
Abstract
The prefrontal cortex helps adjust an organism's behavior to its environment. In particular, numerous studies have implicated the prefrontal cortex in the control of social behavior, but the neural circuits that mediate these effects remain unknown. Here we investigated behavioral adaptation to social defeat in mice and uncovered a critical contribution of neural projections from the medial prefrontal cortex to the dorsal periaqueductal gray, a brainstem area vital for defensive responses. Social defeat caused a weakening of functional connectivity between these two areas, and selective inhibition of these projections mimicked the behavioral effects of social defeat. These findings define a specific neural projection by which the prefrontal cortex can control and adapt social behavior.
Similar content being viewed by others

Main
The medial prefrontal cortex (mPFC) plays an important role in generating appropriate social responses by supporting behavioral flexibility, response inhibition, attention and emotion. It has been proposed that the mPFC evaluates and interprets information within the context of past experiences and is thus critical for selecting suitable behavioral responses within a social environment1. For example, lesions and pharmacological manipulations of the rodent mPFC modify aggression between males2, are required for sex differences in social anxiety3, modulate social position within a hierarchy4 and support learned behavioral responses to social defeat5,6, highlighting the importance of this structure in interpreting and modifying social behaviors in the context of past social experiences.
The mPFC projects to several brain areas that are known to influence sociability, including amygdala, nucleus accumbens, hippocampus and brainstem7. However, although several of these projections have been shown to be critical for mPFC control of nonsocial behaviors8,9 and mPFC projections to the raphe nucleus are able to interfere with the consolidation of adaptation to social defeat6, until now the mPFC outputs that directly modulate social behavior have not been identified. Here we investigated whether projections from mPFC to the dorsal periaqueductal gray (PAG), a brainstem motor control area essential for defensive responses to social threats10,11,12, might play a role in the behavioral adaptation to social defeat. This adaptive response, occurring as a result of repeated exposure to threatening members of the same species, is characterized by a shift toward a more socially avoidant behavioral strategy13, which is presumably aimed at diminishing future harm and facilitating alternative routes to essential resources14. The adaptation to social defeat in animals may have clinical relevance, because mood disorders, including major depression and social anxiety disorder, are thought to involve an extreme form of an adaptive coping strategy elicited by social adversity15,16.
We found that repeated social defeat resulted in increased social avoidance and impaired working memory, both phenotypes that were ameliorated by the antidepressant ketamine. Selective pharmacogenetic inhibition of mPFC projections to PAG mimicked the effect of social defeat, increasing social avoidance and disinhibiting PAG. Social defeat caused a reduction in functional connectivity between mPFC and PAG, resembling observations made in imaging studies of patients with affective disorders17. Cell-type-specific rabies virus tracing and ex vivo channelrhodopsin (ChR2)-assisted circuit mapping demonstrated that layer 5 mPFC projection neurons directly inhibit excitatory inputs to glutamatergic neurons in PAG, and selective inhibition of these target neurons reduced social avoidance. These findings identify a specific projection by which the prefrontal cortex controls social behavior and demonstrates how these inputs can be modulated to adapt social behavior to the environment.
Results
Glutamatergic mPFC projections to dorsal PAG
Anterograde and retrograde tracer studies have demonstrated prominent neural projections from the rat mPFC to PAG18,19. However, the precise location and cell identity of these projections have not been described. Moreover, although mPFC projection neurons are thought to be primarily glutamatergic, at least one study has demonstrated that GABAergic mPFC neurons project to the nucleus accumbens (NAc) and are capable of inducing avoidance behavior in a place-preference task20. To determine the identity of mPFC neurons that project to dorsal PAG (dPAG; we use this term to refer to the entire dorsal half of the PAG, including the dorsomedial, dorsolateral and lateral columns), we simultaneously injected differentially fluorescent cholera toxin B retrograde tracers into NAc and dPAG (Fig. 1a,b) and visualized retrogradely labeled mPFC neurons. Labeled neurons projecting to NAc were located primarily in layer 2/3 with some labeled cells seen in layer 5 (Fig. 1c). Labeled neurons projecting to dPAG, on the other hand, were exclusively located in layer 5 (Fig. 1d), consistent with layer 5 harboring cortical projection neurons targeting brainstem motor areas18,19. No overlap between NAc and dPAG projecting neurons was observed (0/791 and 0/594 neurons, respectively) arguing for a differential identity of these neurons in layer 5.

(a–d) Mice were injected with retrograde tracers (CTB647, green) in (a) dPAG and (CTB555, red) in (b) NAc. (c) Low and (d) high magnification images of retrogradely labeled CTB647 (dPAG-projecting) and CTB555 (NAc-projecting) neurons in layer 5 and layer 2/3, respectively, of mPFC. latPAG, lateral PAG; CG, anterior cingulate cortex; PL, prelimbic cortex; IL, infralimbic cortex; aca, anterior commissure, anterior; AcbSh, nucleus accumbens shell; AcbC, nucleus accumbens core. (e) Retrogradely labeled CTB647 (dPAG projecting) neurons in mPFC of a Thy1::GFP mouse. (f) Retrogradely labeled CTB647 (dPAG-projecting) cells demonstrate that they are not colocalized with GABAergic neurons in mPFC of Gad2::tdTomato mouse. Scale bar: 500 μm in a–c, 100 μm in d and f, and 50 μm in e. n = 2.
To identify the specific cell-types involved, we first repeated the retrograde labeling experiment in Thy1::GFP-M transgenic mice, in which sparse GFP labeling facilitates the morphological identification of neurons. Layer 5 mPFC neurons projecting to dPAG could be overwhelmingly identified as pyramidal in morphology, consistent with a glutamatergic identity (Fig. 1e). Second, the retrograde labeling experiment was repeated in Gad2::tdTomato transgenic mice, in which GABAergic neurons are fluorescently labeled. No overlap between mPFC neurons projecting to dPAG and the GABAergic marker was detected (0/583 neurons; Fig. 1f and Supplementary Table 1). These results suggest that, unlike the mPFC–NAc pathway, the mPFC–dPAG pathway consists exclusively of layer 5 glutamatergic projection neurons.
Social defeat induces social avoidance
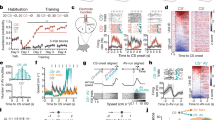
Chronic exposure of mice to an aggressor leads to social avoidance and also causes more generalized changes in anxiety and depression-like behavior21, which might confound our search for plastic changes in the brain that drive behavioral adaptation to social threat. As a result, we sought to establish a subchronic social defeat model associated with a selective adaptation of social behavior. Initially, we exposed male mice in their home cage once a day for 5 min to an aggressive conspecific confined behind a wire mesh barrier, and then allowed them to freely interact for a further 10 min. During this 10-min period, the intruder repeatedly attacked the resident. Over 7 d of social defeat, resident mice exhibited a gradual increase in upright submissive postures and freezing and a decrease in rearing during the direct encounter with the aggressor (Fig. 2a). In addition, a gradual increase in social avoidance was observed during the anticipatory period in which the aggressor remained confined to the wire mesh barrier (Supplementary Fig. 1a). Importantly, the number of attacks received by the resident did not differ across days (Supplementary Fig. 1b) demonstrating that the changes in behavior elicited in the resident reflected a gradual adaptation to repeated social defeat. Because the behavioral adaptation of the resident tended to plateau after 4 d of social defeat, we chose a 3-day defeat procedure for all further experiments to reduce potential generalization or habituation to the stress exposure.

(a) Left, upright-defensive postures (day: F6,7 = 3.8, P = 0.0042; n = 8); middle, freezing (day: F6,7 = 4.2, P = 0.0022; n = 8); and right, rearing (day: F6,7 = 3.2, P = 0.012; n = 8) performed by the resident mouse when exposed to an aggressive intruder across social defeat sessions. (b) Timeline showing behavioral testing of social approach behaviors. (c) Left, time investigating a novel aggressor (defeat: F1,22 = 16.1, P = 0.006; day: F3,22 = 2.8, P = 0.047; defeat × day: F3,66 = 2.4, P = 0.079; control, n = 12; defeat, n = 12); middle, investigation bouts (defeat: F1,22 = 20.2, P = 0.0002; day: F3,22 = 2.6, P = 0.063, defeat × day: F3,66 = 2.1, P = 0.11; control, n = 12; defeat, n = 12); and right, retreats from social investigation periods (defeat: F1,17 = 57.9, P < 0.0001; day: F3,22 = 1.9, P = 0.14; defeat × day: F3,51 = 8.7, P < 0.0001; control, n = 12; defeat, n = 10) during the social defeat procedure and one week after the last defeat session. (d) Left, time spent investigating (defeat: F1,12 = 7.6, P = 0.018, stimulus: F2,12 = 12.4, P = 0.0002, defeat × stimulus: F2,24 = 8.9, P = 0.0013; control, n = 7; defeat, n = 7); right, duration of investigation bouts (defeat: F1,12 = 7.5, P = 0.018, stimulus: F2,12 = 5.0, P = 0.016, defeat × stimulus: F2,24 = 3.9, P = 0.033; control, n = 7; defeat, n = 7) of male or female intruders or a novel object. (e) Same-arm returns (t14 = 2.9, P = 0.013; control, n = 8; defeat, n = 8), (f) spontaneous alternation (t14 = 1.9, P = 0.081; control, n = 8; defeat, n = 8), (g) latency to exit the start arm (control, n = 8; defeat, n = 8) and (h) overall distance (control, n = 8; defeat, n = 8) traveled by defeated and control mice in the Y-maze. +P < 0.1; *P < 0.05; **P < 0.01; ***P < 0.001. In a and c black circles represent mean of control mice, and red squares represent mean of defeated mice. In d–h, gray circles represent individual control mice, light red squares represent individual defeated mice and horizontal bar marks mean. All error bars represent s.e.m.
To determine whether the subchronic social defeat procedure induced a persistent change in social coping strategy, we monitored the behavior of the resident mouse during the anticipatory period immediately before each defeat session (Days 1–3), as well as during a test session (Test), during which an aggressor was placed into the resident's cage within a wire mesh barrier one week later (Fig. 2b). Resident mice spent progressively less time investigating the intruder both during the social defeat procedure and one week later. Social defeat was accompanied by a progressive and persistent decrease in the duration of investigation bouts (Fig. 2c) as well as an increase in the fraction of investigation bouts that were terminated by a rapid withdrawal movement, which we called 'retreat' (Fig. 2c). Social defeat also elicited avoidance behavior when a female mouse, but not a novel object, was placed into the wire mesh barrier on the test day, suggesting a selective adaptation of social behavior (Fig. 2d and Supplementary Fig. 1c). In the Y-maze test, a short-term memory task known to depend on mPFC function22, defeated mice showed a significant increase in same arm returns, reflecting impaired working memory, but had normal latency to exit the arms and no substantial difference in distance traveled, confirming unaltered exploratory behavior (Fig. 2e–h). No significant changes in anxiety- or stress-related behavior was seen in the elevated plus maze (P = 0.38; Supplementary Fig. 1d–f) or tail suspension test (P = 0.51; Supplementary Fig. 1g), confirming the selective impact of our defeat procedure on social behavior.
