
Abstract
The hedgehog and Wnt signaling pathways play important roles in human cancers with possible interaction. This study aimed at analysis and correlation of the expression of Gli1, a transcriptional factor and target gene of hedgehog signaling pathway, with clinicopathological parameters and expression of β-catenin, an important member of the Wnt pathway, in normal, hyperplastic and malignant endometrium. Immunohistochemical study on 15 normal endometrium, 14 simple and complex hyperplasia without atypia, 37 atypical complex hyperplasia and 80 endometrial cancers showed significant Gli1 overexpression and β-catenin nuclear immunoreactivity in endometrial cancers and atypical endometrial hyperplasia when compared with normal endometrium (P<0.05). Overexpression of Gli1 in endometrial cancers correlated with well-differentiated histological grade (P<0.001), non-myometrial invasion (P=0.004) and superficial myometrial invasion (P=0.041). β-Catenin nuclear immunoreactivity was also associated with well-differentiated histology (P=0.013). Gli1 overexpression positively correlated with β-catenin nuclear immunoreactivity in atypical complex hyperplasia (P=0.013) and endometrial carcinoma (P=0.017). Similar Gli1 and β-catenin protein expression pattern was observed in normal and endometrial cancer cell lines by western blotting. We further showed a complex formation between Gli1 and β-catenin protein in endometrial cancer cell lines in an immunoprecipitation study. Ectopic overexpression of Gli1 into endometrial cancer cells led to reduced expression of β-catenin in cell cytoplasm and increased expression of β-catenin in the nuclei. In summary, overexpression of Gli1 was an early event in endometrial carcinogenesis. Aberrant activation of hedgehog pathway may play important roles in endometrial cancer through β-catenin nuclear accumulation.
Similar content being viewed by others

Main
Endometrial cancer is the most common malignancy of the female genital tract worldwide,1 and its occurrence has risen significantly in Asian cities.2 On the basis of morphology, clinical behavior and epidemiology, endometrial cancer was classified into two major types: the estrogen-dependent type I endometrioid adenocarcinoma that comprises 70–80% of endometrial cancers and the type II non-endometroid cancers, which is estrogen-independent, including papillary serous adenocarcinoma and clear-cell adenocarcinoma.3 Common molecular mechanisms involved in type I endometrioid cancer include PTEN and K-ras gene mutations, microsatellite instability and aberrant β-catenin expression4 whereas p53 mutation and high expression of Her2/neu oncogene are two major genetic alterations observed in type II endometrial cancer. In Hong Kong, the increase in the incidence of endometrial cancer is particularly significant in type I cancers (unpublished data on cases managed in Queen Mary Hospital).
β-Catenin is crucial in cell–cell adhesion through complex formation with E-cadherin and is an important member of the Wnt signal pathway.5 In the Wnt/β-catenin pathway, the free (cytoplasmic) β-catenin is demolished intracellularly in the absence of Wnt ligand. β-catenin is translocated to the nucleus either in the presence of Wnt ligand or β-catenin gene mutation, leading to the disruption of the β-catenin degradation.6 β-catenin mutation is present in about 20% of endometrioid cancers.7 Although microsatellite instability, PTEN and K-ras mutations frequently coexist with each other, these molecular abnormalities are seldom seen in tumors with β-catenin alterations.8
Aberrant activation of the hedgehog signaling pathway has been described in a variety of human cancers, including those of the skin,9 prostate,10 pancreas11 and gastrointestinal tract.12 This pathway controls different aspects of tissue patterning, cell proliferation, differentiation and regeneration, including the stem cell population in adult tissues.13 In mammals, three hedgehog ligands including Sonic hedgehog (Shh), Indian hedgehog (Ihh) and Desert hedgehog (Dhh) have been identified. The hedgehog pathway is inactive in the absence of ligands. Patched, a transmembrane receptor, acts as a tumor suppressor that inhibits Smoothened (Smo) from activating downstream signaling molecules and subsequent Gli-mediated gene transcription. Through binding of any one of the three ligands to Patched, the pathway is activated by alleviating Patched-mediated suppression of Smo. The continued increased endogenous expression of hedgehog (ligand-dependent) or mutations of Patched, Smo and Supressor of fused (Sufu) (ligand-independent) can lead to aberrant activation of hedgehog pathway.14
Ihh has been recently shown to antagonize Wnt signaling in colonic epithelial cell differentiation.15 A reverse association between Gli1 immunoreactivity and β-catenin nuclear accumulation was observed in colon cancer.16 However, other studies reported a conflicting finding of enhancing interaction between hedgehog and Wnt pathway.17, 18 Although the overexpression of some hedgehog molecules in endometrial cancers and hyperplasias has been reported recently,19 the interaction between Gli1 and β-catenin is unexplored. This study aimed at evaluating the expression of the transcriptional factor Gli1 in normal, hyperplastic and cancerous endometrium in association with β-catenin and various clinicopathological features. In vitro interaction between Gli1 and β-catenin was further analyzed.
