grant NIH 2P30DK057521-16) and J.M.R. (F32 DK103387); a Department of Defense grant to L.T.T. (Discovery Award W81XWH-15-1-0251); an American Heart Association Postdoctoral Fellowship to J.N.C. (14POST20100011); the Lundbeck Foundation and the Benzon Foundation (T.H.P.); the Stanley Center for Psychiatric Research (S.A.M.); and the Stanley-MGH Fellowship in Psychiatric Neuroscience (E.Z.M.).
Author information
Authors and Affiliations
Contributions
J.N.C., L.T.T., E.Z.M., S.A.M., E.D.R. and B.B.L. conceived the study. J.N.C., L.T.T., E.Z.M., E.D.R. & B.B.L. designed the study. J.N.C., A.M.J.V. and L.T.T. prepared samples for Drop-seq. L.T.T., D.T., J.N.C., E.Z.M. and M.G. did Drop-seq. D.T., M.G. and L.T.T. made Drop-seq libraries. J.N.C. did single-cell RNA-seq. L.T.T., J.N.C., A.L. and E.Z.M. analyzed transcriptomic data. J.M.R. did in situ hybridization. J.N.C. and H.F. did histology and imaging, with advice from B.B.L. H.F. did electrophysiology. H.F. and A.M.J.V. did stereotaxic injections and feeding studies. T.H.P. performed DEPICT analyses. J.N.C., L.T.T. and A.L. prepared figures. J.N.C., L.T.T., S.A.M., E.D.R. and B.B.L. wrote the manuscript with input from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Cluster composition, pars tuberalis expression of Cck, oligodendrocyte marker expression and subclustering analysis
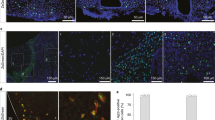
(A) Left, all-cell tSNE plot with cells colored according to treatment group/batch. Each cluster contains cells from all batches/groups. Right, a color-keyed description of each treatment group and sample batch, including: number of mice, their sex and mean age ± S.D; number of cells analyzed from each sample; and the estimated number of cells from each sample that were input to Drop-seq. (B) Co-expression of Tshb and Cck by single cells in the pars tuberalis cell clusters. (C) Micrograph of TSHβ (magenta; top-left) and Cck-IRES-Cre::loxSTOPlox-GFP (green; top-right) immunofluorescence in the pars tuberalis. Overlay (with DAPI-stained DNA; bottom) confirms that most TSHβ immunoreactive PT cells are also CCK immunoreactive. Note that micrograph panels were separately pseudocolored and contrast adjusted. Scale bar, 50μm (D) Stages of oligodendrocyte development and a heatmap showing relative expression of genes marking those developmental stages per Marques et al. (2016). (E) All-cell subcluster tSNE plot with cells colored according to results of subclustering of non-neuronal populations, in which each spatially distinct cluster on the original tSNE plot was independently re-clustered to further describe heterogeneity. (F, G) Volcano plots comparing averaged gene expression in “mural cells” subclusters s12 (pericytes) and s13 (smooth muscle cells) and in “macrophages” subclusters s14 (microglia) and s15 (perivascular macrophages), with selected subcluster-specific marker genes labeled, including well-described markers (orange) that guided our subcluster annotation.
Supplementary Figure 2 Ependymal cell clusters and marker expression
(A) Ependymal cell clusters derived from original tSNE plot in Supplemental Figure 1E (thumbnail). Clusters are annotated based on anatomical localization of marker genes in Figure 2B. (B) Violin plots showing ependymal cell cluster expression of previously characterized ependymal cell subtype markers. (C) Heatmap showing single-cell expression of genes differentially expressed by the ependymal cell clusters. Genes shown are each cluster’s top10 most enriched genes by fold-difference. (D) Violin plots showing ependymal cell cluster expression of novel candidate ependymal cell subtype markers.
Supplementary Figure 3 Neuron subclustering and expression of neuropeptide- and neurotransmitter-related genes
(A) Overview of neuron-only analysis, including iterative subclustering process used to resolve spatially indistinct clusters. (B) Diagram showing selection of cells for each round of subclustering. (C) Dotplot showing cluster-average expression of neuropeptide/neurotransmitter-related genes; color of the dot indicates the relative level of expression and the size of the dot indicates the percent of neurons in that cluster expressing the gene. Cluster names in gray likely originated from outside the Arc-ME (see Supplemental Figure 5A–D).
Supplementary Figure 4 Differential expression analysis of POMC neuron clusters
(A) Heatmap of single-cell expression of genes differentiating three POMC neuron clusters (genes selected showed >2 fold change differential expression between POMC neuron clusters). (B) Volcano plots comparing averaged gene expression of three POMC neuron clusters.