Reversal of social avoidance by antidepressant treatment
Major depression is associated with increased social withdrawal and deficits in working memory that can be reversed by antidepressant treatment23,24. To test whether the behavioral effects of social defeat demonstrated here might share pharmacological substrates with clinical depression, we tested the effect of the rapidly acting antidepressant ketamine, an NMDA receptor antagonist, in our social defeat model. On the day following social defeat animals received a single systemic injection of either ketamine (2.5 or 5 mg/kg) or vehicle, and social interaction with an aggressive intruder was investigated one week later (Supplementary Fig. 2a). Ketamine treatment was associated with a dose-dependent increase of time spent investigating the intruder (Supplementary Fig. 2b). Ketamine did not significantly increase the duration of investigation bouts (P = 0.93; Supplementary Fig. 2c) but was associated with a dose-dependent reversal of the increased retreats induced by social defeat (Supplementary Fig. 2d). No difference in locomotor activity was detected between control and ketamine-treated mice (Supplementary Fig. 2e), suggesting a selective effect of the drug on social behavior. Ketamine treatment also ameliorated defeat-induced deficits in working memory but had no significant effect on latency to exit (P = 0.28) the arms or distance traveled (P = 0.17; Supplementary Fig. 2f–i). These findings demonstrate that the persistent changes in social and cognitive behavior induced by subchronic social defeat depend on neural substrates shared with antidepressant treatment.
Inhibition of mPFC–PAG projections mimics social defeat
To test whether mPFC–PAG projections might contribute to the behavioral effects of social defeat, we used a pharmacogenetic inhibition method to selectively suppress neurotransmission in mPFC–PAG projections (Fig. 3a). Mice were infected bilaterally in mPFC with an adeno-associated virus (AAV) expressing the Venus fluorescent protein and human influenza hemagglutinin (HA)-tagged hM4D (AAV-Syn::Venus-2A-HA-hM4D)12, a designer Gαi-coupled receptor activated exclusively by the otherwise inert agonist clozapine-N-oxide (CNO)25. Infected mice were implanted with a guide cannula above the dPAG and subsequently subjected to social defeat or control conditions. Several weeks after infection, HA-immunopositive afferents could be observed in PAG (Fig. 3b), confirming the presence of hM4D on direct mPFC projections to this structure. CNO or vehicle was administered locally to dPAG 5 min before behavioral testing one week after social defeat (Fig. 3c). CNO-treated control mice spent less time investigating the aggressor, displayed shorter investigation bouts and retreated more when compared to vehicle-treated control animals (Fig. 3d–f). CNO-treated control mice were indistinguishable from vehicle-treated and CNO-treated defeated mice in time spent investigating the aggressor, duration of investigation bouts and increase in retreats (Fig. 3d–f), suggesting that mPFC promotes social interaction via direct projections to PAG. Additionally, social defeat may involve a weakening of mPFC–PAG projections, an interpretation that is consistent with the observation that CNO-treated defeated mice behaved similarly to defeated mice administered vehicle (Fig. 3d–f). CNO treatment did not affect overall locomotor activity, arguing against a general role for these projections in exploratory behavior (Supplementary Fig. 3c). Lastly, we performed an mPFC-projection inhibition experiment, in which CNO was delivered to the overlying superior colliculus rather than dPAG. In this experiment, no change in social interaction behavior was detected (Supplementary Fig. 3e–g), suggesting that CNO delivery in the brain is local and affects a relatively restricted area.

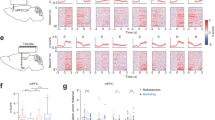
(a) Top: schematic describing bilateral infection of AAV-Syn::Venus-2A-HA-hM4D in the mPFC and implantation of a guide cannula over dPAG. Bottom: Venus-labeled infected cells (green) in the mPFC. ACg, anterior cingulate cortex; PrL, prelimbic cortex; IL, infralimbic cortex. (b) HA immunostaining showing expression of hM4D in mPFC projections in the PAG. dm, dorsomedial; dl, dorsolateral; l, lateral; vl. Ventrolateral. (c) Timeline of mPFC–dPAG projection inhibition experiments. (d) Time spent investigating the aggressor (defeat: F1,1 = 3.54, P = 0.067; CNO: F1,1 = 2.42, P = 0.13; defeat × CNO: F1,39 = 2.32, P = 0.14; t19 = 2.1, P = 0.047; control + vehicle, n = 10; control + CNO, n = 11; defeat + vehicle, n = 10; defeat + CNO, n = 12), (e) investigation bouts (defeat: F1,1 = 2.23, P = 0.14; CNO: F1,1 = 5.1, P = 0.03; defeat × CNO: F1,38 = 1.47, P = 0.23; t19 = 2.9, P = 0.0088; control + vehicle, n = 10; control + CNO, n = 10; defeat + vehicle, n = 10; defeat + CNO, n = 12) and (f) retreats (defeat: F1,1 = 2.78, P = 0.1; CNO: F1,1 = 0.54, P = 0.47; defeat × CNO: F1,38 = 2.5, P = 0.12; t19 = 2.2, P = 0.042; control + vehicle, n = 10; control + CNO, n = 10; defeat + vehicle, n = 10; defeat + CNO, n = 12) in control and defeat mice administered vehicle (saline) or CNO before testing. (g) Representative images of c-Fos immunopositive cells in the dorsomedial (dm), dorsolateral (dl) and lateral (l) PAG of mice described above. Images were selected based on mean counts of each treatment group. (h–j) Quantification of c-Fos immunopositive cells in (h) dorsomedial, (i) dorsolateral and (j) lateral PAG of mice described above (dorsomedial PAG (dmPAG), defeat × drug: F1,38 = 6.74, P = 0.013, dorsolateral PAG (dlPAG), defeat × drug: F1,38 = 6.5, P = 0.015; control + vehicle, n = 10; control + CNO, n = 10; defeat + vehicle, n = 10; defeat + CNO, n = 12). Scale bars: 200 μm in a, b and g. *P < 0.05. In d–f and h–j, gray circles represent individual control mice, light red squares represent individual defeated mice, horizontal bars mark means and error bars represent s.e.m.
Following behavioral testing, animals were killed and their brains processed for c-Fos immunohistochemistry as an indirect measure of neural activity induced in dPAG by exposure to the aggressor (Fig. 3g–j)12,26. Vehicle-treated defeated mice showed significantly more c-Fos immunopositive neurons in dPAG (dorsomedial and dorsolateral PAG) than similarly treated control mice, suggesting that enhanced activation of dPAG is a neural correlate of social defeat and consistent with a role for this structure in defensive responses to a conspecific aggressor10,12 (Fig. 3g–j and Supplementary Fig. 3h). CNO-treated control mice, on the other hand, showed an increase in c-Fos immunostaining across PAG subdivisions similar to that found in socially defeated mice, compared to vehicle-treated controls (Fig. 3h–j), demonstrating an inhibitory effect of mPFC inputs on PAG activity and corroborating a role for PAG in social avoidance. No further increase in the number of c-Fos immunopositive cells was seen in CNO-treated animals that had been exposed to social defeat when compared to similar vehicle-treated mice (Fig. 3h–j), supporting the hypothesis that the effects of mPFC–PAG inhibition are occluded in defeated animals (Fig. 3d–f).
Social defeat weakens mPFC–dPAG functional connectivity
Deficient mPFC activity, as well as reduced functional connectivity between mPFC and subcortical areas, has been reported in persons experiencing major depression or social anxiety27,28,29 suggesting that mPFC–subcortical projections might be amenable to remodeling in response to social adversity. To determine whether social defeat might weaken mPFC–dPAG projections, we examined local field potential (LFP) coherence as a measure of functional connectivity between these structures in mice undergoing social defeat (Fig. 4a). Social defeat was associated with a significant decrease in LFP coherence between mPFC and dPAG in both the theta and beta frequency bands in resident mice measured when they were physically close to the intruder during the anticipatory period preceding social defeat, compared to control animals (Fig. 4b,c). A similar trend was observed when the mice were far from the intruder (Supplementary Fig. 4b,c). Moreover, Granger causality analysis of the LFP data revealed a shift in theta causality during defeat, with a significant increase in relative dPAG–mPFC causality found in defeated animals when compared to undefeated controls (Fig. 4d and Supplementary Fig. 4d). These results suggest a greater propensity for ascending information flow in this circuit following defeat. LFP spectral power in the theta band was decreased in defeated mice in both mPFC and dPAG relative to control animals (Fig. 4e–h and Supplementary Fig. 4e–h), suggesting that changes in oscillatory activity in one or both of these structures might underlie the altered functional connectivity in the theta frequency band. These findings are consistent with changes in LFP coherence in the theta frequency band reported between mPFC and both cortical and subcortical structures during cognitive and anxiety-related behaviors in mice that have been shown to reflect altered exchange or coordination of information between structures30. Decreased coherence observed in defeated mice is not explained by any changes in oscillatory activity in either mPFC or dPAG (Fig. 4f,h), suggesting a specific decrease in functional connectivity between these regions in this frequency band.

(a) Schematic showing placement of electrodes used to measure LFP activity in mPFC and dPAG. (b,c) Relative coherence (coherence differential) in defeated (n = 7) and control (n = 7) mice (theta: U = 9, P = 0.048, beta: U = 8, P = 0.035). (d) Relative causality (causality differential) in the PAG→mPFC and mPFC→PAG direction in defeated (n = 7) and control (n = 7) mice (U = 12, P = 0.038). (e–h) Power spectra differential in (e,f) mPFC and (g,h) PAG in defeated (n = 7) and control (n = 7) mice when proximal to the aggressor (U = 6, P = 0.018). Power spectra were averaged across mice. Power in each frequency band was calculated as the sum of the power values. *P < 0.05. In c, d, f and h, gray circles represent individual control mice, light red squares represent individual defeated mice, horizontal bars mark means and error bars represent s.e.m.
