
Abstract
The ability of pneumococcal conjugate vaccine (PCV) to decrease transmission by blocking the acquisition of colonization has been attributed to herd immunity. We describe the role of mucosal immunoglobulin G (IgG) to capsular polysaccharide (CPS) in mediating protection from carriage, translating our findings from a murine model to humans. We used a flow cytometric assay to quantify antibody-mediated agglutination demonstrating that hyperimmune sera generated against an unencapsulated mutant was poorly agglutinating. Passive immunization with this antiserum was ineffective to block acquisition of colonization compared to agglutinating antisera raised against the encapsulated parent strain. In the human challenge model, samples were collected from PCV and control-vaccinated adults. In PCV-vaccinated subjects, IgG levels to CPS were increased in serum and nasal wash (NW). IgG to the inoculated strain CPS dropped in NW samples after inoculation suggesting its sequestration by colonizing pneumococci. In post-vaccination NW samples pneumococci were heavily agglutinated compared with pre-vaccination samples in subjects protected against carriage. Our results indicate that pneumococcal agglutination mediated by CPS-specific antibodies is a key mechanism of protection against acquisition of carriage. Capsule may be the only vaccine target that can elicit strong agglutinating antibody responses, leading to protection against carriage acquisition and generation of herd immunity.
Similar content being viewed by others


Introduction
The human nasal mucosa forms the first line of defence against respiratory pathogens. Some of these pathogens such as Streptococcus pneumoniae (the pneumococcus) can asymptomatically colonize the upper respiratory tract (the carrier state).1 Although most episodes of pneumococcal carriage do not result in disease, the organism may gain access to normally sterile sites in its human host from its niche on the mucosal surfaces of the upper airways.2 Mucosal immune responses, therefore, have a critical role in the defence against pneumococcal infections as they dictate the outcome of host–pathogen interactions at the mucosa.
Murine models have demonstrated that once carriage is established the generation of mucosal antibodies is ineffective at clearing the organism.3, 4 However, mucosal antibody, if present before stable colonization occurs, may block acquisition through its agglutinating activity, a mechanism dependent on its multi-valency and independent of Fc, complement and opsonophagocytosis.5 The ability of agglutinating antibody to inhibit the establishment of mucosal colonization could be attributed to more efficient mucociliary clearance of larger particles and the requirement for a larger colonizing dose. As pneumococci enzymatically inactivate the agglutinating activity of human IgA1, the most abundant form of immunoglobulin on the airway surface, the prevention of colonization requires sufficient mucosal levels of other subclasses such as immunoglobulin G (IgG).6 The ability of the pneumococcus to target and evade human-specific components of humoral immunity emphasizes the need to examine the mechanisms of mucosal protection in the natural host.
The serotype-specific success of the pneumococcal conjugate vaccine (PCV) in reducing rates of carriage of vaccine-type strains in immunized populations indicates that anti-capsular antibodies reduce transmission by blocking the acquisition of colonization.7 PCV vaccination induces high levels of serum IgG that access the mucosal surface in vaccinated children, however, the exact mechanism by which this vaccine mediates mucosal protection has not been described.8 We recently reported that PCV conferred a 78% reduction in carriage acquisition compared to a control group following inoculation of adults with live type 6B pneumococci in an experimental human pneumococcal carriage (EHPC) study.9
In this report, we utilize a flow cytometric assay to quantify the agglutinating effect of anti-pneumococcal antibodies. This assay allowed us to examine the role of pneumococcal surface antigens and demonstrate the importance of antibodies to its immunodominant antigen, capsular polysaccharide (CPS), in eliciting agglutinating IgG that protects from the acquisition of colonization. This assay was then used to investigate the role of mucosal antibodies to capsule antigens in mediating agglutination and protection against acquisition of pneumococcal carriage in the natural host in an EHPC study of PCV.
