
Abstract
The hygiene hypothesis postulates that the recent increase in allergic diseases such as asthma and hay fever observed in Western countries is linked to reduced exposure to childhood infections. Here we investigated how infection with a gammaherpesvirus affected the subsequent development of allergic asthma. We found that murid herpesvirus 4 (MuHV-4) inhibited the development of house dust mite (HDM)-induced experimental asthma by modulating lung innate immune cells. Specifically, infection with MuHV-4 caused the replacement of resident alveolar macrophages (AMs) by monocytes with regulatory functions. Monocyte-derived AMs blocked the ability of dendritic cells to trigger a HDM-specific response by the TH2 subset of helper T cells. Our results indicate that replacement of embryonic AMs by regulatory monocytes is a major mechanism underlying the long-term training of lung immunity after infection.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
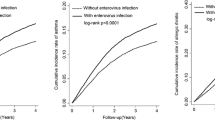

References
Brodin, P. et al. Variation in the human immune system is largely driven by non-heritable influences. Cell 160, 37–47 (2015).
Carr, E.J. et al. The cellular composition of the human immune system is shaped by age and cohabitation. Nat. Immunol. 17, 461–468 (2016).
Netea, M.G. et al. Trained immunity: A program of innate immune memory in health and disease. Science 352, aaf1098 (2016).
Lambrecht, B.N. & Hammad, H. The immunology of the allergy epidemic and the hygiene hypothesis. Nat. Immunol. 18, 1076–1083 (2017).
Lambrecht, B.N. & Hammad, H. The immunology of asthma. Nat. Immunol. 16, 45–56 (2015).
Beura, L.K. et al. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 532, 512–516 (2016).
Webley, W.C. & Aldridge, K.L. Infectious asthma triggers: time to revise the hygiene hypothesis? Trends Microbiol. 23, 389–391 (2015).
Nilsson, C. et al. Does early EBV infection protect against IgE sensitization? J. Allergy Clin. Immunol. 116, 438–444 (2005).
Saghafian-Hedengren, S., Sverremark-Ekström, E., Linde, A., Lilja, G. & Nilsson, C. Early-life EBV infection protects against persistent IgE sensitization. J. Allergy Clin. Immunol. 125, 433–438 (2010).
Balfour, H.H. Jr. et al. Age-specific prevalence of Epstein-Barr virus infection among individuals aged 6-19 years in the United States and factors affecting its acquisition. J. Infect. Dis. 208, 1286–1293 (2013).
Condon, L.M. et al. Age-specific prevalence of Epstein-Barr virus infection among Minnesota children: effects of race/ethnicity and family environment. Clin. Infect. Dis. 59, 501–508 (2014).
Dowd, J.B., Palermo, T., Brite, J., McDade, T.W. & Aiello, A. Seroprevalence of Epstein-Barr virus infection in U.S. children ages 6-19, 2003-2010. PLoS One 8, e64921 (2013).
Cesarman, E. Gammaherpesviruses and lymphoproliferative disorders. Annu. Rev. Pathol. 9, 349–372 (2014).
Virgin, H.W., Wherry, E.J. & Ahmed, R. Redefining chronic viral infection. Cell 138, 30–50 (2009).
Barton, E.S. et al. Herpesvirus latency confers symbiotic protection from bacterial infection. Nature 447, 326–329 (2007).
Reese, T.A. et al. Sequential infection with common pathogens promotes human-like immune gene expression and altered vaccine response. Cell Host Microbe 19, 713–719 (2016).
MacDuff, D.A. et al. Phenotypic complementation of genetic immunodeficiency by chronic herpesvirus infection. eLife 4, 4 (2015).
Barton, E., Mandal, P. & Speck, S.H. Pathogenesis and host control of gammaherpesviruses: lessons from the mouse. Annu. Rev. Immunol. 29, 351–397 (2011).
Hammad, H. et al. House dust mite allergen induces asthma via Toll-like receptor 4 triggering of airway structural cells. Nat. Med. 15, 410–416 (2009).
Cadwell, K. The virome in host health and disease. Immunity 42, 805–813 (2015).