Alterations in functional connectivity between brain structures as measured by LFP coherence can result from changes in synaptic connectivity between the structures, changes in the neural activity of one or the other structure, or changes in neural activity in a third structure mutually connected to the recorded structures. To test the first possibility, we recorded evoked field potentials in dPAG in response to electrical stimulation of the mPFC in mice undergoing social defeat (Fig. 5a). Periodic stimulation of mPFC during the habituation and barrier phases each testing day elicited short-latency, multimodal population responses in dPAG (Fig. 5b and Supplementary Fig. 5a). No substantial effect of social defeat could be detected across the experimental days on short-latency response amplitudes (Fig. 5b), despite significant avoidance developing in defeated animals (Fig. 5c). However, changes in synaptic strength can be encoded as changes in either postsynaptic response amplitude or presynaptic release probability. To examine possible changes in presynaptic release probability in the mPFC–dPAG pathway during social defeat, we repeated the evoked LFP experiments using a double-pulse protocol that allows for measurement of paired-pulse facilitation (PPF), a measure dependent on neurotransmitter release probability (Fig. 5d,e). Initial experiments found that maximal PPF in this pathway occurred at 50-ms pulse intervals (Fig. 5d), and this interval was used for subsequent PPF monitoring. No substantial differences in PPF were detected across testing days and groups (Fig. 5e), suggesting an absence of synaptic plasticity in the direct mPFC–dPAG pathway during social defeat.

(a) Schematic showing location of recording and stimulating (St) electrodes implanted chronically in mPFC and dPAG. (b) Field excitatory postsynaptic potential (fEPSP) amplitudes in mPFC–dPAG in defeated and control mice (control, n = 7; defeat, n = 7). (c) Time spent proximal to the aggressor in defeated and control mice with electrodes implanted in mPFC and dPAG (control, n = 7; defeat, n = 8; F1,10 = 10.51, P = 0.0088). (d) PPF of fEPSP recorded in dPAG after stimulation of mPFC, expressed as percent amplitude change (± s.e.m.) of the second fEPSP (A2) compared to the first (A1) for 5 interpulse intervals (control, n = 4; defeat, n = 4). (e) Evolution of PPF of fEPSP along the sessions recorded in the dPAG after stimulation of mPFC (control, n = 4; defeat, n = 4). (f) Schematic showing location of recording and stimulating electrodes implanted chronically in mPFC and MDT. (g) fEPSP amplitude in defeated and control mice in MDT–mPFC (control, n = 10; defeat, n = 12; F3,51 = 5.58, P = 0.0022). (h) Time spent proximal to the aggressor in defeated and control mice with electrodes implanted in MDT and dPAG (control, n = 10; defeat, n = 12; F3,57 = 13.93, P < 0.0001). *P < 0.05; *P < 0.01; ***P < 0.001. In b–e, g and h, black circles represent mean of control mice, red squares represent mean of defeated mice and error bars represent s.e.m.
Next, we tested the possibility that reduced LFP connectivity between mPFC and dPAG could be driven by changes in afferent synaptic strengths in mPFC. The mPFC receives prominent inputs from the medial dorsal nucleus of the thalamus (MDT), and reductions in this pathway have been hypothesized to occur in major depression31. To examine potential changes in this afferent pathway that could underlie weakened mPFC–dPAG functional connectivity, we measured evoked field potentials in mPFC in response to stimulation of MDT during social defeat (Fig. 5f). Periodic stimulation of MDT during the habituation and barrier phases each testing day elicited short-latency, multimodal population responses in mPFC (Fig. 5g and Supplementary Fig. 5b). A significant reduction of short-latency response amplitudes was detected across testing days in socially defeated mice when compared to control animals (Fig. 5g), which paralleled the development of avoidance (Fig. 5h). These findings demonstrate that weakening of mPFC afferent synaptic strength occurred during social defeat and suggests that changes in mPFC afferent input strength underlie the weakened functional connectivity observed between mPFC and dPAG (Fig. 4b,c).
mPFC projections target glutamatergic dPAG cells
Our anatomical tract tracing (Fig. 1) and pharmacogenetic projection inhibition (Fig. 3) data argue that glutamatergic projections from mPFC act to inhibit dPAG function. To identify the local dPAG cell types that mediate mPFC afferent control, we performed cell-type-specific monosynaptic circuit tracing using Cre-dependent pseudotyped rabies virus32. Cre-dependent AAV expressing either the pseudotyped rabies EnvA receptor TVA (AAV-Ef1a::DIO-TVA-mCherry) or the rabies virus protein G (AAV-CAG::DIO-RabiesG) were simultaneously delivered to dPAG of mice carrying either Vglut2::Cre or Gad2::Cre transgenes, followed by infection with a pseudotyped G-deleted rabies virus (ΔG-EnvA rabies-GFP; Fig. 6a). Following rabies infection, mice were killed and the brains were processed to systematically identify and visualize retrograde infected neurons (GFP+, mCherry− cells) across the entire brain rostral to the infection site. A total of 3,231 cells were identified following infection of Vglut2::Cre mice (Fig. 6b–e and Supplementary Table 2). The number of input cells present in each mouse was weighted to the density of starter cells in dPAG at the center of the infection site and then averaged (Fig. 6e). From the weighted averages, we observed that 90% of input cells were found in hypothalamus and thalamus, consistent with the major inputs of PAG deriving from diencephalic structures33. Only 6% of retrogradely infected neurons resided in cortex, of which 20/182 were found in mPFC. Overwhelmingly, labeled mPFC neurons had a pyramidal morphology (Fig. 6c), consistent with a layer 5 projection neuron identity (Fig. 1)34. Similarly, 85% of cells identified following infection of Gad2::Cre mice resided in hypothalamus or thalamus (Supplementary Fig. 6a and Supplementary Table 3), but we were unable to identify any retrogradely labeled cells in cortex of infected Gad2::Cre mice. The relatively low frequency of long-distance retrograde labeling in this line (total = 14 cells) suggested that long-distance afferents onto this class of cells were rare. These findings demonstrate that glutamatergic Vglut2+ neurons in dPAG were the major target of mPFC afferents and suggest that this cell class mediated the inhibitory input of mPFC on dPAG-mediated defensive responses.

(a) Schematic describing infection in the dPAG of Cre-dependent AAV expressing TVA-mCherry and rabies protein G and subsequent infection of EnvA pseudo-typed G-deleted rabies-GFP virus in Vglut2::Cre and Vgat::Cre mice. (b) Cre-dependent targeting of TVA-mCherry (red) and rabies-GFP (green) to Vglut2+ neurons in dPAG. (c) Low (left) and high (right) magnification images of a retrogradely labeled rabies-GFP infected layer 5 pyramidal cell in mPFC. (d) Summary of rabies-infected neurons (GFP+, mCherry−) in the forebrain of Vglut2::Cre animals (percentage of the mean number of retrograde neurons weighted to the number of starter cells present in each animal, n = 8). Thal, thalamus; hypothal, hypothalamus. (e) Number and weighted average of rabies-infected neurons in mPFC and hypothalamic nuclei (VMH, LH, AH and PMD) of Vglut2::Cre animals (n = 8). (f) Example images showing ChR2+ axonal projections (green) from mPFC in PAG, cell bodies of Vglut2+ neurons (red) and two neurons filled with biocytin and processed after whole-cell recording (cyan). Blue arrow points to a neuron with monosynaptic input from the mPFC and white arrow indicates a neuron without mPFC input. (g) Light-evoked monosynaptic excitatory postsynaptic currents (EPSCs) in a Vglut2+ neuron. Light red traces, individual trials; dark red, average. (h) Mean probability of detecting mPFC inputs in Vglut2+ and Vgat+ neurons. (i) Average EPSC onset latency across all cells (left, 3.6 ± 0.14 ms) and response peak amplitude (right, 23.1 ± 4.5 pA). (j) Example traces of spontaneous EPSC (sEPSC) recordings in a Vglut2+ neuron that did not receive direct mPFC input, before and after 20 trials of ChR2+ stimulation (20 pulses at 10 Hz) of mPFC terminals, showing a decrease in sEPSC frequency. (k) Mean change in sEPSC frequency with mPFC ChR2 stimulation for all Vglut2+ neurons (left; 58.5 ± 6% of control, P < 0.0001, n = 25) and Vgat+ neurons (middle; 91.4 ± 8% of control, P = 0.34, n = 12; t24 = 6.58, P < 0.0001). Right, light stimulation without ChR2 infection does not change sEPSC frequency. Lines show individual data points; in the left panel, blue lines are cells with direct mPFC input. Scale bars: 200 μm in b and c (left), 100 μm in c (right) and 50 μm in f. ***P < 0.001. Error bars represent s.e.m.
To test whether neural activity in Vglut2+ dPAG cells was selectively modulated by mPFC inputs as predicted by the rabies data, we performed ex vivo ChR2-mediated circuit mapping35. Following delivery of AAV-CAG::ChR2-YFP to mPFC, acute slices were taken from dPAG, and patch-clamp recording was performed to examine light-evoked synaptic responses. Experiments were performed in either Vglut2::tdTomato or Vgat::tdTomato reporter mice to allow selective recording from identified glutamatergic and GABAergic neurons (Fig. 6f and Supplementary Fig. 6b)36. Short-latency excitatory postsynaptic currents (Fig. 6g–i) were identified in a small fraction (13%) of recorded Vglut2+ cells but in none of the Vgat+ cells (Fig. 6h). However, whether they received short latency inputs or not, the majority of Vglut2+ cells showed a significant reduction in the frequency of spontaneous excitatory inputs following ChR2 activation, which was absent in control slices from noninfected animals (Fig. 6j,k). Vgat+ cells, on the other hand, did not show a significant change in spontaneous excitatory inputs following ChR2 activation (Fig. 6k), arguing for a selective inhibition of glutamatergic target cell afferents. Given the long latency of the inhibitory effect and the fact that the experiments were conducted under conditions in which light delivery failed to elicit action potentials, these findings demonstrate that glutamatergic mPFC projections directly suppressed excitatory inputs onto Vglut2+ dPAG neurons via presynaptic neuromodulatory mechanisms.