Materials and methods
Clinical Samples
Formalin-fixed, paraffin-embedded tissues of 80 endometrial cancers, 15 samples of normal endometrium (7 proliferative, 7 secretory and 1 interval phase), 14 hyperplastic endometrium (7 simple hyperplasia and 7 complex hyperplasia) as well as 37 atypical complex hyperplasia (15 pure atypical hyperplasia and 22 atypical hyperplasia with metachronous endometrial cancer) were retrieved for immunohistochemistry. All specimens were collected from the Department of Pathology, the University of Hong Kong, Queen Mary Hospital. Histological classification was assessed according to the established criteria.20 Among the 80 endometrial cancers, there were 68 endometrioid adenocarcinomas, 8 papillary serous adenocarcinomas and 4 clear cell carcinomas. Patients’ age ranged from 20 to 82 years. There were 56 early stage (stage I) and 24 late-stage (stage II–IV) cancers. The mean follow-up period was 87 months (range: 7–168 months).
Cell Lines
Human endometrial cancer cell lines HEC-1A, HEC-1B and RL95-2 were purchased from American Type Culture Collection, Manassas, VA, USA. A normal endometrial cell line, extended from primary culture of normal endometrial cells from one premenopausal female around 40 years old with regular 28-day menstrual cycles, was generously provided by Professor Doris Benbrook, University of Oklahoma Health Sciences Center.21 HEC-1B and the normal human endometrial cell line (NEM) were maintained in minimum essential medium. HEC-1A was maintained in McCoy's 5A medium (Invitrogen, Carlsbad, CA, USA). RL95-2 was maintained in Dulbecco's modified Eagle's medium/Ham's F12 medium with 0.005 mg/ml insulin (Sigma chemicals co. St Louis, MO, USA). The medium was supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. Cells were harvested for mRNA and protein extraction.
Immunohistochemistry
In all, 5-μm thick sections of formalin-fixed, paraffin-embedded tissues were deparaffinized. Antigen retrieval for Gli1and β-catenin staining was performed by heating for 5 min in a pressure cooker using 10 mM EDTA buffer (pH8.0) and in a microwave oven at 95°C using 0.01 M citrate buffer (pH 6.0), respectively.22 The sections were treated with 3% hydrogen peroxide and incubated with 10% normal serum to block non-specific binding. They were then incubated with primary antibodies: anti-Gli1 (sc-20687, 1:50, Santa Cruz Biotechnology, Santa Cruz, CA, USA) and anti-β-catenin (Clone 14, 1:1000, BD Transduction Laboratory, Lexington, KY, USA) at 4°C overnight. Secondary antibodies (Gli1, goat antirabbit and β-catenin, rabbit antimouse) were added for 30 min at room temperature. 3,3-diaminobenzidine-hydrogen peroxide was used as chromogen. Sections were then lightly counterstained with hematoxylin. A case of prostate cancer and a case of endometrial cancer known to be positive for β-catenin and Gli110 were used as positive control, whereas substitution of primary antibody by TBS was used as a negative control.