Results
A flow cytometric assay to quantify pneumococcal agglutination by antibody
To quantify bacterial agglutination, we developed and optimized a flow cytometric assay. After a brief incubation of pneumococci with type-specific antibody, there was a dose-dependent increase in the shift in forward scatter (FSC) (Figure 1a). At higher concentrations, there was also an increase in side scatter. Samples were then analyzed under similar conditions using an Amnis Imaging Flow Cytometer to visualize the individual events detected by the laser. The change in particle size, as detected by shift in FSC, and complexity, as detected by shift in side scatter, correlated with a progressive bridging of particles to form longer chains (threading reaction) by antibody.10 As the concentration of antibody was increased further these formed into aggregates of increasing size. Furthermore, divalent F(ab’)2 fragments generated from this antibody5 caused a similar shift in FSC and corresponding visual agglutination of bacteria, unlike an equivalent concentration of monovalent Fab fragments (Figure 1b). Together these data confirm that flow cytometry is a sensitive method to detect and quantify bacterial agglutination by antibody.


Flow cytometric assay to quantify agglutination. (a) Representative dot plots from BD FACS Calibur with FSC v SSC of P1121 with increasing concentrations of type-specific rabbit anti-pneumococcal serum (corresponding to a total IgG concentration of 0, ∼2.5, ∼25, and ∼250 μg ml–1). Bacterial cells were gated to remove small debris particles. Percent agglutination is calculated by the sum of events in Q1, Q2, and Q3. Representative images are shown from Amnis ImageStream Imaging flow cytometer corresponding to the conditions shown above. (b) Representative dot plots and images from P1121 incubated with either Fab or F(ab’)2 fragments of type-specific rabbit anti-pneumococcal IgG at 50 μg ml–1.
Capsule is the major agglutinating antigen and leads to enhanced protection
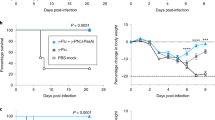
Using this flow cytometric assay for agglutination, we examined which pneumococcal surface antigens could generate agglutinating antibodies. We raised hyperimmune rabbit antisera against whole-cell heat-killed P1121, a type 23F isolate (P2109, encapsulated strain) and a genetically modified mutant of P1121 lacking the capsular antigen (unencapsulated strain). Immunization with these strains led to a robust antibody response, and the enzyme-linked immunosorbent assay (ELISA) IgG titers to whole bacteria were not significantly different whether antiserum was raised against the encapsulated or unencapsulated strain (Figure 2a). As expected, only antiserum raised to the encapsulated strain contained antibody recognizing purified type 23 CPS by ELISA (Figure 2b). Using these rabbit antisera, we then compared agglutination using the flow cytometric assay and found that only antiserum raised against the encapsulated strain was able to agglutinate the wild-type encapsulated bacteria (Figure 2c). In contrast, the antiserum raised against the isogenic strain lacking the capsular antigen showed minimal agglutination of either the encapsulated or the unencapsulated strains. This suggests that although the array of non-capsular antigens underlying the capsule is able to induce a strong immune response, antibody to these antigens is relatively poorly agglutinating. Therefore, capsular antigens may be the only antigens that can efficiently promote agglutination. To confirm the requirement for the capsular antigen in agglutination, isogenic strains in which the capsule-type was genetically switched were compared using the flow cytometric assay. Pneumococcal agglutination required type-specific antibody regardless of genetic background (Figure 2d). There did not appear to be a significant contribution of other constituents determined by genetic background to agglutination. The agglutinating ability of antibodies has been shown to protect from the acquisition of colonization.5 We used a murine model of colonization to compare the antisera raised to the isogenic encapsulated and unencapsulated strains. After intranasal challenge with strain P1121, mice that had been passively immunized with antisera against the encapsulated strain were more protected from colonization compared with mice immunized with the antisera against its unencapsulated mutant (Figure 2e). This confirmed that the agglutinating ability of anti-capsular antibody is important for limiting colonization acquisition.

Capsule is the major agglutinating antigen and leads to enhanced protection. (a) ELISA geometric mean titer (GMT) of rabbit antisera raised to isogenic capsular polysaccharide (cps) ± strains, binding to encapsulated (P1121) or unencapsulated (P1121Δcps) whole-cell pneumococci. Mean±s.d., n=2–4 per condition. (b) ELISA GMT of rabbit sera raised to cps±strains binding to type 23 purified CPS. Mean±s.d., n=3. (c) Flow cytometric agglutination assay comparing the titers at which a threefold increase in percent agglutination of pneumococci is observed with rabbit antisera raised to cps±strains. Mean±s.d., n=3–4 per condition. (d) Flow cytometry agglutination assay comparing the titers at which a threefold increase in percent agglutination for strains in which the capsule genes and type were switched is observed with rabbit antisera raised to encapsulated P1121 (type 23F). Mean±s.d., n=4–5 per condition. The baseline percent agglutination was calculated for each individual experiment and ranged from only 2–10%. (e) Passive protection experiment in mice immunized IP with rabbit serum raised to cps±strains. Four hours later, mice were given an intranasal dose of P1121 and colonization measured at 20 h post inoculation. Mean±s.e.m. Kruskal–Wallis test with Dunn's post-test was performed for (a), (c), and (d). Unpaired t-test was performed for (b), and Mann–Whitney test was performed for (E). ns, non-significant, **P<0.01, and ****P<0.001.