Finally, we examined the functional contribution of Vglut2+ and Gad2+ neurons in dPAG to social avoidance behavior during social defeat. Selective pharmacogenetic inhibition of neurons in dPAG was achieved by local infection of Vglut2::Cre or Gad2::Cre mice with AAV-Syn::DIO-hM4D-mCherry and subsequent systemic delivery of CNO 45 min before behavioral testing on Day 10 (Fig. 7a and Supplementary Fig. 7). For Vglut2+ neurons, a significant increase in time spent investigating the aggressor was seen in Cre+ mice when compared to Cre− littermates, whether they experienced social defeat or not (Fig. 7b). Inhibition of Vglut2+ PAG neurons had no significant effect on the duration of investigation bouts or the number of retreats (Fig. 7c,d). No substantial difference in avoidance behavior between Cre+ and Cre− mice was seen during the 3 d of social defeat before CNO administration (Supplementary Fig. 7a), ruling out a confounding effect of genotype in these results. These data demonstrate that Vglut2+ neurons in dPAG were responsible for promoting avoidance during social interaction with an aggressor, a finding that is in line with the observations that optogenetic activation of these cells produces defensive behavior and analgesia37 and that nonspecific pharmacogenetic inhibition of this structure blocks defensive responses to social threat12. On the other hand, selective pharmacogenetic inhibition of Gad2+ neurons elicited no significant change in either the time spent investigating the aggressor (P = 0.36) or the duration of investigation bouts (P = 0.27), although there was a decreased number of retreats in CNO-treated mice when compared to vehicle treated littermates (Supplementary Fig. 7d–f). These findings suggest that Gad2+ neurons in dPAG did not make a substantial contribution to social approach behavior, at least under the conditions used in our experiments, but they may promote some aspects of defensive behavior.

(a) AAV-Syn::DIO-hM4D-mCherry infection in the dPAG of Vglut2::Cre mice. (b) Time spent investigating the intruder (defeat: F1,11 = 26.77, P = 0.0003; genotype: F1,11 = 13.12, P = 0.004), (c) investigation bouts (defeat: F1,11 = 6.72, P = 0.025) and (d) retreats (defeat: F1,11 = 22.28, P = 0.0006) in defeated and control Vglut2::Cre mice that received systemic administration of CNO before testing. Cre−, n = 6; Cre+, n = 7. Scale bar, 500 μm. **P < 0.01, effect of genotype. In b–d, gray circles represent individual control mice, light red squares represent individual defeated mice, horizontal bars mark means and error bars represent s.e.m.
Discussion
Considerable data has implicated neural activity in mPFC in the direct modulation of social behavior4,5, but until now the projections mediating this effect were unknown. Our data demonstrate that the modulation of social approach and avoidance behavior by mPFC was mediated via direct projections to PAG, a structure required for the expression of innate motivated behaviors including defense, aggression, sex, maternal care, hunting and foraging12,38,39,40. Moreover, the existence of major mPFC projections to both dorsal (defense-related) and lateral (approach-related) behavioral control columns in PAG (Fig. 3c) suggests that these direct projections are likely to play important roles in the cortical modulation of behavioral adaptation under multiple environmental conditions, not just those described here. For example, firing of specific classes of neurons in mPFC has been shown to correlate with behavioral engagement and disengagement during foraging41, and mPFC is proposed to play a general role in decision-making in the face of environmental uncertainty42,43.
We used retrograde tracing, trans-synaptic rabies labeling and ex vivo electrophysiology to show that layer 5 glutamatergic neurons in mPFC made monosynaptic excitatory connections onto glutamatergic neurons in dPAG and that, unlike mPFC projections to NAc, GABAergic neurons did not contribute to these afferents (Figs. 1 and 6). Our discovery that these neurons were exclusively layer 5 excitatory pyramidal neurons is consistent with known projections from this cortical layer to brainstem motor control areas involved in triggering and modulating behavior18,19. Moreover, simultaneous retrograde labeling from dPAG and NAc showed that these mPFC projection neurons were non-overlapping (Fig. 1). NAc afferents arose primarily from mPFC neurons residing in layer 2/3 and included long-range GABAergic neurons. This distinction suggests that different neuronal firing information was provided by mPFC to dPAG and NAc, a structure implicated in reward and behavioral selection. ChR2-assisted circuit mapping showed that only a small fraction (∼10%) of Vglut2+ neurons in dPAG received direct excitatory mPFC inputs but that the vast majority received strong indirect inhibitory mPFC inputs via presynaptic neuromodulatory mechanisms (Fig. 6). These findings suggest that glutamatergic mPFC projection neurons exerted an inhibitory effect on dPAG by suppressing excitatory PAG afferents, possibly including those from medial hypothalamic regions that promote defensive behavior. Our findings raise the possibility that the small fraction of dPAG neurons receiving direct mPFC excitatory inputs may represent a specialized subclass of Vglut2+ neurons (Fig. 6). Our behavioral findings showing that selective pharmacogenetic inhibition of Vglut2+ but not Gad2+ cells in dPAG increased social approach are consistent with a selective inhibitory presynaptic effect on Vglut2+ neurons in dPAG (Fig. 7 and Supplementary Fig. 7).
We established a subchronic social defeat procedure that induced long-lasting increase in avoidance of social stimuli, including aggressive males as well as females, but not of nonsocial stimuli, such as novel objects (Fig. 2). Under these conditions, selective inhibition of mPFC–dPAG projections by pharmacogenetic hM4D-mediated projection inhibition caused a disinhibition of neural activity in dPAG and an increase in social avoidance (Fig. 3). The observation that projection inhibition was not effective in socially defeated mice (Fig. 3) suggested that the pathway was weakened by social defeat. This hypothesis was corroborated by LFP coherence data demonstrating a reduction in mPFC–dPAG functional connectivity in defeated mice and a switch in direction of causality, with dPAG driving mPFC more strongly in defeated mice (Fig. 4). Follow-up experiments using evoked field potential recording in behaving mice found that weakened functional connectivity between mPFC and dPAG was driven by a decrease in synaptic strength of afferent inputs to mPFC in the absence of any change in presynaptic or postsynaptic strength in the direct mPFC–dPAG pathway (Fig. 5).
Our data have several implications. First, they support a critical role for dPAG in social behavior. Extensive lesion, pharmacological and imaging data implicate dPAG in defensive responses to predators44,45,46. However, recent data show that dPAG is also required for flight, freezing and avoidance behavior following exposure of rodents to aggressive conspecifics12,26. Our findings extend this role to social avoidance in anticipation of threat (Fig. 2). Such a role in modulating anticipatory avoidance is consistent with human imaging data demonstrating a rapid switch of blood-oxygen-level-dependent (BOLD) signal activity from mPFC to dPAG in anticipation of predators11 or predator-like46 visual stimuli and suggests that dPAG may be involved in anxiety- as well as fear-related behaviors across species.
Second, our data demonstrate that functional connectivity between mPFC and dPAG can be moderated by social experience. Our in vivo evoked field potential experiments failed to find significant alterations in presynaptic or postsynaptic strengths in the mPFC–dPAG pathway during defeat but instead found a significant reduction in evoked responses in mPFC to thalamic stimulation (Fig. 5). These data suggest that mPFC–dPAG functional connectivity was weakened by a reduction in upstream afferent drive during defeat. Numerous studies have found that dendrites of mPFC pyramidal neurons can atrophy in response to chronic stress47. Reductions in the amplitude of excitatory inputs onto mPFC layer 5 pyramidal neurons were observed in subordinate mice, and bidirectional manipulation of these receptors was sufficient to induce changes in stable hierarchies among cage mates4. Notably, one current theory of the physiological deficits underlying major depression proposes that reductions in thalamic inputs to mPFC are associated with a switch in mPFC processing from external to internal sensory information31.
While until now selective manipulation of mPFC outputs has not been shown to directly modulate social behavior6,48, Challis et al.6 have shown that mPFC–brainstem projections play a role in the induction of behavioral plasticity during social defeat. In this study, daily optogenetic activation or inhibition of mPFC terminals in the dorsal raphe nucleus immediately following social defeat blocked or precipitated social avoidance measured 24 h after the last defeat experience. Because mPFC neurons provide excitatory input to local GABAergic neurons that tonically inhibit serotonin neuron firing in the raphe nucleus (and thus control serotonin release across the brain)6, mPFC projections may have a dual role in regulating global neuromodulatory tone (via dorsal raphe) and behavior (via dPAG) to achieve adaptation to social threats. We note, however, that there are also key procedural differences between our study and Challis et al.6. In our social defeat procedure, mice were tested for social avoidance in the same context as that in which the aggression occurred, and thus our findings may be dependent to some degree on this aspect of classical contextual conditioning.
Both our cell-type-specific retrograde rabies tracing and ex vivo electrophysiology experiments identified Vglut2+ neurons as the major target of mPFC projections in dPAG (Fig. 6). Selective inhibition of Vglut2+ neurons in dPAG reduced social avoidance during presentation of the intruder (Fig. 7), and recent studies have shown that optogenetic activation of this population of cells evokes defensive behaviors37. Our discovery that the vast majority of these cells received presynaptic inhibitory inputs from mPFC provides a mechanism for the inhibitory effects of mPFC projections on c-Fos and social avoidance responses during exposure to an aggressor (Fig. 3). The absence of either direct or presynaptic mPFC modulation of Vgat+ neurons (Fig. 6 and Supplementary Fig. 6) and the absence of a behavioral effect of pharmacogenetic inhibition of this class of dPAG neurons was unexpected, but suggests that cortical modulation of dPAG did not substantially depend on feedforward GABAergic inhibition.
Evidence from neuroimaging studies suggests that the mPFC–dPAG circuit we describe is likely relevant for understanding prefrontal cortical control of human behavior. Direct projections between mPFC and dPAG have been described in primates49 and magnetic resonance imaging studies report a switch in brain activity from mPFC to dPAG during the prestrike phase in a pseudo-predator video game situation46, suggesting that reciprocal activity in these structures may be involved in anticipatory fear in humans. While our study was limited to males due to its reliance on inter-male aggression, mPFC–dPAG projections are conserved across sexes and are likely to control instinctive behavioral outputs also in females. Electrical stimulation of human dPAG elicits the sensation of being chased, supporting its role in mediating avoidance responses to threat50. Furthermore, our observation that the mPFC–dPAG pathway-dependent social avoidance induced by social defeat can be reversed by treatment with a single dose of ketamine (Supplementary Fig. 2), a potent antidepressant, suggests that this pathway may be a target of antidepressants and could serve as a neural substrate for the testing of antidepressant efficacy. Further work will be needed to identify the molecular mechanisms by which social experience remodels this pathway.
Methods
Animals.