Assessment of Immunohistochemical Staining
Immunoreactivity of Gli1 in cytoplasm was scored as follows: 0 (none), 1+ (weak/moderate) and 2+ (intense) according to their staining intensity. Gli1 overexpression was identified for cases with staining intensity of 2+. Nuclear β-catenin staining was evaluated as the proportion (%) of carcinoma cells with distinct nuclear β-catenin staining compared with total cancers cells. More than 500 carcinoma cells were counted in each section. Cases were considered positive when at least 5% tumor cells had nuclear staining.
Transient Transfection of Gli1
pcDNA3.1-HisB-hGli1 was a gift from Dr Hiroshi Sasaki (Riken Center for Developmental Biology, Kobe, Japan).23 Transient transfection of Gli1 into HEC-1B cells was carried out using GeneJuice Transfection Reagent (Novagen, Darmstadt, Germany) (plasmid:genejuice=1:3) according to the manufacturer's instructions. Gli1 mRNA expression was confirmed by quantitative real-time RT-PCR. Cells with ectopic expression of Gli1 were harvested for protein analysis.
Subcellular Protein Extraction
Cytoplasmic and nuclear extraction from HEC-1B cells with ectopic Gli1 expression were performed using the NE-PER Nuclear and cytoplasmic extraction Reagents (Pierce biotechnology, Rockford, IL, USA) at 4°C.24 Briefly, Transfected cells with Gli1 or empty vector were collected by centrifugation for 3 min at 500 g after removal of the medium and washed twice with cold PBS. The pellet was resuspended in 200 μl of ice-cold CER I with protease inhibitors and incubated on ice for 10 min. After 11 μl of ice-cold CER II were added and vortexed, the supernatant (cytoplasmic extact) was immediately transferred to a prechilled tube following centrifugation at maximum speed for 5 min at 4°C. The insoluble pellet fraction was saved for nuclear extracts and added in 100 μl of ice-cold NER. Then, the sample was placed on the ice and continued vortexing for 15 s every 10 min for total 40 min. The nuclear suspension was centrifuged at maximum speed for 10 min, and the supernatant (nuclear extract) was immediately collected.
Immunoprecipitation
Total cell lysate from HEC-1A and RL95-2 cells was prepared in immunoprecipitation lysis buffer containing protease inhibitors. Protein concentrations were measured using a protein assay kit (Bio-Rad) according to the manufacturer's recommendation. In all, 2 μg β-catenin antibodies (BD transduction) were added into 500 μg total cellular protein and incubated overnight at 4°C. Then, 20 μl protein A/G PLUS-Agarose (sc-2003, Santa Cruz) were added and incubated for 4 h at 4°C on a rotator. The antibody-bound beads were collected through centrifugation and washed four times with PBS. Finally, the immunoprecipitated proteins were resuspended with SDS sample buffer by boiling for 3 min and subjected to western blotting. Lysate that was not incubated with antibody served as controls.
Western blotting
Quantified total cell lysates, cytoplasmic and nuclear extracts and the immunoprecipitated proteins were separated in 7.5 or 10% SDS-polyacrylamide gel, and transferred to polyvinylidene difluoride membrane (Amersham Biosciences, Piscataway, NJ, USA). The membrane was blotted with 5% non-fat milk, and probed with Gli1, α-tubulin and histone H1 (Santa Cruz Biotechnology), β-catenin (BD transduction)25 and β-actin (Sigma-Aldrich, St Louis, MO, USA). After washing, the membranes were incubated with appropriate horseradish peroxidase–conjugated secondary antibody (Santa Cruz) and detected by the enhanced chemiluminescence plus system (Amersham Biosciences).
Statistical Analysis
Correlations among the expression of Gli1 and β-catenin were analyzed by Spearman's test. The difference of Gli1 and β-catenin expression in categories with different clinical parameters was tested by χ2- or Fisher's exact tests. The protein expression after Gli1 transfection in western blotting was compared to that with empty vector by student t-test. All statistical tests were two-sided. Differences were considered significant if P<0.05. Survival curves were calculated by the Kaplan–Meier method and the significance was analyzed by the log–-rank test. All analyses were carried out by the use of SPSS 13.0 for windows (SPSS Inc., Chicago, IL, USA).