Anti-PS IgG-mediated protection against experimental carriage in PCV-vaccinated subjects
To investigate whether anti-capsular antibody generated through vaccination with PCV could mediate protection against carriage acquisition through agglutination, we studied samples from the PCV/EHPC study in which PCV-vaccinated subjects had 78% protection against carriage acquisition with 6B strain.9 IgG levels to the vaccine-included CPS 6B and 23F were measured at Pre-V, Post-V and 21 days after pneumococcal inoculation in sera samples as well as at day 2 after inoculation in nasal wash (NW) samples (Supplementary Figure S1 online).
As expected, PCV-vaccinated subjects had increased IgG levels to both capsule-types in serum Post-V compared with Pre-V levels (Figure 3). Only 5 out of 48 subjects vaccinated with PCV became colonized following pneumococcal inoculation (carriage+). In carriage+ subjects, only a modest 2.6-fold increase in levels of IgG to 6B CPS was observed in serum following vaccination, considerably less than the 8.4-fold increase observed in subjects protected from carriage (carriage−) (Figure 3a). A similar pattern was observed in NW samples (Figure 3c). Interestingly, amongst carriage+ subjects, only one volunteer had increased IgG levels in NW Post-V. Pneumococcus was recovered from the nasopharynx of this volunteer at a very low density and at only one time point.

Drop in type-specific IgG levels to CPS in nasal wash following pneumococcal inoculation. (a, c, e, g) IgG levels to 6B CPS (b, d, f, h) and IgG levels to 23F CPS. Levels were measured in serum and nasal wash samples in (a–d) PCV-vaccinated subjects and (e–h) control group using WHO standardised ELISA. Each dot represents IgG levels from a subject expressed in ng ml–1. Levels were measured from carriage− (closed dots) and carriage+ (open dots) subjects at the time points indicated on the X axis. Data are presented as GMC and 95% CI. Results were analyzed using one-way ANOVA test and Bonferroni’s multiple comparison tests. ***P<0.0001, **P<0.005, *P<0.01.
Although IgG levels remained unaltered following pneumococcal inoculation in sera (Figures 3a and b), levels of IgG to the CPS of inoculated 6B strain significantly dropped in NW 2 days after inoculation compared with levels prior to inoculation (Figure 3c). No decrease was observed in levels of IgG to 23F CPS following pneumococcal inoculation with the 6B strain (Figure 3d). These data suggest that capsule-specific antibody is sequestered in the nasal lumen following pneumococcal inoculation that could have a role in protection against carriage acquisition.
Carriage boosts pre-existing levels of IgG to CPS of inoculated strain
We have previously shown that exposure to pneumococci following intranasal inoculation boosted pre-existing levels of CPS IgG to 6B CPS in serum only for subjects who developed carriage.11 In this study, we confirmed this observation in a larger group of participants and further observed that levels are also increased in NW following carriage (Figures 3e and g). As expected, we observed no increase in levels of IgG to both capsular-types in the control group following vaccination with the Hep-A vaccine (Figures 3e–h) and no alteration in levels of IgG to the 23F CPS following inoculation with the 6B strain (Figures 3f and h).
Correlation between levels of IgG to 6B CPS in serum and NW samples
There was a positive correlation between levels of IgG to 6B CPS in serum and NW in PCV-vaccinated subjects (Figure 4), which indicates that systemic antibodies elicited by vaccination transudate to the nasal lumen. A similar correlation was observed between levels of systemic and nasal IgG in carriage+ subjects 21 days following carriage (Supplementary Figure S2).