Fowler, P., Marques, S., Simas, J.P. & Efstathiou, S. ORF73 of murine herpesvirus-68 is critical for the establishment and maintenance of latency. J. Gen. Virol. 84, 3405–3416 (2003).
Plantinga, M. et al. Conventional and monocyte-derived CD11b+ dendritic cells initiate and maintain T helper 2 cell-mediated immunity to house dust mite allergen. Immunity 38, 322–335 (2013).
Hammad, H. et al. Inflammatory dendritic cells--not basophils--are necessary and sufficient for induction of Th2 immunity to inhaled house dust mite allergen. J. Exp. Med. 207, 2097–2111 (2010).
van Helden, M.J. & Lambrecht, B.N. Dendritic cells in asthma. Curr. Opin. Immunol. 25, 745–754 (2013).
Guilliams, M. et al. Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nat. Rev. Immunol. 14, 571–578 (2014).
Mesnil, C. et al. Resident CD11b+Ly6C− lung dendritic cells are responsible for allergic airway sensitization to house dust mite in mice. PLoS One 7, e53242 (2012).
Holt, P.G., Haining, S., Nelson, D.J. & Sedgwick, J.D. Origin and steady-state turnover of class II MHC-bearing dendritic cells in the epithelium of the conducting airways. J. Immunol. 153, 256–261 (1994).
Hussell, T. & Bell, T.J. Alveolar macrophages: plasticity in a tissue-specific context. Nat. Rev. Immunol. 14, 81–93 (2014).
Bedoret, D. et al. Lung interstitial macrophages alter dendritic cell functions to prevent airway allergy in mice. J. Clin. Invest. 119, 3723–3738 (2009).
Holt, P.G. et al. Downregulation of the antigen presenting cell function(s) of pulmonary dendritic cells in vivo by resident alveolar macrophages. J. Exp. Med. 177, 397–407 (1993).
Lauzon-Joset, J.F., Marsolais, D., Langlois, A. & Bissonnette, E.Y. Dysregulation of alveolar macrophages unleashes dendritic cell-mediated mechanisms of allergic airway inflammation. Mucosal Immunol. 7, 155–164 (2014).
Lawler, C., Milho, R., May, J.S. & Stevenson, P.G. Rhadinovirus host entry by co-operative infection. PLoS Pathog. 11, e1004761 (2015).
Spinelli, L., Carpentier, S., Montañana Sanchis, F., Dalod, M. & Vu Manh, T.P. BubbleGUM: automatic extraction of phenotype molecular signatures and comprehensive visualization of multiple Gene Set Enrichment Analyses. BMC Genomics 16, 814 (2015).
van de Laar, L. et al. Yolk sac macrophages, fetal liver, and adult monocytes can colonize an empty niche and develop into functional tissue-resident macrophages. Immunity 44, 755–768 (2016).
Gibbings, S.L. et al. Transcriptome analysis highlights the conserved difference between embryonic and postnatal-derived alveolar macrophages. Blood 126, 1357–1366 (2015).
Askenase, M.H. et al. Bone-marrow-resident NK cells prime monocytes for regulatory function during infection. Immunity 42, 1130–1142 (2015).
Guilliams, M. & Scott, C.L. Does niche competition determine the origin of tissue-resident macrophages? Nat. Rev. Immunol. 17, 451–460 (2017).
Shi, C. & Pamer, E.G. Monocyte recruitment during infection and inflammation. Nat. Rev. Immunol. 11, 762–774 (2011).
Scott, C.L. et al. Bone marrow-derived monocytes give rise to self-renewing and fully differentiated Kupffer cells. Nat. Commun. 7, 10321 (2016).
Guilliams, M. et al. Alveolar macrophages develop from fetal monocytes that differentiate into long-lived cells in the first week of life via GM-CSF. J. Exp. Med. 210, 1977–1992 (2013).
Hashimoto, D. et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity 38, 792–804 (2013).
Schneider, C. et al. Induction of the nuclear receptor PPAR-γ by the cytokine GM-CSF is critical for the differentiation of fetal monocytes into alveolar macrophages. Nat. Immunol. 15, 1026–1037 (2014).