Results
Gli1 Protein Overexpression is an Early Event in Endometrial Carcinogenesis
By immunohistochemistry, all 15 normal endometrial samples, including those in proliferative, secretory phase or interval phase showed either weak or moderate Gli1 expression (1+). The Gli1 staining was observed mainly in the cytoplasm with inconspicuous nuclear staining. Three of the 14 (21%) hyperplastic endometrium without atypia and 17 of the 37 (46%) atypical hyperplastic endometrium showed Gli1 overexpression (2+; Figure 1), whereas the others showed 1+ expression. There was thus significant increased expression of Gli1 in endometrium with atypical complex hyperplasia (P=0.001) compared with that of normal endometrium. No difference was found between Gli1 overexpression in hyperplastic endometrium without atypia and that of normal endometrium (P=0.10) between Gli1 overexpression in atypical complex hyperplasia and that in endometrial hyperplasia without atypia (P=>0.05), as well as between the overexpression of Gli1 in atypical hyperplasia and that in endometrial cancers (P=>0.05; Figure 1, Table 1).

Gli1 and β-catenin expression was assessed in endometrial tissues by immunohistochemistry. Photomicrographs were taken with assistance of Aperio Spectrum (magnification: 9.4 × 5.0, inset: 20 × 20): (a) normal endometrium with weak Gli1 (1+) expression and membranous β-catenin expression; (b) endometrium with atypical complex hyperplasia showing increased Gli1 overexpression (2+) and β-catenin nuclear expression; (c) endometrioid adenocarcinoma with weak Gli1 (1+) expression displayed no nuclear immunoreactivity for β-catenin; (d) endometrioid adenocarcinoma with Gli1 overexpression (2+) showed distinct β-catenin nuclear expression (inset).
In endometrial cancer group, although Gli1 staining was still observed mainly in the cytoplasm, nuclear staining was sporadically present in endometrial cancer cells as reported earlier by Feng et al.19 In 80 cases of endometrial cancer, 26 (33%), 12 (15%) and 42 (53%) cancers were scored as 0, 1 and 2, respectively (Figure 1). Overexpression of Gli1 (2+) was found to be correlated significantly with well-differentiated histology (P=<0.001), non-myometrial invasion (P=0.004) and superficial myometrial invasion (invasion <50%; P=0.041). There was no significant correlation between Gli1 expression and histological type, clinical stage, cervical infiltration, vascular invasion, patients’ age and survival (P=>0.05; Table 1).
Nuclear Expression of β-Catenin Correlated with Better Differentiation of Endometrial Cancer
Membranous staining of β-catenin was present in all 15 cases of normal endometrium, 51 hyperplastic endometrium and 69 cases of endometrial cancers. In contrast, distinct nuclear β-catenin immunoreactivity (>5%) was seen in 18 of 80 (23%) endometrial cancers, 12 of the 37 (32%) hyperplastic endometrium with atypia, one of the 14 (7%) hyperplastic endometrium without atypia, but in none of the normal endometrium. The percentage of cells with nuclear β-catenin expression varied from 5 to 70%. There was significantly higher proportion of nuclear immunoreactivity for β-catenin in complex hyperplastic endometrium with atypia compared with that in normal endometrium (P=0.011; Figure 1). No difference was found between nuclear β-catenin expression in atypical hyperplastic endometrium and that in hyperplasia without atypia (P=>0.05) as well as between that in atypical endometrial hyperplasia and in endometrial cancers (P=>0.05).
Among the 18 endometrial cancers with positive nuclear β-catenin staining, squamous metaplasia was present in five cases, in which the squamous morules always showed β-catenin nuclear staining. Marginally, significant correlation between β-catenin nuclear staining and the endometrioid histologic type was observed as 18 of the 68 (27%) endometrioid cancers had nuclear immunoreactivity, whereas none of 12 non-endometrioid cancers did (P=0.059). In other words, all the cancers with positive β-catenin nuclear staining were of endometrioid type. Nuclear β-catenin staining was found to correlate with low-grade cancers (P=0.013) and younger age of patients (P=0.041), but not with clinical stage, myometrial invasion, vascular invasion, cervical infiltration and patients’ survival (P=>0.05; Table 1).