Supplementary Figure 5 Dissection scheme, regional marker expression, and RIP-Cre neuron gene expression
(A) Nissl stained mouse brain sections (from Allen Mouse Brain Atlas) marked to show dissection plan. (B) Neuron-only tSNE plot re-colored to show known vs. unknown neuron clusters. A neuron cluster was considered “known” if it generally expressed at least one previously characterized marker of Arc-ME neuron types (Agrp, Pomc, Tac2, Sst, Ghrh, or Th). (C) Expression of genes enriched in regions neighboring the Arc-ME, shown by in situ hybridization of mouse brain sagittal sections (Allen Mouse Brain Atlas; top) and by re-coloring of neuron-only tSNE plot (bottom). (D) Expression of genes enriched in Arc-ME relative to neighboring regions, shown by in situ hybridization of mouse brain sagittal sections (Allen Mouse Brain Atlas; top) and by re-coloring of neuron-only tSNE plot (bottom). (E) Histogram of gene expression in individual RIP-Cre Arc-ME neurons profiled by single-cell RNA-seq.
Supplementary Figure 6 Differential expression analysis of AgRP neuron clusters and SST neuron clusters
(A) Heatmap of single-cell expression (left) and average expression by cluster (right) of genes differentiating n13.Agrp/Gm8773 and n23.Sst/Unc13c neuron clusters. Selected genes show >2 fold-difference in expression between n13.Agrp/Gm8773 or n23.Sst/Unc13c clusters and all other arcuate neuron clusters. The n12.Agrp/Sst population expresses gene markers of both n13.Agrp/Gm8773 and n23.Sst/Unc13c clusters. (B) Left, heatmap of cluster-average expression of cluster marker genes that are differentially expressed in n13.Agrp/Gm8773 and n12.Agrp/Sst clusters. Hierarchical clustering dendrogram (right) of Arc-ME neuron subtypes using these genes reveals a gene expression program shared by n12.Agrp/Sst and other Sst-expressing arcuate populations. Dendrogram branches of arcuate clusters expressing significant Sst are colored orange. (C) Relative densities of axon innervation of various brain regions by AgRP neurons and ARCSST neurons. (D) Heatmap of single-cell expression of genes differentiating the Sst+ neuron clusters. Selected genes show >2 fold-difference in expression between Sst+ populations. (E) Heatmap of cluster-average expression, highlighting individual genes related to neuropeptide/neurotransmitter signaling, extracellular receptors, and transcriptional regulation that are differentially expressed in Sst+ clusters.
Supplementary Figure 7 Transcriptional effects of fasting and high-fat diet
(A) Number of genes significantly affected by fasting and high-fat diet in each All-Cell cluster. (B) Volcano plots of genes affected by fasting vs. HFD in Agrp/Gm8773 neurons. (C) Comparison of fasting-induced gene expression in AgRP neuron subtypes with previously published data from pooled AgRP neurons (left) and pooled POMC neurons (right) (Henry et al., 2015). AgRP neuron subclusters correlated well with pooled AgRP neuron data (C) but not with pooled POMC neuron data (D). (C) n13.Agrp/Gm8773: slope=1.4444, r2=0.33556, n=741 genes, p=3.86e−59; n12.Agrp/Sst: slope=1.0155, r2=0.41957, n=245 genes, p=1.82e−33. (D) n13.Agrp/Gm8773: slope=-0.5117, n=38 genes, p=0.283; n12.Agrp/Sst: slope=-0.3535, n=13 genes, r2=0.03115, p=0.534. (E) Comparison of fasting-induced gene expression in POMC neuron subtypes with previously published data from pooled POMC neurons (E) and pooled AgRP neurons (F) from Henry et al. (2015). POMC neuron subclusters correlated well with pooled POMC neuron data (E) but not with pooled AgRP neuron data (F). (E) n15.Pomc/Anxa2: slope=0.82, r2=0.67463, n=12 genes, p=1.82e−33; n14.Pomc/Ttr: slope=1.522, r2=0.64653, n=10 genes, p=8.13e−5. (F) n15.Pomc/Anxa2: slope=-1.8, r2=0.93174, n=4 genes, p=2.87e−7; n14.Pomc/Ttr: slope=-0.59, r2=0.56174, n=4 genes, p=0.11. (G) Heatmap of fasting responses of all Arc-ME neurons for genes differentially expressed in at least one neuron subcluster (FDR<0.05; fold-change, FC>0.75). Top significant GO terms using DAVID Gene ontology analysis are shown. (H) Heatmap of fasting vs. HFD responses for genes differentially expressed in at least one AgRP neuron subtype or POMC neuron subtype (FDR<0.05; FC>0.75). Top significant GO terms using DAVID Gene ontology analysis are shown.