Sera IgG levels correlate with mucosal IgG following PCV vaccination. Correlation between IgG levels to 6B CPS measured in sera and nasal wash samples collected post PCV vaccination (Post-V) in carriage− (closed dots) and carriage+(open dots) subjects. Spearman r=0.59 and P<0.0001.
IgG mediates bacterial agglutination in the nasal mucosa in PCV-vaccinated subjects
We then examined whether IgG present in NW from PCV-vaccinated subjects could promote bacterial agglutination. Using concentrated NW samples incubated with the 6B strain, we observed increased pneumococcal agglutination in Post-V samples compared with Pre-V samples (19.4±15.1 vs 12.4±7.8). This increased antibody-mediated agglutination was observed only in volunteers protected from carriage acquisition (carriage−) (Figure 5a). Levels of IgG to 6B CPS correlated with levels of agglutination observed in NW from carriage− (Figure 5b; closed circles) but not carriage+ subjects (Figure 5b; open circles).

Increased nasal antibody-mediated pneumococcal agglutination promoted by PCV vaccination. (a) Percentage of pneumococcal agglutination promoted by nasal wash samples from PCV-vaccinated subjects before inoculation, carriage− (closed dots, n=20) and carriage+ subjects (open dots, n=5) before (Pre-V) and after (Post-V) vaccination. Data are presented as GMC and 95% CI. Results were analyzed using Mann–Whitney test. (b) Correlation between IgG levels to CPS 6B Post-V in nasal washes and promoted pneumococcal agglutination % in carriage− (closed dots) and carriage+ (open dots) subjects. Spearman r=0.60 and P=0.002.
We also examined agglutination capacity of IgG present in NW from Hep-A-vaccinated control subjects prior to pneumococcal inoculation, to investigate whether agglutination capacity of naturally acquired IgG was associated with protection against carriage acquisition. No difference was observed between carriage− and carriage+ groups (Figure 6a) and agglutination levels were low in both groups, which could be due to the low levels of IgG to 6B CPS present in these samples (Figure 3g). No correlation was observed between IgG to 6B CPS present in NW and agglutination levels in either carriage− or carriage+ groups (Figures 6b and c).

Nasal wash agglutination capacity does not correlate with protection from experimental carriage in control-vaccinated subjects. (a) Percentage of pneumococcal agglutination promoted by nasal wash samples obtained from control group (Hep-A vaccinated) before pneumococcal inoculation, carriage− (closed dots, n=20) and carriage+ subjects (open dots). Data are presented as GMC and 95% CI. Results were analyzed using Mann–Whitney test. (b) Correlation between IgG levels to CPS 6B in nasal washes and promoted pneumococcal agglutination % in carriage− subjects. Spearman r=0.06 and P=0.79. (c) Correlation between IgG levels to CPS 6B in nasal washes and promoted pneumococcal agglutination % in carriage+ subjects. Spearman r=0.13 and P=0.56.
Discussion
Our study provides insight into the mechanisms of mucosal defence against pathogens and how humoral immunity generated through vaccination contributes to protection. We demonstrate that the ability of antibody to block the establishment of colonization in the human host, the first step in pathogenesis of disease caused by S. pneumoniae, correlates with its agglutinating activity.1, 12 Our focus was on IgG because it is generated in high concentrations in response to systemic immunization and has been shown to be sufficient to promote agglutination on the mucosal surface.5 We have measured both IgA1 and IgA2 in NW samples pre- and post-inoculation with pneumococcus and IgA1 was the dominant IgA subclass in the nasal mucosa (data not shown). Secretory antibodies are unlikely to be sufficient factor in agglutination owing to the activity of pneumococcal IgA1 protease and the moderated increase of S-IgM levels post vaccination.13, 14
This study required a sensitive method to quantify agglutination. Through use of technology that simultaneously provides images of individual events detected during flow cytometric analysis, we confirmed that flow characteristics were a sensitive and specific measure of the magnitude of antibody-induced agglutination. By comparing hyperimmune sera generated to isogenic strains differing only in expression of CPS amount and type, we showed that type-specific antibody to CPS was necessary for agglutination. Data from the EHPC study with parenterally administered PCV confirmed that anti-CPS IgG is protective from colonization and is sufficient to generate mucosal agglutinating activity. This observation provided mechanistic understanding of the effectiveness of CPS-based immunity in reducing rates of mucosal infection and conferring herd immunity in the population.15 This same mechanism may be applicable to vaccines using the CPSs of other encapsulated pathogens that also impact mucosal colonization.16, 17 In our study using whole pneumococci, only antibody to CPS was agglutinating. Yet, it remains possible that a sufficient amount of antibody to another pneumococcal target or combination of targets could elicit agglutinating antibody. It also remains possible, however, that CPS is the only pneumococcal target that may elicit agglutination by specific antibodies. CPS is a highly abundant surface antigen and capable of inducing high levels of immunoglobulin, particularly when conjugated to an immunogenic protein carrier. In addition, CPS variably shields underlying surface antigens from recognition by antibody. However, even when equivalent amounts of antibody to whole pneumococci were compared, only sera containing antibody to type-specific CPS elicited detectable agglutination. Thus, the amount of bound antibody does not appear to be the only factor contributing to agglutination.