Gli1 Protein Overexpression Correlated with β-Catenin Nuclear Expression
All cases with β-catenin nuclear expression in endometrial cancers were seen in tumors with a score of 2 or 1 for Gli1 expression, whereas none of the tumors with negative Gli1 expression had β-catenin nuclear staining. There was a significant correlation between Gli1 overexpression and β-catenin nuclear staining in endometrial cancers (P=0.017), type I endometrioid cancer (P=0.001), and in 37 cases of atypical complex endometrial hyperplasia (P=0.013), but not in type II endometrial cancer (P=>0.05).
Gli1 and β-Catenin Protein Expression in Endometrial Cancer Cell Lines
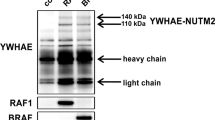
By western blotting, the Gli1 protein expression pattern in cell lines was found to concur with its expression in clinical samples (Figure 2a). Increased β-catenin expression was shown in cancer cell lines compared with normal endometrial cells (Figure 2a).

(a) Western blotting showed the heterogeneous Gli1 expression and increased β-catenin expression in endometrial cancer cell lines when compared with normal endometrial cells (NEM). (b) Immunoprecipitation study showed that Gli1 protein physically binds to β-catenin in HEC-1A and RL95-2. HEC-1B cells were used as positive control; (c) ectopic overexpression of Gli1 by transient transfection in HEC-1B cells showed decreased expression of β-catenin in the cytoplasm, but increased expression of β-catenin in the nucleus compared with cells with empty vector; V, empty vector; Gli1, pcDNA3.1-HisB-hGli1. The right bar chart showed a significant difference in their β-catenin expression level (P=<0.001). (d) Molecules that form interacting complexes with β-catenin. Besides Glil, β-catenin can also bind to E-cadherin, Sufu, Tcf, axin, adenomatous polyposis coli (APC) protein and glycogen synthase kinase 3 (GSK3) in Wnt pathway. Dashed line indicates indirect effects and crossbar represents repression; β-cat, β-catenin.
Gli1 and β-Catenin Protein Formed a Complex and Ectopic Overexpression of Gli1 in Endometrial Cancer Cells Enhanced β-Catenin Nuclear Accumulation
To further explore the mechanisms underlying the correlation between Gli1 and β-catenin, immunoprecipitation was performed on protein lysates of HEC-1A and RL95-2 cells that expressed endogenous Gli1 and β-catenin. β-catenin monoclonal antibody was applied followed by detection of the coimmunoprecipitate with an anti-Gli1 antibody. Western blotting confirmed an interaction between Gli1 and β-catenin proteins (Figure 2b). Furthermore, ectopic overexpression of Gli1 by transient transfection in HEC-1B cells induced increased expression of β-catenin in cancer cell nuclei, but decreased the expression of β-catenin in cell cytoplasm (P<0.001; Figure 2c).
Discussion
Gli1 is a transcriptional factor and is considered as one of the indicators for Hedgehog pathway activation.26 By immmunohistochemistry, overexpression of Gli1 was observed in endometrial cancer when compared with that in normal endometrium. Western blotting showed similar expression pattern in endometrial cell lines. Such findings are in keeping with that reported by Feng et al.19 We also found that Gli1 overexpression in endometrial cancers correlated with favorable clinicopathological index, such as well-differentiated tumor histology, absent or superficial myometrial invasion. In addition, overexpression of Gli1 was observed in atypical endometrial hyperplasia, but not in normal endometrium, suggesting that the overexpression of Gli1 may be an early event in endometrial tumorigenesis. However, overexpression of Gli1 was also observed in half of the type II cancers (6/12), which usually are high-grade cancers and deeply invasive. This may be related to the small number of type II cancer used in the study. Alternatively, the correlation between Gli1 overexpression and favorable clinicopathological parameters might be confined to type I endometrioid cancer and does not apply to type II cancers. More cases of type II cancers are needed to clarify this situation.