Sabatel, C. et al. Exposure to bacterial CpG DNA protects from airway allergic inflammation by expanding regulatory lung interstitial macrophages. Immunity 46, 457–473 (2017).
Grainger, J.R. et al. Inflammatory monocytes regulate pathologic responses to commensals during acute gastrointestinal infection. Nat. Med. 19, 713–721 (2013).
Peacock, J.W. & Bost, K.L. Murine gammaherpesvirus-68-induced interleukin-10 increases viral burden, but limits virus-induced splenomegaly and leukocytosis. Immunology 104, 109–117 (2001).
Siegel, A.M., Herskowitz, J.H. & Speck, S.H. The MHV68 M2 protein drives IL-10 dependent B cell proliferation and differentiation. PLoS Pathog. 4, e1000039 (2008).
van Rijt, L.S. et al. Essential role of dendritic cell CD80/CD86 costimulation in the induction, but not reactivation, of TH2 effector responses in a mouse model of asthma. J. Allergy Clin. Immunol. 114, 166–173 (2004).
Busse, W.W., Lemanske, R.F. Jr. & Gern, J.E. Role of viral respiratory infections in asthma and asthma exacerbations. Lancet 376, 826–834 (2010).
Blaskovic, D., Stanceková, M., Svobodová, J. & Mistríková, J. Isolation of five strains of herpesviruses from two species of free living small rodents. Acta Virol. 24, 468 (1980).
Machiels, B., Stevenson, P.G., Vanderplasschen, A. & Gillet, L. A gammaherpesvirus uses alternative splicing to regulate its tropism and its sensitivity to neutralization. PLoS Pathog. 9, e1003753 (2013).
Latif, M.B. et al. Deletion of murid herpesvirus 4 ORF63 affects the trafficking of incoming capsids toward the nucleus. J. Virol. 90, 2455–2472 (2015).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Gentleman, R.C. et al. Bioconductor: open software development for computational biology and bioinformatics. Genome Biol. 5, R80 (2004).
Acknowledgements
We thank U. Eriksson (Center for Molecular Cardiology, University of Zurich) for BALB/c CD45.1+ genitor mice; A. Osterhaus, T. Marichal and C. Desmet for critical discussions; and L. Dams, C. Delforge, E. Deglaire, C. Espert, A. Guillaume, M. Sarlet and A. Vanderlinden for technical and secretary assistance. Supported by the University of Liège (VIR-IMPRINT ARC), “Fonds de la Recherche Scientifique - Fonds National Belge de la Recherche Scientifique” (“credit de recherche” J007515F; “projet de recherche” T.0195.16; research associate support for B.D.) and Institut MERIEUX (starting grant).
Author information
Authors and Affiliations
Contributions
B.M., M.D. and L.G. designed the experiments with the help of H.H., M.G., B.N.L. and F.B.; B.M. and M.D. did most of the experiments and compiled the data; B.M., M.D., X.X. and L.G. prepared the figures; X.X. performed the transcriptomic and statistical analyses; J.J., C.M., C.S., F.L., and P.M. were involved in specific experiments; B.M., M.D., D.D., H.H., M.G., B.D., A.V., B.N.L., F.B. and L.G. analyzed the data; and B.M., M.D. and L.G. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Infection with MuHV-4 protects both BALB/c and C57BL/6 mice against HDM-induced allergic asthma.