Using an experimental model of human pneumococcal carriage, we previously demonstrated that PCV is highly protective against 6B pneumococcal carriage acquisition, conferring a 78% reduction in carriage acquisition.9 We now document that in protected adults, PCV induced high levels of IgG to 6B CPS at the nasal mucosa. NWs collected post PCV vaccination had increased pneumococcal agglutination capacity compared with NW samples collected before vaccination. Agglutination levels correlated with levels of IgG to 6B CPS. Interestingly, although levels of IgG to 23F CPS were unaltered in NW after inoculation, levels of IgG to 6B CPS were reduced—this suggests that antibodies to 6B CPS were sequestered onto the bacterial surface in the nasal lumen following inoculation. Thus, the immunogenicity of PCV is sufficient to generate levels of IgG that reach the mucosal surface in amounts that bind to and agglutinate pneumococci when the host is exposed to the pathogen. Optimal protection may require the presence of agglutinating levels of antibody at the time of first exposure on the mucosal surface potentially explaining why the vaccine may prevent new carriage events but does not impact pre-existing colonization. It remains unknown whether humoral immunity generated by natural carriage generates protective agglutinating antibody that affects subsequent type-specific exposure to the organism. In previous EHPC studies we have shown that pneumococcal carriage protects healthy adults against subsequent carriage following re-exposure to the homologous strain11 but not against acquisition of a heterologous strain type.18 More recently we have shown that high baseline levels of circulating memory B-cells secreting IgG to CPS, but not to protein antigens, were associated with protection against carriage acquisition in unvaccinated adults.18 Taken together, our findings suggest that anti-capsular IgG-mediated agglutination is a key mechanism of protection against pneumococcal carriage acquisition, in particular in subjects with high antibody titers, such as those vaccinated with PCV.
Our findings have several implications for vaccine development. Current anti-pneumococcal vaccines are limited by a serotype-specific approach and the diversity of pneumococcal types. Most of the effectiveness of pneumococcal vaccines, however, results from their ability to reduce transmission by decreasing rates of colonization in the population.15 Thus, for any novel vaccine containing more conserved target(s) to be as effective it would need to similarly impact carriage, and our study suggests that this would require the generation of agglutinating antibody on the mucosal surface of the upper airways.
Materials and Methods
Pneumococcal strains The strains used in this study include BHN418 (type 6B, used for human experimental carriage studies),19 P1121 (type 23F, isolated from the nasopharynx in a human experimental carriage study),20 P2109 (P1121Δcps)20 intermediate mutant in creating 2140), P1542 (type 4 isolate),21, 22 P1690 (P1542 background with type 23 F capsule),23 P2140 (P1121 background with type 4 capsule).23 A GFP-expressing version of P1121 was used to differentiate bacteria from calibration beads in experiments with the Amnis ImageStream Imaging Flow Cytometer. All strains were grown in tryptic soy broth at 37 °C, and were passaged intranasally in mice prior to preparation of frozen stocks, with exception of the 6B strain which were grown in Vegetone broth (for human inoculation) at 37 °C 5% CO2 until early log phase and frozen.
PCV/EHPC study The PCV/EHPC study was conducted in 2012 and detailed methods for recruitment, nasopharyngeal pneumococcal inoculation and carriage detection as well as study design and study outcomes have been previously described.9 Ethical approval was obtained from the National Health Service Research Ethics Committee (REC) (12/NW/0873). This study was co-sponsored by the Royal Liverpool and Broadgreen University Hospitals Trust and the Liverpool School of Tropical Medicine.