Earlier reports showed that β-catenin expression was significantly correlated with type I endometrioid cancers,27 and β-catenin gene mutation had an association with the early onset of cancer,28 low-grade histology and absence of lymph node metastasis in endometrial cancers.7 In our current study, β-catenin nuclear expression was only found in atypical complex hyperplastic endometrium and endometrial cancers, but not in normal endometrium and hyperplastic endometrium without atypia. Moreover, nuclear β-catenin expression correlated with well-differentiated tumor, confirming that β-catenin nuclear expression is an early event in endometrial tumorigenesis. Our study also confirmed a marginal association between β-catenin nuclear immunoreactivity with histological type.
We then correlated the protein expression of Gli1 and β-catenin in endometrial cancers and their precursors, atypical hyperplastic endometria. Both hedgehog and Wnt pathways are known to regulate embryonic growth and patterning, as well as to control the stem cell number in a postembryonic epithelium.6 The interaction between hedgehog and Wnt pathways is complicated. Ihh can suppress Wnt–β-catenin signaling in colonic epithelial differentiation.15 Gli1 overexpression was found to inversely correlate with β-catenin nuclear expression in gastric and colon cancers.16, 29 Ectopic overexpression of Gli1 inhibited proliferation of gastric and colon cancer cells, as well as suppressed Wnt transcriptional activity and nuclear β-catenin accumulation in gastric cancer cells.16, 29 However, in the epidermis, activation of β-catenin leads to increased expression of Shh.17 Exogenous β-catenin can enhance Gli1 transcriptional activity in vitro in human gastric, colon and lung cancer cells.18 In our immunostaining study, overexpression of Gli1 in endometrioid cancers and atypical hyperplastic endometria was found to be positively correlated with β-catenin nuclear expression. However, in type II cancers, whereas 50% (6/12) of the cases showed Gli1 overexpression, none of them had nuclear β-catenin expression, suggesting that Gli1 and β-catenin interaction might adopt divergent pathways in the development of the two types of endometrial cancers with distinct genetic characteristics.
We further showed a complex formation between Gli1 and β-catenin in endometrial cancer cells with endogenous expression of both Gli1 and β-catenin protein in an immunoprecipitation study. Earlier study has actually shown complex formation between β-catenin and suppressor of fused (Sufu), a negative regulator of the hedgehog signaling pathway that controls the nuclear-cytoplasmic distribution of Gli through direct protein–protein interactions. The complex functions as a negative regulator of T-cell factor (Tcf) transcription, a molecule in Wnt pathway.30 Moreover, ectopic overexpression of Gli1 in endometrial cancer cells induced increased nuclear β-catenin accumulation and decreased cytoplasmic expression of β-catenin, suggesting that overexpression of Gli1 may induce relocalization of β-catenin from cytoplasm to the nucleus. Such findings are in agreement with the findings of Li et al31 that Gli1 induced relocalization of β-catenin to the nucleus through E-cadherin and Snail. In animal model of mice skin, uncontrolled hedgehog signaling dependent on Wnt/β-catenin signaling has been found to be sufficient to drive basal cell carcinoma or basal cell carcinoma such as tumorigenesis.32 Nuclear β-catenin in human basal cell carcinoma is correlated with increased proliferation.33 Our immunohistochemical and immunoprecipitation results suggest that Gli1 overexpression might be directly involved in the proliferation of endometrial cancer cells through inducing nuclear accumulation of β-catenin of Wnt pathway in the tumorigenesis. As β-catenin can bind to E-cadherin, Sufu and Tcf, axin, adenomatous polyposis coli protein and glycogen synthase kinase 3 in Wnt pathway with different functions (Figure 2d),34 the underlying mechanisms for their competition binding to β-catenin and the roles in carcinogenesis need further study.