Supplementary Figure 8 DEPICT analysis for height, menarche, and menopause
(A) DEPICT –log10 p values for enrichment each Arc-ME cell type for height, menarche, and menopause. (B) DEPICT –log10 p values for Arc-ME neuron types. Dotted line at p=0.01.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–8 (PDF 2910 kb)
Supplementary Table 1: The number and percentage representation of cells within each all-cell and neuron cluster parsed by experiment, age, sex, and dietary intervention.
Number and percentage are presented in reference to total cells/neurons evaluated (top) or total Arc-ME cells/neurons (bottom) to account for dissection artifacts. Likely sex-of-origin of Fasted and Re-fed cells was determined using MLSeq package (see Online Methods). Red lettering indicates non-arcuate cell clusters. (XLSX 93 kb)
Supplementary Table 2: For each all-cell cluster, a table of fold-change values for genes differentially expressed with false-discovery rate (FDR) <25% in at least one comparison, sorted based on specificity (i.e., number of positive fold-change values) and average expression.
Fold-change values with FDR>25% are indicated by zeroes. (XLSX 38327 kb)
Supplementary Table 3: For each all-cell subcluster, a table of fold-change values for genes differentially expressed with false-discovery rate (FDR) <25% in at least one comparison, sorted based on specificity (i.e., number of positive fold-change values) and average expression.
Fold-change values with FDR>25% are indicated by zeroes. (XLSX 98822 kb)
Supplementary Table 4: For each neuronal cluster, a table of fold-change values for genes differentially expressed with false-discovery rate (FDR) <25% in at least one comparison, sorted based on specificity (i.e., number of positive fold-change values) and average expression.
Fold-change values with FDR>25% are indicated by zeroes. (XLSX 61278 kb)
Supplementary Table 5
For each all-cell cluster from fed vs. fasted mice, tables of fold-change values, false-discovery rates (FDR), and fold-change values with at least one significant comparison (FDR<25%). (XLSX 4484 kb)
Supplementary Table 6
For each all-cell cluster from low-fat diet vs. high-fat diet fed mice, tables of fold-change values, false-discovery rates (FDR), and fold-change values with at least one significant comparison (FDR<25%). (XLSX 3630 kb)
Supplementary Table 7
For each neuronal cluster from fed vs. fasted mice, tables of fold-change values, false-discovery rates (FDR), and fold-change values with at least one significant comparison (FDR<25%). (XLSX 6084 kb)
Supplementary Table 8
For each neuronal cluster from low-fat diet vs. high-fat diet fed mice, tables of fold-change values, false-discovery rates (FDR), and fold-change values with at least one significant comparison (FDR<25%). (XLSX 4280 kb)
Supplementary Table 9
(DEPICT_GWAS Sources Tab) Overview of GWAS p value cutoffs used to define loci, the number of resulting DEPICT loci (defined by linkage disequilibrium r2>0.5), the number of genes in DEPICT loci, PubMed identifiers of the publications from which the GWAS summary statistics were sourced, and links to the GWAS summary statistics file. For the GWAS only waist hip ratio summary statistics, please contact the authors. (TRH_Lef1_CandidateObesityGenes Tab) Evidence in support of inclusion as candidate gene for obesity with increased expression in the n25.Trh/Lef1 neuron cluster, adapted from Vimaleswaran et al. DEPICT_Statistics tab. (Slc17a6_Trhr_CandidateObesityGen Tab) Evidence in support of inclusion as candidate gene for obesity with increased expression in the n32.Slc17a6/Trhr neuron cluster, adapted from Vimaleswaran et al. (DEPICT_Statistics Tab) Summary DEPICT p-values and FDR for included GWAS phenotypes represented in Figure 8 and Supplemental Figure 8. (XLSX 37 kb)
Rights and permissions
About this article

Cite this article
Campbell, J., Macosko, E., Fenselau, H. et al. A molecular census of arcuate hypothalamus and median eminence cell types. Nat Neurosci 20, 484–496 (2017). https://doi.org/10.1038/nn.4495
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nn.4495
This article is cited by
-
Maternal dietary fat during lactation shapes single nucleus transcriptomic profile of postnatal offspring hypothalamus in a sexually dimorphic manner in mice
Nature Communications (2024)
-
A brainstem–hypothalamus neuronal circuit reduces feeding upon heat exposure
Nature (2024)
-
An RNA-seq atlas of mouse brain areas during fasting and diet-induced obesity
Scientific Data (2024)
-
Single-Cell Transcriptome Reveals Cell Type–Specific Molecular Pathology in a 2VO Cerebral Ischemic Mouse Model
Molecular Neurobiology (2024)
-
Comprehensive mapping of Epithelial Na+ channel α expression in the mouse brain
Brain Structure and Function (2024)