In brief, 96 healthy adults aged 18–50 years were enrolled with informed consent and randomised to receive a single dose of either PCV-13 (Prevnar, Pfizer) or Hep-A vaccine as a control group (Avaxim, Sanofi Pasteur MSD). After 4–5 weeks following vaccination, subjects were intranasally inoculated with live 6B pneumococcus (BHN418) (80,000 CFUs (colony-forming units) per nostril).24 Sera samples were collected before vaccination (Pre-V), after vaccination/prior to pneumococcal inoculation (Post-V) and 21 days after pneumococcal inoculation. NW samples were collected at the same time points and also at days 2, 7, and 14 following pneumococcal inoculation (Supplementary Figure S1).
Antibody level measurements IgG levels to CPCs 6B and 23F were measured using the WHO standardised ELISA method, as previously described.7, 11, 25 Whole-cell ELISA was performed, as previously described21 with the following minor modifications; serum was diluted in doubling concentrations, and the standardized development time altered to 30 min. Antigen-specific antibodies were detected by goat anti-mouse IgG (1/4,000; heavy and light chains)-alkaline phosphatase (Sigma, St. Louis, MO). Purified type 23 CPS (ATCC) was fixed to Immulon 1B plates at a final concentration of 5 μg ml–1 in saline at 4 °C and used in an ELISA to quantify anti-capsular antibody in rabbit serum, as described previously.5
Hep-A specific IgG purification from human serum samples Hep-A IgG was purified from seven pooled sera samples taken from Hep-A-vaccinated subjects to be used as a negative control in agglutination assays. Protein specific IgG purification was done in two-stages following the manufacturer’s instructions and as previously published by our group.26 First, total IgG was purifed by Sepharose protein G (GE Healthcare, Little Chalfont, UK) and then anti-Hep-A IgG by CNBr activated Sepharose (300 mg CNBr activated Sepharose gel coupled with 1 mg Hep-A purified protein, Abcam (Cambridge, UK) ab49011). Dot blot and ELISA were performed to confirm anti-Hep-A IgG purification and concentration (data not shown).
Rabbit antisera Fixed and heat-killed whole-cell bacteria were prepared as follows; P1121 and P2109 were grown at 37 °C to mid-log-phase, fixed in 1% paraformaldehyde for 1 h at room temperature, washed in phosphate-buffered saline, incubated for 30 min at 65 °C, and stored at 4 °C.27 Antisera were generated in rabbits by Cocalico Biological (Reamstown, PA, USA). In total, 50 μg bacterial protein (∼2 × 108 CFU) was injected intravenously three times weekly for 16 weeks the point at which titers no longer increased, as previously described.28 Fab and F(ab’)2 fragments were generated from type-specific rabbit IgG as previous described.5
Agglutination assay Pneumococci were grown to mid-log-phase and stored at −80 °C in glycerol. On the day of the experiment, cells were thawed and washed with phosphate-buffered saline. For agglutination assays with human NW samples 3 μl of 105 bacteria was incubated with 47 μl of concentrated NW supernatant (1 ml of NW concentrated to 100 μl using vacuum concentrator RVC2-18) and samples were vortexed lightly. Antiserum to group 6 (Statens Serum Institute, Neufeld antisera to group 6) was used as a positive control and Anti-Hep-A purified human IgG was used as a negative control. Samples were incubated for 1.5 h at 37 °C, fixed with paraformaldehyde and analyzed on a BD LSR II Flow Cytometer (BD Biosciences, San Jose, CA, USA). Bacterial population was gated in the FSC and Sideward scatter dot plot referring to cell size and granularity. PMT voltages and threshold were gated on negative control bacteria. Agglutination was quantified by calculating the proportion of the bacterial population with altered FSC and side scatter and values were expressed as % of agglutination, as previously described.29, 30 All samples were analyzed in duplicate and 30,000 events were acquired using FacsDiva Software 6.1 (BD Biosciences). Analysis was performed using FlowJo software version 10.0 (Tree Star, San Carlos, CA, USA).