In conclusion, our study showed that the overexpression of Gli1 protein was related to nuclear accumulation of β-catenin in atypical complex hyperplasia of endometrium and endometrial cancers. Overexpression of Gli1 correlates with favorable clinicopathological parameters in cancer group, indicating that Gli1 protein overexpression might be an early event in endometrial carcinogenesis. Furthermore, we showed that complex formation between Gli1 and β-catenin and ectopic overexpression of Gli1 may function in inducing β-catenin relocalization from cytoplasm to the nucleus in endometrial cancer cells. This finding may provide theoretical supports for the prevention and treatment of endometrial cancer by antagonizing the interaction of Gli1 with β-catenin.
References
Ries L, Melbert D, Krapcho M, et al. SEER Cancer Statistics Review, 1975–2004. National Cancer Institute: Bethesda, MD, 2007.
Hospital Authority. Hong Kong Cancer Statistics, Hong Kong Cancer Registry 2006.
Bokhman JV . Two pathogenetic types of endometrial carcinoma. Gynecol oncol 1983;15:10–17.
Hecht JL, Mutter GL . Molecular and pathologic aspects of endometrial carcinogenesis. J Clin Oncol 2006;24:4783–4791.
Watt FM . Unexpected Hedgehog-Wnt interactions in epithelial differentiation. Trends Mol Med 2004;10:577–580.
Taipale J, Beachy PA . The Hedgehog and Wnt signalling pathways in cancer. Nature 2001;411:349–354.
Saegusa M, Hashimura M, Yoshida T, et al. beta-Catenin mutations and aberrant nuclear expression during endometrial tumorigenesis. Br J Cancer 2001;84:209–217.
Palacios J, Catasus L, Moreno-Bueno G, et al. Beta- and gamma-catenin expression in endometrial carcinoma. Relationship with clinicopathological features and microsatellite instability. Virchows Arch 2001;438:464–469.
Xie J, Murone M, Luoh SM, et al. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature 1998;391:90–92.
Sanchez P, Hernandez AM, Stecca B, et al. Inhibition of prostate cancer proliferation by interference with SONIC HEDGEHOG-GLI1 signaling. Proc Natl Acad Sci USA 2004;101:12561–12566.
Kayed H, Kleeff J, Keleg S, et al. Indian hedgehog signaling pathway: expression and regulation in pancreatic cancer. Int J Cancer 2004;110:668–676.
Lees C, Howie S, Sartor RB, et al. The hedgehog signalling pathway in the gastrointestinal tract: implications for development, homeostasis, and disease. Gastroenterology 2005;129:1696–1710.
Stecca B, Mas C, Ruiz i Altaba A . Interference with HH-GLI signaling inhibits prostate cancer. Trends Mol Med 2005;11:199–203.
Evangelista M, Tian H, de Sauvage FJ . The hedgehog signaling pathway in cancer. Clin Cancer Res 2006;12:5924–5928.
van den Brink GR, Bleuming SA, Hardwick JC, et al. Indian Hedgehog is an antagonist of Wnt signaling in colonic epithelial cell differentiation. Nat Genet 2004;36:277–282.
Akiyoshi T, Nakamura M, Koga K, et al. Gli1, downregulated in colorectal cancers, inhibits proliferation of colon cancer cells involving Wnt signalling activation. Gut 2006;55:991–999.
Alonso L, Fuchs E . Stem cells in the skin: waste not, Wnt not. Genes Dev 2003;17:1189–1200.
Maeda O, Kondo M, Fujita T, et al. Enhancement of GLI1-transcriptional activity by beta-catenin in human cancer cells. Oncol Rep 2006;16:91–96.
Feng YZ, Shiozawa T, Miyamoto T, et al. Overexpression of hedgehog signaling molecules and its involvement in the proliferation of endometrial carcinoma cells. Clin Cancer Res 2007;13:1389–1398.
Ronnett BM, Zaino RJ, Ellenson LH, et al. Endometrial carcinoma. In: Kurman RJ (ed). Blaustein's Pathology of the Female Genital Tract. Springer-Verlag: New York, 2002, pp 501–559.
Kamelle S, Sienko A, Benbrook DM . Retinoids and steroids regulate menstrual phase histological features in human endometrial organotypic cultures. Fertil Steril 2002;78:596–602.