C57BL/6J and CD-1 mice were obtained from local EMBL or EMMA colonies, or from Charles River Laboratories. CD-1 intruders were selected as aggressors if they attacked during the first 3 min after placement in the home cage of a novel C57BL/6J mouse across 3 consecutive days, as previously described51. These mice typically represented the most aggressive 15% of CD-1 mice tested. Vglut2::Cre52 and Gad2::Cre (Jackson Laboratory stock 019022) mice were used in a heterozygous state. Vglut2::Cre;RC::LSL-tdTomato (called Vglut2::tdTomato), Gad2::Cre;RC::LSL-tdTomato (called Gad2::tdTomato) and Vgat::Cre; RC::LSL-tdTomato (called Vgat-tdTomato) mice were obtained by crossing either the Vglut2::Cre line, Gad2::Cre line or Vgat::Cre line with Rosa26-CAG::loxP-STOP-loxP-tdTomato (Jackson Laboratory stock 007914). Vglut2::Cre;RC::LSL-EYFP (called Vglut2-EYFP) mice were obtained by crossing Vglut2::Cre (Jackson Laboratory stock 016963) with Rosa26-LSL-EYFP (Jackson Laboratory stock 006148). Thy1::GFP-M (ref. 53) mice were used in a homozygous state. Mice were maintained in a temperature- and humidity-controlled facility on a 12-h light–dark cycle (lights on at 7:00) with food and water provided ad libitum. All behavioral testing occurred during the animals' light cycle. All mice were handled according to protocols approved by the Italian Ministry of Health (#137/2011-B, #231/2011-B and #541/2015-PR) and commensurate with NIH guidelines for the ethical treatment of animals, except for during the in vitro electrophysiology experiments, which were conducted in the United Kingdom and were licensed under the United Kingdom Animals (Scientific Procedures) Act of 1986 following local ethical approval (Project License 70.7652).
Social defeat.
Mice were randomly assigned to either defeat or control groups. For defeat, singly-housed adult male mice (C57BL/6, 3–6 months old) were subjected to social defeat by placing an aggressive male CD-1 intruder mouse into the home cage of the experimental animal for 15 min each day. During the first 5 min the intruder was contained within a wire-mesh enclosure to prevent violent contact. Social approach and avoidance behavior, including number of investigations, investigation bout length, total time spent investigating and number of retreats (sudden movements away from the intruder) was quantified during the first 3 min of this anticipatory period (Observer XT 11, Noldus) by an experimenter blind to the treatment group. For defeated mice, the wire-mesh enclosure was removed, after which the intruder invariably attacked the resident repeatedly. Submissive (freezing and upright defensive postures) and exploration (rearing) behaviors of the resident and aggressive attacks of the intruder were quantified during the 10-min interaction period. Control animals were treated in the same manner, except that the wire mesh enclosure was not removed. This allowed control mice similar levels of visual, olfactory and auditory contact with the aggressor as defeated mice.
Social avoidance test.
Five to seven days after the last social defeat session, animals were subjected to a social interaction test in which an aggressive CD-1 intruder (or a novel female or object, where specified) was constrained within a wire-mesh enclosure placed into the home cage of the experimental animal. The animals were allowed to interact through the wire-mesh barrier for 5 min, and approach and avoidance behaviors were scored in the same way as during the anticipatory period of social defeat. For mPFC–dPAG projection inhibition, CNO was slowly infused via a single indwelling cannula (0.0015 mg, 0.15 μl; see below) immediately before testing. For Vglut2+ and Gad2+ dPAG inhibition, all mice were first tested under control conditions and then tested under the defeat condition. Seven d after the last control session, and 7 d after the last defeat session, CNO (3 mg/kg i.p.) or vehicle was systemically administered 45–60 min before testing. Testing consisted of a habituation session during which the experimental animal was allowed free exploration of their home cage for 5 min in the testing room, followed by the introduction of the intruder, behind a barrier, for a further 5 min.
Y-maze.
The Y-maze consisted of three gray, opaque plastic arms arranged at 120° angles around a center area. Animals were placed in a counterbalanced manner into one arm of the Y-maze and allowed to explore all arms of the maze for 8 min. Following a 2-min habituation period, the percentage of correct choices and same arm returns were assessed for 6 min. A correct choice was quantified as each time the mouse entered all three arms without returning to an arm previously entered. Same arm returns (SARs) counted the number of times a mouse entered fully into the center area and then returned to the arm they had just exited. Latency to exit the start arm and total distance traveled during the test were also quantified. Control and defeated mice were tested in the Y-maze 1 to 2 weeks after the last defeat session. Following the defeat treatment, mice either remained undisturbed or were injected with vehicle, 2.5 mg/kg ketamine or 5 mg/kg ketamine 1 d after the last defeat session. Defeated mice in one cohort of animals tested were not included in the ketamine experiment because the CD-1 mice used did not attack reliably. All injected mice tested in the Y-maze were also previously tested in the social avoidance test.
Elevated plus maze.
Mice were placed for 10 min on a four-arm plus maze made of two open and two closed arms (gray PVC, 30 cm × 6 cm) raised 50 cm above the ground. Manual scoring was done to quantify rearing and stretch attends in protected (body in closed arm) versus unprotected (body in open arm) areas as a measure of risk assessment. All elevated plus maze data was collected from implanted mice previously tested in the social avoidance test.
Tail suspension test.
Mice were suspended by their tail from a hook (43 cm from floor) for 6 min. A plastic cylinder was placed around the tail to prevent tail climbing. All tail suspension data was collected from implanted mice previously tested in the social avoidance test.
Stereotactic surgery.
Prior to surgery, mice were anesthetized with ketamine (100 mg/kg, i.p.) and xylazine (10 mg/kg, i.p.) and placed in a stereotactic frame (Kopf Instruments); isoflurane in oxygen was administered as needed to maintain anesthesia. For cholera-toxin-mediated retrograde tracing, the skull surface was exposed and mice were infused with 0.2 μl cholera toxin subunit B 0.5% (CTB647 and CTB555, Life Technologies) into dPAG (AP: −4.2; L: −1.18; from bregma DV: −2.36 from skull; angle: −26°) and into NAc (AP: +1.42 mm; L: −1.33 mm from bregma; DV: −3.5 mm from brain surface) using a glass capillary. In separate experiments Thy1::GFP (n = 8) or Gad2::Cre;RC::LSL-tdTomato (n = 1) mice were used. Serial coronal sections (250 μm, except in Gad2::Cre;RC::LSL-tdTomato, for which 50 μm was given) were cut on a vibratome and visualized using confocal microscopy. For mPFC–dPAG or mPFC–SuColl projection inhibition, the skull surface was exposed and mice were infused bilaterally with 0.2 μl of an adeno-associated virus expressing Venus and hM4D (AAV-Syn::Venus-2A-HAhM4D-WPRE)54 using a glass capillary filled with 1 μl of virus that was lowered unilaterally into the mPFC. After a 2-min delay, the capillary was retracted, and the contralateral mPFC was similarly infused. For local CNO delivery, a single 26-gauge stainless steel guide cannula (PlasticsOne) was implanted after viral infection into dPAG (AP: −4.16 mm; L: −1.0 mm; DV: −1.98 mm, angle: −26°; 1.25 mm projection from the pedestal), or into SuColl (AP: −4.1 mm; L: −0.75 mm; DV: −1.85 mm, angle: −30°, 0.20 mm projection from the pedestal) and secured to the skull using dental cement. For LFP recordings, the skull surface was exposed and two stainless steel watch screws were fixed permanently into the posterior and anterior portions of the skull, to serve as a ground and a reference, respectively. Teflon-coated tungsten wire electrodes were implanted unilaterally into PrL or Cg (AP: +1.65 mm; L: −0.50, DV: −1.50 mm from brain surface)55 and dPAG (AP: −4.16 mm; L: −1.32 mm, DV: −2.00 mm from brain surface, 26° lateral angle). Implanted electrodes were cemented directly to the skull with dental cement (DuraLay). For mPFC–dPAG and MDT–mPFC evoked potentials, animals were implanted unilaterally with bipolar stimulating electrodes into mPFC (AP: +1.72 mm, L: −0.40 mm, DV: −1.35 mm from brain surface) or MDT (AP: −1.2 mm, L: −0.40 mm, DV: −3.250 mm from brain surface) and a recording stereotrode into dPAG (AP: 4.1 mm, L: −1.3 mm, DV: −2.35 mm from skull surface, 26° lateral angle) or mPFC (AP: +1.72 mm, L: −0.40 mm, DV: −1.35 mm from brain surface) respectively. Electrodes were made of 50 μm Teflon-coated tungsten wires (Advent Research Materials) and were used for stimulation or recording purposes as needed. A 0.1-mm bare silver wire was affixed to a stainless steel watch screw fixed permanently in the skull as a ground. The wires were connected to two 3-pin sockets (Archer Connectors, M52). The connectors were fixed directly to the skull using acrylic resin (DuraLay) and connected to the Plexon system using a home-made adaptor. For rabies-mediated retrograde tracing, Vglut2::Cre and Gad2::Cre mice were infused into dPAG as described above with 0.1 μl AAV helper viruses that provided Cre-dependent expression of TVA and rabies protein G (AAV-EF1a::DIO-TVA-mCherry-WPRE and AAV-CAG::DIO-RabiesG-WPRE; from UNC Vector Core) followed 2–3 weeks later by infusion of an EnvA pseudotyped rabies virus in which the protein G gene was replaced by GFP (1 μl; Salk Institute Vector Core)32. AAV and rabies were both targeted toward the midline but injected unilaterally on opposite sides to avoid co-infection of the pipette tract. For cell-specific inhibition in dPAG, Vglut2::Cre or Gad2::Cre mice were infused 14 d before testing with 0.2 μl of AAV-expressed hM4D in a Cre-dependent manner (AAV-Syn::DIO-hM4DmCherry-WPRE; UNC Vector Core). Serial coronal sections (70 μm) were cut on a vibratome and visualized under a microscope to verify placement of all electrodes, cannulas and virus infections (Supplementary Figs. 3a and 4a). Only mice with appropriate placements were included in the reported data. For in vitro electrophysiology, Vglut2::Cre;RC::LSL-tdTomato or Vgat::Cre;RC::LSL-tdTomato male mice were injected bilaterally into mPFC (AP: +1.7; ML: ± 0.6; DV: −1.35) with 0.05 μL of AAV2-CamKIIa-hChR2(H134R)-EYFP virus (UNC Vector Core) delivered via manual hydraulic pump (Narishige). Following injection mice were allowed at least 2 weeks for viral expression.
In vivo electrophysiology.