Similar assays were performed for rabbit sera with minor modifications; several serum dilutions were tested, samples were incubated for 1 h at 37 °C then analyzed on a BD FACSCalibur Flow Cytometer. Agglutination was quantified as above for each serum dilution. Agglutination was confirmed by imaging events at × 60 magnification on an Amnis ImageStream Imaging Flow Cytometer equipped with INSPIRE software (Amnis, EMD Millipore). Image analysis was performed using IDEAS software (Amnis, EMD Millipore, Darmstadt, Germany).
Passive immunization experiments Passive immunization was performed, as described previously.5 C57Bl/6 J (Jackson Laboratories, Bar Harbor, ME, USA) mice were housed in accordance with Institutional Animal Care and Use Committee protocols. In total, 5–6 weeks old adult mice were immunized intraperitoneally with 200 μl of hyperimmune rabbit anti-pneumococcal sera. Mice were inoculated intranasally 4 h post immunization with 10 μl containing ∼2 × 104 CFU P1121 in phosphate-buffered saline. At 20 h post pneumococcal inoculation, mice were euthanized, the trachea cannulated, and 200 μl of phosphate-buffered saline instilled and lavage fluid was collected from the nares for quantitative culture. Lavage fluid was vortexed vigorously prior to plating to ensure any bacterial aggregates were dissociated. We have previously used quantification by a DNA-based assay to confirm that agglutination was not confounding colony counting.5 The limit of detection was 2 CFU per animal.
Statistics Statistical analyses were performed using GraphPad Prism 5 (GraphPad Software, Inc, La Jolla, CA, USA). For comparison of murine data Mann–Whitney test was performed when two groups were compared and Kruskal–Wallis test with Dunn's post-test was performed when three or more groups were compared. Where appropriate, data were logarithmically transformed to obtain data with a normal distribution. Unpaired t-tests were used to compare levels of purified CPS between groups. Multiple comparisons were made within carriage+ and carriage− groups using one-way analysis of variance with Bonferroni post-test. Sera IgG levels correlated with mucosal IgG using Spearman’s correlation, linear regression. Differences were considered significant at P ⩽0.05.
References
Bogaert, D., De, G.R. & Hermans, P.W. Streptococcus pneumoniae colonisation: the key to pneumococcal disease. Lancet Infect. Dis. 4, 144 (2004).
Charlson, E.S. et al. Topographical continuity of bacterial populations in the healthy human respiratory tract. Am. J. Respir. Crit. Care Med. 184, 957–963 (2011).
McCool, T.L. & Weiser, J.N. Limited role of antibody in clearance of Streptococcus pneumoniae in a murine model of colonization. Infect. Immun. 72, 5807 (2004).
Malley, R., Trzcinski, K., Srivastava, A., Thompson, C.M., Anderson, P.W. & Lipsitch, M. CD4+ T cells mediate antibody-independent acquired immunity to pneumococcal colonization. Proc. Natl. Acad. Sci. USA 102, 4848 (2005).
Roche, A.M., Richard, A.L., Rahkola, J.T., Janoff, E.N. & Weiser, J.N. Antibody blocks acquisition of bacterial colonization through agglutination. Mucosal Immunol. 8, 176–185 (2015).
Janoff, E.N. et al. Pneumococcal IgA1 protease subverts specific protection by human IgA1. Mucosal Immunol. 7, 249–256 (2014).
Goldblatt, D. et al. Establishment of a new human pneumococcal standard reference serum, 007sp. Clin. Vaccine Immunol. 18, 1728–1736 (2011).
Nurkka, A., Ahman, H., Korkeila, M., Jantti, V., Kayhty, H. & Eskola, J. Serum and salivary anti-capsular antibodies in infants and children immunized with the heptavalent pneumococcal conjugate vaccine. Pediatr. Infect. Dis. J. 20, 25–33 (2001).
Collins, A.M. et al. First human challenge testing of a pneumococcal vaccine - double blind randomised controlled trial. Am. J. Respir. Crit. Care Med. 192, 853–858 (2015).
Dalia, A.B. & Weiser, J.N. Minimization of bacterial size allows for complement evasion and is overcome by the agglutinating effect of antibody. Cell Host Microbe 10, 486–496 (2011).
Ferreira, D.M. et al. Controlled human infection and rechallenge with Streptococcus pneumoniae reveals the protective efficacy of carriage in healthy adults. Am. J. Respir Crit. Care. Med. 187, 855–864 (2013).