(a) Experimental design of HDM-induced high dose model of asthma in BALB/c mice. Mice were mock-infected or infected intranasally with MuHV-4 (1x104 PFU in 50 μL PBS). Thirty days post-infection (p.i.), animals received 3 sequential intranasal instillations of saline or 100 μg HDM extracts (day 0, 7, 14) before euthanasia and airway allergy evaluation 3 days later (day 17). (b) Experimental design of HDM-induced low dose model of asthma in BALB/c or C57BL/6. Thirty days p.i. as in a, animals received intranasal saline instillation or were sensitized with 10 μg HDM (day 0). One week later, mice were challenged with 5 daily intranasal instillations of 10 μg HDM (days 7 to 11) before euthanasia and airway allergy evaluation 3 days after the last instillation (day 14). (c-g) Total and differential cell counts of BALF cells (c,e), ELISA measurement of cytokine release by MLN cells following ex vivo restimulation with HDM (d,f), and ELISA Measurement of HDM-specific IgE, IgG1 and IgG2c levels in sera (g) at euthanasia of mock or MuHV-4-infected BALB/c (c-d) or C57BL/6 mice (e-g), submitted 30 days p.i. to the HDM-induced low dose model of asthma (as in b). *** p< 0.001, ** p< 0.01 and * p < 0.05 (Two-way ANOVA and Tukey's multiple comparison test (c-e) or Student's t-test (g)). Data are representative of 2 independent experiments with 5 mice per group (mean ± s.e.m. in c-g).
Supplementary Figure 2 Infection with MuHV-4 impairs HDM-induced airway allergy in young mice.
(a-d) Quantification of MuHV-4 genomic copies in splenic cells by qPCR (a) and of MuHV-4 specific immunoglobulins in sera by ELISA (b), histological analysis of lung sections (scale bars, 100 μm) (c), total and differential cell counts of BALF cells (d), at euthanasia of mock or MuHV-4-infected 3 weeks old BALB/c mice, submitted 30 days p.i. to the HDM-induced high dose model of asthma. Data are mean ± s.e.m. of samples from 5 mice per group. *** p< 0.001, ** p< 0.01 and * p < 0.05 (Two-way (a, b) or One-way (d) ANOVA and Tukey's multiple comparison test).
Supplementary Figure 3 Infection with MuHV-4 induces a long-term protection against HDM-induced airway allergy.
(a-d) Quantification of MuHV-4 specific immunoglobulins in sera by ELISA (a), total and differential cell counts of BALF cells (b), histological analysis of lung sections (scale bars, 100 μm) (c), and ELISA measurement of cytokine release by MLN cells following ex vivo restimulation by HDM (d), at euthanasia of mock or MuHV-4-infected BALB/c mice, submitted 7, 30 or 60 days p.i. to the HDM-induced high dose model of asthma. *** p< 0.001, ** p< 0.01 and * p < 0.05 (Two-way ANOVA and Tukey's multiple comparison test). Data are representative of two independent experiments with 5 mice per group (mean ± s.e.m. in a, b, d).
Supplementary Figure 4 Establishment of MuHV-4 latency is not necessary to allow protection against HDM-induced airway allergy.
(a-d) Quantification of MuHV-4 specific immunoglobulins in sera by ELISA (a), quantification of MuHV-4 genomic copies in splenic cells by qPCR (b), histological analysis of lung sections (scale bars, 100 μm) (c), and total and differential cell counts of BALF cells (d) at euthanasia of mock or MuHV-4-infected BALB/c mice either with the WT strain or with latency-deficient viral mutants (FS73 and Del73 strains) or with a corresponding revertant (Rev73), submitted 30 days p.i. to the HDM-induced high dose model of asthma. *** p< 0.001, ** p< 0.01 and * p < 0.05 (Two-way ANOVA and Tukey's multiple comparison test). Data are representative of 2 independent experiments with 5 mice per group (mean ± s.e.m. in a,b,d).
Supplementary Figure 5 Migratory DC subsets in MLNs from mock- and MuHV-4 infected mice after HDM sensitization.
(a) Gating strategy for CD103+ cDC1, CD11b+ cDC2, lung-derived CCR7+ DC populations in the MLN of mock or MuHV-4 infected BALB/c mice, HDM sensitized (100 μg) 30 days p.i. and euthanized 2 days later. MLN cells from MuHV-4 infected mice were labelled with CFSE and then mixed with MLN cells from mock-infected mice prior to antibody staining and flow cytometry analysis of a single mix, allowing unbiased comparisons. Debris and doublets were excluded based on FSC and SSC. MOs were excluded based on Ly6c and CD64 expression. Migratory lung-derived CD11b+ cDC2 were identified as liveCD11c+MHC-IIhiCCR7+CD11b+ cells. Migratory lung-derived CD103+ cDC1s were identified as liveCD11c+MHC-IIhiCCR7+CD103+CD11blo cells. Representative flow cytometry plots are shown with the mean frequency of the different cells subsets. (b) Mean fluorescence intensities of maturation markers (CD40, CD80 and CD86, with independent staining and analysis for each of these markers) and MHCII by migratory DCs subsets were compared between the CFSE+ (originating from MuHV-4 infected mice) and CFSE- (originating from mock-infected mice) populations. A reciprocal experiment comparing CFSE+ DCs from mock-infected mice to CFSE- DCs from MuHV-4 infected has been performed as control and gave similar results (not shown).