All mice were allowed to recover for at least 7 d before testing and were habituated repeatedly for several days to the recording device by attaching a mock device of similar size and weight. LFP recordings were performed using a battery-powered custom wireless amplifier and recording device (23 × 15 × 13 mm, 3.7 g) located on the head of the animal56,57. LFP signals from electrodes located in mPFC and dPAG were sampled at 1,600 Hz (bandpass filter 1–700 Hz) and stored in the onboard 1-GB memory chip at 1,600 Hz58. A built-in accelerometer registered the movements of the animal throughout the experiment and an infrared detector on the device was used to synchronize electrophysiological and video recordings. For evoked potential recordings, the neural signal was amplified (gain 1,000×) and filtered (bandwidth of 0.1 Hz–10 kHz) through a headstage and a differential preamplifier (Omniplex, Plexon). Signals were digitized at 40 kHz and continuous recordings were collected for offline analysis. Synaptic field potentials in dPAG were evoked using a pulse generator (CS-420, Cibertec) and electrical stimulator (ISU-200bip, Cibertec) during home cage exploration and while the intruder was present in the home cage behind a barrier using a single 100-μs, square, biphasic (negative–positive) pulse applied to mPFC at a rate of 0.1 Hz. For each animal, the stimulus intensity was 40–50% of the intensity necessary for evoking a maximum fEPSP. Evoked potentials were monitored using an oscilloscope (Tektronix). At completion of the experiment, mice were anesthetized using 2.5% Avertin (400 mg/kg, i.p.; Sigma-Aldrich) and perfused transcardially (4.0% wt/vol paraformaldehyde, 0.1 M phosphate buffer, pH 7.4). For LFP recordings, a small electrolytic lesion was made around the tip of the electrode (0.4 mA, 3 s; Ugo Basile Lesion Making Device, Ugo Basile) before the animal was killed for analysis. Serial coronal sections (40 or 70 μm) were cut on a vibratome and visualized under a microscope to verify all electrode placements (Supplementary Fig. 3).
In vitro electrophysiology.
Acute coronal slices (200 μm) containing the PAG were prepared from 11–13 week old mice. Animals were killed by decapitation following isoflurane anesthesia. Coronal slices were cut at 4 °C using a 7000smz-2 vibrating microtome (Campden, UK). Brain slices were incubated at 37 °C for 1 h before being kept at room temperature (20–25 °C) before experiments in artificial cerebrospinal fluid (ACSF) containing 125 mM NaCl2, 2.5 mM KCl, 26 mM NaHCO3, 1.25 mM NaH2PO4, 25 mM glucose, 2 mM CaCl2, 1 mM MgCl2, 0.2% biocytin (pH 7.3 when bubbled with 95% O2 and 5% CO2). Borosilicate glass micropipettes with 3–6 MΩ resistance (Harvard Apparatus, UK) were filled with 136 mM K-gluconate, 4 mM KCl, 10 mM HEPES, 1 mM EGTA, 2 mM Na2ATP, 2 mM Mg2ATP, 0.5 mM Na2GTP, filtered (2 μm) before patching. Fluorescent cells were visualized on an upright Slicescope (Scientifica, UK) using a 60× objective and the relative coordinates of each neuron were recorded. Whole-cell patch-clamp recordings were achieved at room temperature, using a HEKA 800 Amplifier (HEKA, Germany). Data was acquired at 25 kHz using custom software. Channelrhodopsin was activated with widefield 490-nm LED illumination (CoolLED; 1-ms pulses). After electrophysiological recordings, slices were fixed in 4% paraformaldehyde for 15 min and incubated for 30 min in blocking solution containing 5% normal goat serum and 0.3 Triton X-100 in PBS, followed by chicken anti-GFP (1:1,000; Molecular Probes catalog #A10262)59 at 4 °C overnight. The slices were then washed with 0.3% Triton X-100 in PBS (PBS-T) for 3 × 10 min, incubated with secondary antibody (Alexa Fluor 488-conjugated goat anti-chicken IgGm 1:1,000; Molecular Probes catalog #A11039)60 for 1 h at room temperature, and after 2 × 10 min washes in PBS-T, they were incubated for 20 min in PBS-T with streptavidin (Alexa Fluor 635-conjugated streptavidin, 1:500; Molecular Probes catalog #S32364; see manufacturer's datasheet) to visualize biocytin-labeled neurons. After an additional 10 min wash in PBS, slices were mounted in Slow Fade mounting medium (Molecular Probes). Recorded slices with biocytin-filled neurons were imaged with 10× and 40× objectives on a Leica SP8 inverted confocal microscope (Leica). Deconvolution was performed using Huygens Software (Scientific Volume Imaging) and tiling of individual images was done in Fiji61.
Immunohistochemistry and immunofluorescence.
Immediately following social avoidance testing, mice were returned to their housing room for 90 min, deeply anesthetized with Avertin (400 mg/kg, i.p.; Sigma-Aldrich), perfused transcardially (4.0% wt/vol paraformaldehyde, 0.1 M phosphate buffer, pH 7.4) and brains were removed and postfixed overnight in 4.0% paraformaldehyde. The posterior half of the brain was cryoprotected (30% sucrose wt/vol, 0.1 M PBS, pH 7.4) at 4 °C overnight and flash frozen in isopentane. Coronal sections were taken with a sliding cryostat (40 μm; Leica Microsystems) and immunohistochemistry was performed. For c-Fos visualization, floating sections were incubated with anti-c-Fos antibody (1:10,000, Ab-5; Calbiochem; see manufacturer's datasheet)26,54 for 72 h at 4 °C, after which the primary antiserum was localized using the avidin–biotin complex system as per manufacturer's instructions (rabbit IgG, catalog #PK-6101, Vector Laboratories)62. The peroxidase complex was visualized by incubating slices for 5 min with a chromogenic solution consisting of 0.05% wt/vol 3,3′-diaminobenzidine tetrahydrochloride (Sigma-Aldrich), 6 μg/ml glucose oxidase (Sigma-Aldrich) and 0.4 mg/ml ammonium chloride in PBS, and then adding 2 mg/ml glucose to the solution. The reaction was stopped by extensive washing in PBS and sections were mounted, dehydrated and coverslipped with quick mounting medium (Eukitt, Fluka Analytical). c-Fos-immunopositive cells were counted using manual thresholding and automatic counting (ImageJ) in a section chosen randomly (bregma −4.16 mm) by an investigator blind to experimental treatment.
For visualization of HA-tagged hM4D, slices were mounted onto SuperPlus slides and allowed to dry. Slides were then boiled in citrate buffer (10 mM, pH 6.0) for 10 min and allowed to cool to room temperature before being submerged in PBS containing 0.4% Triton X-100 (PBS-T) for 1 h. They were then placed in blocking solution (1% BSA, 5% normal goat serum in PBS-T) at room temperature for 1 h, followed by incubation with a rabbit anti-HA mAb (C29F4, catalog #3724, Cell Signaling; see manufacturer's datasheet)63 at 1:500 in blocking buffer. Slides were exposed to secondary antibody (Alexa Fluor 546-conjugated goat anti-rabbit IgG, Invitrogen; see manufacturer's datasheet)64 in blocking buffer at room temperature for 90 min. Slides were then exposed to 4′,6-diamidino-2-phylindole, dihydrochloride65 (DAPI, catalog #D1306, Molecular Probes; see manufacturer's datasheet) at 1:1,000 in PBS at room temperature for 20 min. Slices were washed extensively with PBS between incubations and following DAPI staining.
Electrophysiology data analysis.
LFPs and synaptic field potentials were recorded on the first day and third day of social defeat during the anticipatory period. All differentials were changes calculated by comparing Day 3 to Day 1. LFP data were analyzed using Matlab (Mathworks) with the Chronux toolbox (see “Code availability,” below). To assess synchrony between LFP signals, coherence was calculated with the multitaper method, using a 200-ms window, time-bandwidth product (TW) of 5, and 9 tapers. Granger causality was performed as previously described66 and used an order of 20 estimated by a bivariate autoregressive model. fEPSP slopes were analyzed offline using commercial computer programs (Spike2 and SIGAVG, Cambridge Electronic Design) using the same rate period.
Statistical analyses.
Data analysis was performed using Statview (SAS) or Sigmaplot, except the in vitro electrophysiology data, which was analyzed in Python 2.7 using custom written software (see “Code availability,” below). All data are reported as mean ± s.e.m. Sample sizes were not predetermined using statistical methods, but all sample sizes were similar to previously reported behavioral, molecular and in vivo electrophysiological studies30,67. For all parametric tests, data distribution was assumed to be normal, but this was not formally tested. To measure statistical significance for differences in behavior between control and defeated mice, two-way or repeated measures ANOVAs followed by Fisher's PLSD post hoc testing (when appropriate) were performed. For experiments performed by multiple experimenters, outliers were defined as any data points greater than ± 2 s.d. from the mean and were removed. Two-tailed t-tests planned a priori were used to assess the effects of mPFC–dPAG inhibition separately in control and defeated mice. fEPSP data was analyzed using a repeated two-way ANOVA. For analysis of local field potential data, we used nonparametric Mann-Whitney U-tests as previously described to compare theta, beta and low-gamma coherence between control and defeated mice30. A Supplementary Methods Checklist is available.
Code availability.
In vivo electrophysiology data were analyzed using Matlab (Mathworks) with the Chronux toolbox (coherencyc, http://chronux.org/)68. In vitro electrophysiology data were analyzed using code available at https://github.com/ineuron/NeuroDAQ-Analysis.
Data availability.
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Change history
11 January 2017
In the version of this article initially published online, the annotation above the ChR2 bars in Figure 6k appeared as NNN instead of ***. Also, the third author's name was given as Zina Perova; the correct name is Zinaida Perova. The errors have been corrected in the print, PDF and HTML versions of this article.
References
Arnsten, A.F. Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 10, 410–422 (2009).
Takahashi, A., Nagayasu, K., Nishitani, N., Kaneko, S. & Koide, T. Control of intermale aggression by medial prefrontal cortex activation in the mouse. PLoS One 9, e94657 (2014).
Carrier, N. & Kabbaj, M. Sex differences in social interaction behaviors in rats are mediated by extracellular signal-regulated kinase 2 expression in the medial prefrontal cortex. Neuroscience 212, 86–92 (2012).
Wang, F. et al. Bidirectional control of social hierarchy by synaptic efficacy in medial prefrontal cortex. Science 334, 693–697 (2011).
Covington, H.E. III et al. Antidepressant effect of optogenetic stimulation of the medial prefrontal cortex. J. Neurosci. 30, 16082–16090 (2010).
Challis, C., Beck, S.G. & Berton, O. Optogenetic modulation of descending prefrontocortical inputs to the dorsal raphe bidirectionally bias socioaffective choices after social defeat. Front. Behav. Neurosci. 8, 43 (2014).
Goodson, J.L. The vertebrate social behavior network: evolutionary themes and variations. Horm. Behav. 48, 11–22 (2005).
Warden, M.R. et al. A prefrontal cortex-brainstem neuronal projection that controls response to behavioural challenge. Nature 492, 428–432 (2012).