Liao X, Siu MK, Chan KY, et al. Hypermethylation of RAS effector related genes and DNA methyltransferase 1 expression in endometrial carcinogenesis. Int J Cancer 2008;123:296–302.
Sasaki H, Nishizaki Y, Hui C, et al. Regulation of Gli2 and Gli3 activities by an amino-terminal repression domain: implication of Gli2 and Gli3 as primary mediators of Shh signaling. Development 1999;126:3915–3924.
Chan HY, Siu MK, Zhang HJ, et al. Activated Stat3 expression in gestational trophoblastic disease: correlation with clinicopathological parameters and apoptotic indices. Histopathology 2008;53:139–146.
Siu MK, Wong ES, Chan HY, et al. Overexpression of NANOG in gestational trophoblastic diseases: effect on apoptosis, cell invasion, and clinical outcome. Am J Pathol 2008;173:1165–1172.
Kubo M, Nakamura M, Tasaki A, et al. Hedgehog signaling pathway is a new therapeutic target for patients with breast cancer. Cancer Res 2004;64:6071–6074.
Moreno-Bueno G, Hardisson D, Sanchez C, et al. Abnormalities of the APC/beta-catenin pathway in endometrial cancer. Oncogene 2002;21:7981–7990.
Ikeda T, Yoshinaga K, Semba S, et al. Mutational analysis of the CTNNB1 (beta-catenin) gene in human endometrial cancer: frequent mutations at codon 34 that cause nuclear accumulation. Oncol Rep 2000;7:323–326.
Yanai K, Nakamura M, Akiyoshi T, et al. Crosstalk of hedgehog and Wnt pathways in gastric cancer. Cancer Lett 2008;263:145–156.
Meng X, Poon R, Zhang X, et al. Suppressor of fused negatively regulates beta-catenin signaling. J Biol Chem 2001;276:40113–40119.
Li X, Deng W, Lobo-Ruppert SM, et al. Gli1 acts through Snail and E-cadherin to promote nuclear signaling by beta-catenin. Oncogene 2007;26:4489–4498.
Hoseong Yang S, Andl T, Grachtchouk V, et al. Pathological responses to oncogenic Hedgehog signaling in skin are dependent on canonical Wnt/beta-catenin signaling. Nat Genet 2008;40:1130–1135.
Saldanha G, Ghura V, Potter L, et al. Nuclear beta-catenin in basal cell carcinoma correlates with increased proliferation. Br J Dermatol 2004;151:157–164.
Kalderon D . Similarities between the Hedgehog and Wnt signaling pathways. Trends Cell Biol 2002;12:523–531.
Acknowledgements
We thank Dr Hiroshi Sasaki at Riken Center for Developmental Biology, Kobe, Japan for providing us pcDNA3.1-HisB-hGli1 and Professor Doris Benbrook, University of Oklahoma Health Sciences Center, for the NEM cell line. The study was supported by the University of Hong Kong Conference and Research Grant.
Author information
Authors and Affiliations
Corresponding author
Additional information
Disclosure/conflict of interest
This study has no conflicts of interest (both financial and personal).
Rights and permissions
About this article
Cite this article
Liao, X., Siu, M., Au, C. et al. Aberrant activation of hedgehog signaling pathway contributes to endometrial carcinogenesis through β-catenin. Mod Pathol 22, 839–847 (2009). https://doi.org/10.1038/modpathol.2009.45
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/modpathol.2009.45
Keywords
This article is cited by
-
Glioblastoma stem cells and Wnt signaling pathway: molecular mechanisms and therapeutic targets
Chinese Neurosurgical Journal (2020)
-
β-Catenin-Gli1 interaction regulates proliferation and tumor growth in medulloblastoma
Molecular Cancer (2015)
-
Regulation of tight junctions by sex hormones in normal human endometrial epithelial cells and uterus cancer cell line Sawano
Cell and Tissue Research (2013)
-
Pathways involved in Drosophila and human cancer development: the Notch, Hedgehog, Wingless, Runt, and Trithorax pathway
Annals of Hematology (2012)