Supplementary Figure 6 Intranasal infection with MuHV-4 protects mice against HDM-induced airway allergy, but intraperitoneal infection does not.
(a-d) Quantification of MuHV-4 genomic copies in splenic cells by qPCR (a), quantification of MuHV-4 specific immunoglobulins in sera by ELISA (b), total and differential cell counts of BALF cells (d), and ELISA measurement of cytokine release by MLN cells following ex vivo restimulation by HDM (d) at euthanasia of mock, or MuHV-4 infected BALB/c mice either intranasally or intraperitoneally (1x104 PFU), submitted 30 days p.i. to the HDM-induced high dose model of asthma. ** p< 0.01 (Mann-Whitney t-test (a, b) or one-way ANOVA and Tukey's multiple comparison test (c, d)). Data are representative of 2 independent experiments with 5 mice per group (mean ± s.e.m. in a-d).
Supplementary Figure 7 Details of the transcriptomics analysis of AMs.
(a) Sequence and mapping statistics for raw Illumina data. (b) Validation of sample purity by assessing the expression of lineage-restricted marker genes for potential contaminants; eosinophils, neutrophils, B and T cells. (c) Unsupervised, hierarchical clustering of individual lanes demonstrating discrete clustering of biologic replicates. (d) Summary of differentially expressed (DE) genes (P<1e-5) in each pairwise comparison showing DE genes in blue in volcano plot, showing the total number of DE genes outside the bidirectional arrows, and showing in the arrowheads the direction of upregulated expression for all, moderately (log 2-fold change ± 2-4) and highly (log 2-fold change > 4) DE genes.
Supplementary Figure 8 Gating strategy for AM and MO subsets in BM, blood and BALF following infection with MuHV-4.
(a-c) Flow plots on MO subsets in BM, gated as liveCD19-CD11b+SiglecF-Ly6G-Ly6C+ cells (a), on MO subsets in blood gated as live CD11b+Ly6G-CD19-SiglecF-Ly6C+ cells (b), on AM defined as liveautofluorescent+CD11chi cells (c) and BALF MO gated as nonautofluorescentCD11cloCD19-CD11b+Ly6G-CCR2+Ly6C+ further analyzed for MHCII and Sca-1 expression (c). (d) Representative flow cytometry overlays of AM (as defined above) from BALB/c mock-infected mice (red) and AM (green) and MOs (blue) from MuHV-4 infected mice isolated from BALF at the different times p.i.. (e,f) Flow cytometry quantification of AM viability in BALF at different times p.i. using Annexin V-APC/7-AAD staining. Representative flow cytometry plots (e) and quantification of Annexin+/7AAD- and Annexin+/7AAD+ cells among AM (f) from mock or MuHV-4-infected BALB/c mice at different times p.i.. Data are mean ± s.e.m. of samples from 5 mice per group. *** p< 0.001, ** p< 0.01 and * p < 0.05 (in (f), all data were compared to values obtained at day 0 by one-way ANOVA and Dunnett's multiple comparison test).
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–8 and Supplementary Tables 1 and 2 (PDF 2596 kb)
Rights and permissions
About this article

Cite this article
Machiels, B., Dourcy, M., Xiao, X. et al. A gammaherpesvirus provides protection against allergic asthma by inducing the replacement of resident alveolar macrophages with regulatory monocytes. Nat Immunol 18, 1310–1320 (2017). https://doi.org/10.1038/ni.3857
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.3857