Bossert, J.M. et al. Role of projections from ventral medial prefrontal cortex to nucleus accumbens shell in context-induced reinstatement of heroin seeking. J. Neurosci. 32, 4982–4991 (2012).
Faturi, C.B., Rangel, M.J. Jr., Baldo, M.V. & Canteras, N.S. Functional mapping of the circuits involved in the expression of contextual fear responses in socially defeated animals. Brain Struct. Funct. 219, 931–946 (2014).
Mobbs, D. et al. From threat to fear: the neural organization of defensive fear systems in humans. J. Neurosci. 29, 12236–12243 (2009).
Silva, B.A. et al. Independent hypothalamic circuits for social and predator fear. Nat. Neurosci. 16, 1731–1733 (2013).
Berton, O. et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 311, 864–868 (2006).
Price, J.S. & Sloman, L. The evolutionary model of psychiatric disorder. Arch. Gen. Psychiatry 41, 211 (1984).
Allen, N.B. & Badcock, P.B. Darwinian models of depression: a review of evolutionary accounts of mood and mood disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 30, 815–826 (2006).
Ottenbreit, N.D. & Dobson, K.S. Avoidance and depression: the construction of the cognitive-behavioral avoidance scale. Behav. Res. Ther. 42, 293–313 (2004).
Anand, A. et al. Antidepressant effect on connectivity of the mood-regulating circuit: an FMRI study. Neuropsychopharmacol. 30, 1334–1344 (2005).
Gabbott, P.L., Warner, T.A., Jays, P.R., Salway, P. & Busby, S.J. Prefrontal cortex in the rat: projections to subcortical autonomic, motor, and limbic centers. J. Comp. Neurol. 492, 145–177 (2005).
Floyd, N.S., Price, J.L., Ferry, A.T., Keay, K.A. & Bandler, R. Orbitomedial prefrontal cortical projections to distinct longitudinal columns of the periaqueductal gray in the rat. J. Comp. Neurol. 422, 556–578 (2000).
Lee, A.T., Vogt, D., Rubenstein, J.L. & Sohal, V.S. A class of GABAergic neurons in the prefrontal cortex sends long-range projections to the nucleus accumbens and elicits acute avoidance behavior. J. Neurosci. 34, 11519–11525 (2014).
Krishnan, V. et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 131, 391–404 (2007).
Wei, J., Bai, W., Liu, T. & Tian, X. Functional connectivity changes during a working memory task in rat via NMF analysis. Front. Behav. Neurosci. 9, 2 (2015).
Shiroma, P.R. et al. Neurocognitive performance and serial intravenous subanesthetic ketamine in treatment-resistant depression. Intl. J. Neuropsychopharmacol. 17, 1805–1813 (2014).
Vauterin, C. & Bazot, M. A double-blind controlled trial of amineptine versus trimipramine in depression. Curr. Med. Res. Opin. 6, 101–106 (1979).
Armbruster, B.N., Li, X., Pausch, M.H., Herlitze, S. & Roth, B.L. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. USA 104, 5163–5168 (2007).
Motta, S.C. et al. Dissecting the brain's fear system reveals the hypothalamus is critical for responding in subordinate conspecific intruders. Proc. Natl. Acad. Sci. USA 106, 4870–4875 (2009).
Fitzgerald, P.B., Laird, A.R., Maller, J. & Daskalakis, Z.J. A meta-analytic study of changes in brain activation in depression. Hum. Brain Mapp. 29, 683–695 (2008).
Seminowicz, D.A. et al. Limbic-frontal circuitry in major depression: a path modeling metanalysis. Neuroimage 22, 409–418 (2004).
Mayberg, H.S. et al. Reciprocal limbic-cortical function and negative mood: converging PET findings in depression and normal sadness. Am. J. Psychiatry 156, 675–682 (1999).
Likhtik, E., Stujenske, J.M., Topiwala, M.A., Harris, A.Z. & Gordon, J.A. Prefrontal entrainment of amygdala activity signals safety in learned fear and innate anxiety. Nat. Neurosci. 17, 106–113 (2014).
Northoff, G. & Sibille, E. Why are cortical GABA neurons relevant to internal focus in depression? A cross-level model linking cellular, biochemical and neural network findings. Mol. Psychiatry 19, 966–977 (2014).
Wall, N.R., Wickersham, I.R., Cetin, A., De La Parra, M. & Callaway, E.M. Monosynaptic circuit tracing in vivo through Cre-dependent targeting and complementation of modified rabies virus. Proc. Natl. Acad. Sci. USA 107, 21848–21853 (2010).
Vianna, D.M. & Brandão, M.L. Anatomical connections of the periaqueductal gray: specific neural substrates for different kinds of fear. Braz. J. Med. Biol. Res. 36, 557–566 (2003).
Spruston, N. Pyramidal neurons: dendritic structure and synaptic integration. Nat. Rev. Neurosci. 9, 206–221 (2008).
Petreanu, L., Huber, D., Sobczyk, A. & Svoboda, K. Channelrhodopsin-2-assisted circuit mapping of long-range callosal projections. Nat. Neurosci. 10, 663–668 (2007).
Haubensak, W. et al. Genetic dissection of an amygdala microcircuit that gates conditioned fear. Nature 468, 270–276 (2010).
Tovote, P. et al. Midbrain circuits for defensive behaviour. Nature 534, 206–212 (2016).
McCarthy, M.M., Pfaff, D.W. & Schwartz-Giblin, S. Midbrain central gray GABAA receptor activation enhances, and blockade reduces, sexual behavior in the female rat. Exp. Brain Res. 86, 108–116 (1991).
Mota-Ortiz, S.R., Sukikara, M.H., Felicio, L.F. & Canteras, N.S. Afferent connections to the rostrolateral part of the periaqueductal gray: a critical region influencing the motivation drive to hunt and forage. Neural Plast. 2009, 612698 (2009).
Lonstein, J.S. & Stern, J.M. Role of the midbrain periaqueductal gray in maternal nurturance and aggression: c-fos and electrolytic lesion studies in lactating rats. J. Neurosci. 17, 3364–3378 (1997).
Kvitsiani, D. et al. Distinct behavioural and network correlates of two interneuron types in prefrontal cortex. Nature 498, 363–366 (2013).
Karlsson, M.P., Tervo, D.G. & Karpova, A.Y. Network resets in medial prefrontal cortex mark the onset of behavioral uncertainty. Science 338, 135–139 (2012).
Behrens, T.E., Woolrich, M.W., Walton, M.E. & Rushworth, M.F. Learning the value of information in an uncertain world. Nat. Neurosci. 10, 1214–1221 (2007).
Lisboa, S.F., Camargo, L.H., Magesto, A.C., Resstel, L.B. & Guimaraes, F.S. Cannabinoid modulation of predator fear: involvement of the dorsolateral periaqueductal gray. Intl. J. Neuropsychopharmacolog. 17, 1193–1206 (2014).
Dielenberg, R.A., Leman, S. & Carrive, P. Effect of dorsal periaqueductal gray lesions on cardiovascular and behavioral responses to cat odor exposure in rats. Behav. Brain Res. 153, 487–496 (2004).
Mobbs, D. et al. When fear is near: threat imminence elicits prefrontal-periaqueductal gray shifts in humans. Science 317, 1079–1083 (2007).
Duman, R.S., Aghajanian, G.K., Sanacora, G. & Krystal, J.H. Synaptic plasticity and depression: new insights from stress and rapid-acting antidepressants. Nat. Med. 22, 238–249 (2016).
Christoffel, D.J. et al. Excitatory transmission at thalamo-striatal synapses mediates susceptibility to social stress. Nat. Neurosci. 18, 962–964 (2015).
An, X., Bandler, R., Ongür, D. & Price, J.L. Prefrontal cortical projections to longitudinal columns in the midbrain periaqueductal gray in macaque monkeys. J. Comp. Neurol. 401, 455–479 (1998).
Amano, K. et al. Endorphins and pain relief. Further observations on electrical stimulation of the lateral part of the periaqueductal gray matter during rostral mesencephalic reticulotomy for pain relief. Appl. Neurophysiol. 45, 123–135 (1982).
Tsankova, N.M. et al. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 9, 519–525 (2006).
Borgius, L., Restrepo, C.E., Leao, R.N., Saleh, N. & Kiehn, O. A transgenic mouse line for molecular genetic analysis of excitatory glutamatergic neurons. Mol. Cell. Neurosci. 45, 245–257 (2010).
Feng, G. et al. Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron 28, 41–51 (2000).
Silva, B.A. et al. Independent hypothalamic circuits for social and predator fear. Nat. Neurosci. 16, 1731–1733 (2013).
Paxinos, G. & Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates (Academic Press, San Diego, 2001).
Vyssotski, A.L. et al. EEG responses to visual landmarks in flying pigeons. Curr. Biol. 19, 1159–1166 (2009).
Vyssotski, A.L. et al. Miniature neurologgers for flying pigeons: multichannel EEG and action and field potentials in combination with GPS recording. J. Neurophysiol. 95, 1263–1273 (2006).
Zhan, Y. et al. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat. Neurosci. 17, 400–406 (2014).
Suzuki, Y., Kiyokage, E., Sohn, J., Hioki, H. & Toida, K. Structural basis for serotonergic regulation of neural circuits in the mouse olfactory bulb. J. Comp. Neurol. 523, 262–280 (2015).
Szulwach, K.E. et al. Cross talk between microRNA and epigenetic regulation in adult neurogenesis. J. Cell Biol. 189, 127–141 (2010).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Hsu, S.M. & Raine, L. Protein A, avidin, and biotin in immunohistochemistry. J. Histochem. Cytochem. 29, 1349–1353 (1981).
Field, J. et al. Purification of a RAS-responsive adenylyl cyclase complex from Saccharomyces cerevisiae by use of an epitope addition method. Mol. Cell. Biol. 8, 2159–2165 (1988).
Nishiguchi, K.M. et al. Gene therapy restores vision in rd1 mice after removal of a confounding mutation in Gpr179. Nat. Commun. 6, 6006 (2015).
Kubista, M., Akerman, B. & Nordén, B. Characterization of interaction between DNA and 4′,6-diamidino-2-phenylindole by optical spectroscopy. Biochemistry 26, 4545–4553 (1987).
Zhan, Y. Theta frequency prefrontal-hippocampal driving relationship during free exploration in mice. Neuroscience 300, 554–565 (2015).
Vialou, V. et al. DeltaFosB in brain reward circuits mediates resilience to stress and antidepressant responses. Nat. Neurosci. 13, 745–752 (2010).
Mitra, P. & Bokil, H. Observed Brain Dynamics (Oxford University Press, Oxford; New York, 2008).
Acknowledgements
We thank F. Zonfrillo for animal husbandry and P. Heppenstall (Mouse Biology Unit, EMBL) for providing the Vglut2::Cre mouse line. Funding was provided by EMBL (C.T.G., T.B.F.), the ERC Advanced Grant “Corefear” (C.T.G.), the Swiss National Science Foundation Advanced Fellows Program (T.B.F.), the Wellcome Trust/Royal Society Henry Dale Fellowship (098400/Z/12/Z, T.B.) a Medical Research Council (MRC) grant MC-UP-1201/1 (T.B.), Marie Sklodowska-Curie grant no. 659842 (Z.P.), the University of Zurich, Forschungskredit (A.L.V.) and the One Hundred Talents Program of CAS and funding from the Shenzhen city government (JCYJ20140901003938992 and KQCX2015033117354153) (Y.Z.).
Author information
Authors and Affiliations
Contributions
T.B.F. designed, performed and analyzed all experiments, except the retrograde tracer experiments, which were designed, performed and analyzed by L.M.; the in vitro electrophysiology experiments, which were designed, performed and analyzed by Z.P. and T.B.; the monosynaptic rabies experiment, which was designed, performed and analyzed by B.A.S.; the evoked field potential experiments, which were designed, performed and analyzed by M.E.M.; the Granger causality and power analyses, which were carried out by Y.Z.; and some behavioral experiments and imaging, which were performed and analyzed by A.K., V.V., L.G., A.H. and S.P. The AAV-Syn::Venus-2A-HAhM4D virus was packaged and tested by V.G. and A.I. The wireless recording device was built by A.L.V. The project was conceived and the manuscript written by T.B.F and C.T.G., with critical input from T.B.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Social, anxiety-like and depressive-like behaviors in defeated and control mice.
(a) Social avoidance (n=9; day, F[6,8] = 5.81, p = 0.0001) and (b) number of attacks (n=8) across social defeat sessions during the period in which the intruder was prevented from attacking the resident by an enclosure. (c) Retreats made by defeated (n=7) and control (n=7) mice in response to aggressors, females or a novel object (defeat: F[1,12] = 6.5, p = 0.026, stimulus: F[2,12]=3.48, p=0.047). (d) Time spent in the open arm (control, n=18; defeat, n=18), (e) number of unprotected and protected stretch attends (control, n=8; defeat, n=8), and (f) rearing in defeated and control mice in the elevated plus maze (control, n=8; defeat, n=8). (g) Immobility in defeated and control mice in the tail suspension test (control, n=9; defeat, n=9). a, b, red squares represent mean of defeated mice. c-g, grey circles represent individual control mice, light red squares represent individual defeated mice, horizontal bar denotes mean. All error bars represent standard error of the mean.
Supplementary Figure 2 Behavioral effects of ketamine on defeated and control mice.
(a) Timeline showing behavioral testing and ketamine/vehicle administration. (b) Time spent investigating (defeat: F[1, 57] = 65.8, p < 0.0001; defeat x ketamine: F[2,57] = 4.3, p = 0.018; control/vehicle, n=13; control/2.5, n=8; control/5, n=15; defeat/vehicle, n=8; defeat/2.5, n=8; defeat/5, n=11), (c) duration of investigation bout (control/vehicle, n=14; control/2.5, n=8; control/5, n=14; defeat/vehicle, n=9; defeat/2.5, n=8; defeat/5, n=10), (d) retreats (defeat: F[1,56] = 31.9, p < 0.0001; defeat x treatment: F[2, 56]= 5.9, P=0.0048; control/vehicle, n=14; control/2.5, n=7; control/5, n=14; defeat/vehicle, n=9; defeat/2.5, n=8; defeat/5, n=10) and (e) baseline locomotor behavior (control/vehicle, n=12; control/2.5, n=4; control/5, n=14; defeat/vehicle, n=8; defeat/2.5, n=4; defeat/5, n=9) in control and defeated mice given vehicle or ketamine. (f) Same arm returns (SARs) (control/vehicle, n=13; control/2.5, n=7; control/5, n=11; defeat/vehicle, n=8; defeat/2.5, n=7; defeat/5, n=9; defeat x drug: F[2, 49] = 5.56, P=0.0067), (g) spontaneous alternation (control/vehicle, n=14; control/2.5, n=8; control/5, n=12; defeat/vehicle, n=8; defeat/2.5, n=8; defeat/5, n=9), (h) latency to exit the start arm (control/vehicle, n=13; control/2.5, n=7; control/5, n=12; defeat/vehicle, n=8; defeat/2.5, n=8; defeat/5, n=8), and (i) overall distance (control/vehicle, n=14; control/2.5, n=8; control/5, n=12; defeat/vehicle, n=8; defeat/2.5, n=8; defeat/5, n=9) travelled in the Y-maze. 0.05<+p < 0.1, *p < 0.05; **p < 0.01; ***p < 0.001. b-i, grey circles represent individual control mice, light red squares represent individual defeated mice, horizontal bar denotes mean, error bars represent standard error of the mean. veh, vehicle.
Supplementary Figure 3 mPFC infection sites for mPFC–dPAG inhibition and behavioral effects of mPFC–SuColl inhibition in control mice.
Area of viral infection (AAV-Syn::Venus-2A-HAhM4D-WPRE) visualized by endogenous Venus in mPFC of (a) control and (b) defeated mice administered CNO. (c) Distance travelled in the home cage in (left) control and (right) defeated mice after intra-PAG administration of CNO or vehicle (control/vehicle, n=10; control/CNO, n=10; defeat/vehicle, n=10; defeat/CNO, n=12). (d-g) Schematic describing bilateral infection of the mPFC with AAV expressing Venus fluorescent protein and HA-tagged hM4D (AAV-Syn::Venus-2A-HA-hM4D), and implantation of a guide cannula over SuColl. (d) Time investigating the aggressor (vehicle, n=10; CNO, n=10), (e) investigation bouts (vehicle, n=10; CNO, n=10), (f) retreats (vehicle, n=10; CNO, n=10) and (g) overall activity (vehicle, n=8; CNO, n=6) in control mice administered CNO or vehicle prior to testing. (h) Quantification of c-Fos immunopositive cells in ventrolateral (vl) PAG (control/vehicle, n=10; control/CNO, n=10; defeat/vehicle, n=10; defeat/CNO, n=12). c-h, grey circles represent individual control mice, light red squares represent individual defeated mice, horizontal bar denotes mean. All error bars represent standard error of the mean. veh, vehicle.
Supplementary Figure 4 Effects of social defeat on mPFC–dPAG functional connectivity when distal to the aggressor.
(a) Electrode placements for LFPs recorded in mPFC and dPAG of control (grey) and defeated (red) mice. (b, c) Relative coherence (coherence differential) in defeated (n=8) and control (n=8) mice when distal to the aggressor. (d) Relative causality (causality differential) in the dPAG->mPFC and mPFC-> dPAG direction in defeated (n=8) and control (n=8) mice when distal to the aggressor (U=8, p=0.038). Power spectra differential (e, f) mPFC and (g, h) PAG of defeated (n=8) and control (n=8) mice when distal to the aggressor (beta, U=6, p=0.0047). Power spectra were averaged across mice. Power in each frequency band was calculated as the sum of the power values. *P<0.05; **P<0.01. c, d, f, h, grey circles represent individual control mice, light red squares represent individual defeated mice, horizontal bar denotes mean. All error bars represent standard error of the mean.
Supplementary Figure 5 Extracellular synaptic field potentials in dPAG and MDT of defeated and control mice.
Superimposed recordings illustrating extracellular synaptic field potentials recorded at (a) dPAG following electrical stimulation of mPFC and at (b) mPFC following electrical stimulation of MDT along the different sessions. (gray scale: control group; red scale: defeated group).
Supplementary Figure 6 Cell-specific retrograde labeling in GAD2::Cre mice and location of Vglut2+ neurons used for whole cell recordings.
(a) Summary of rabies-infected neurons (GFP+, mCherry-) in the forebrain of VGgat::Cre animals (percentage of the average number of retrograde neurons weighted to the number of starter cells present in each animal, n=7). No neurons were found in the olfactory bulb (grey), hippocampus (grey), or cortex. Areas not counted (midbrain and hindbrain) are indicated in white. Thal – thalamus; Hypothal – hypothalamus. (b) Schematic showing the location of all Vglut2+ neurons from which whole-cell recordings were made. Blue circles represent neurons with monosynaptic inputs from the PFC, and orange circles are neurons without PFC input. Aq – cerebral aqueduct; dmPAG – dorsomedial PAG; dlPAG – dorsolateral PAG; lPAG – lateral PAG; vlPAG – ventrolateral PAG; DR – dorsal raphe.
Supplementary Figure 7 Acquisition of social defeat in Vglut2::Cre mice and behavioral effects of inhibition of GAD2+ cells in the dPAG.
(a) Time spent investigating, (b) mean investigation bout, and (c) retreats in Cre+ and Cre- defeated and control mice during the three days of acquisition. (d) Time spent investigating (vehicle, n=6; CNO, n=7), (e) mean duration of investigation (vehicle, n=6; CNO=7) and (f) retreats (vehicle, n=6; CNO, n=7) after systemic administration of vehicle or CNO in GAD2::Cre mice infected with AAV-Syn::DIO-hM4D-mCherry in the dPAG (t(11)=2.2, p=0.016). a-c, black circles represent mean of control mice, red squares represent mean of defeated mice. d-f, grey circles represent individual control mice, light red squares represent individual defeated mice, horizontal bar denotes mean. All error bars represent standard error of the mean.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–7 and Supplementary Tables 1–3 (PDF 1822 kb)
Rights and permissions
About this article

Cite this article
Franklin, T., Silva, B., Perova, Z. et al. Prefrontal cortical control of a brainstem social behavior circuit. Nat Neurosci 20, 260–270 (2017). https://doi.org/10.1038/nn.4470
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nn.4470
This article is cited by
-
A prefrontal-thalamic circuit encodes social information for social recognition
Nature Communications (2024)
-
A distinct cortical code for socially learned threat
Nature (2024)
-
Premotor projections from the locus coeruleus and periaqueductal grey are altered in two rat models with inborn differences in emotional behavior
Experimental Brain Research (2024)
-
Enriched Environment Enhances Sociability Through the Promotion of ESyt1-Related Synaptic Formation in the Medial Prefrontal Cortex
Molecular Neurobiology (2024)
-
Somatostatin neurons in prefrontal cortex initiate sleep-preparatory behavior and sleep via the preoptic and lateral hypothalamus
Nature Neuroscience (2023)