Simell, B. et al. The fundamental link between pneumococcal carriage and disease. Expert Rev. Vaccines 11, 841–855 (2012).
Lue, C., Tarkowski, A. & Mestecky, J. Systemic immunization with pneumococcal polysaccharide vaccine induces a predominant IgA2 response of peripheral blood lymphocytes and increases of both serum and secretory anti-pneumococcal antibodies. J. Immunol. 140, 3793–3800 (1988).
Petrunov, B. et al. Cellular and humoral systemic and mucosal immune responses stimulated in volunteers by an oral polybacterial immunomodulator "Dentavax". Int. Immunopharmacol. 6, 1181–1193 (2006).
Klugman, K.P. Herd protection induced by pneumococcal conjugate vaccine. Lancet Glob. Health 2, e365–e366 (2014).
Fernandez, J. et al. Prevention of Haemophilus influenzae type b colonization by vaccination: correlation with serum anti-capsular IgG concentration. J. Infect. Dis. 182, 1553–1556 (2000).
Bijlsma, M.W., Brouwer, M.C., Spanjaard, L., van de Beek, D. & van der Ende, A. A decade of herd protection after introduction of meningococcal serogroup C conjugate vaccination. Clin. Infect. Dis. 59, 1216–1221 (2014).
Pennington, S.H. et al. Polysaccharide-specific memory B-cells predict protection against experimental human pneumococcal carriage. Am. J. Respir. Care Med. e-pub ahead of print 12 July 2016.
Browall, S. et al. Intraclonal variations among Streptococcus pneumoniae isolates influence the likelihood of invasive disease in children. J. Infect. Dis. 209, 377–388 (2014).
McCool, T.L., Cate, T.R., Moy, G. & Weiser, J.N. The immune response to pneumococcal proteins during experimental human carriage. J. Exp. Med. 195, 359 (2002).
Roche, A.M., King, S.J. & Weiser, J.N. Live attenuated Streptococcus pneumoniae strains induce serotype-independent mucosal and systemic protection in mice. Infect. Immun. 75, 2469–2475 (2007).
Tettelin, H. et al. Complete genome sequence of a virulent isolate of Streptococcus pneumoniae. Science 293, 498–506 (2001).
Lysenko, E.S., Lijek, R.S., Brown, S.P. & Weiser, J.N. Within-host competition drives selection for the capsule virulence determinant of Streptococcus pneumoniae. Curr. Biol. 20, 1222–1226 (2010).
Gritzfeld, J.F. et al. Experimental human pneumococcal carriage. J. Vis. Exp. 72 (2013).
Wright, A.K. et al. Human nasal challenge with streptococcus pneumoniae is immunising in the absence of carriage. PLoS Pathog. 8, e1002622 (2012).
Glennie, S. et al. Modulation of nasopharyngeal innate defenses by viral coinfection predisposes individuals to experimental pneumococcal carriage. Mucosal Immunol. 9, 56–67 (2016).
Austrian, R. & Bernheimer, H.P. Simultaneous production of two capsular polysaccharides by pneumococcus. I. Properties of a pneumococcus manifesting binary capsulation. J. Exp. Med. 110, 571–584 (1959).
Schiffman, G., Bornstein, D.L. & Austrian, R. Capsulation of pneumococcus with soluble cell wall-like polysaccharide. II. Nonidentity of cell wall and soluble cell wall-like polysaccharides derived from the same and from different pneumococcal strains. J. Exp. Med. 134, 600–617 (1971).
Habets, M.N., van Salm, S., van der Gaast- de Jongh, C.A., Diavatopoulos, D.A. & de Jonge, M.I. A novel flow cytometry-based assay for the quantification of antibody-dependent pneumococcal agglutination. PLoS One.
Yitzhaki, S., Barnea, A., Keysary, A. & Zahavy, E. New approach for serological testing for leptospirosis by using detection of leptospira agglutination by flow cytometry light scatter analysis. J. Clin. Microbiol. 42, 1680–1685 (2004).
Acknowledgements
We gratefully would like to thank all subjects who participated in this study as well as all staff of the Clinical Research Unit at the Royal Liverpool Hospital and the clinical staff of the Respiratory Infection Group at the Liverpool School of Tropical Medicine. We also thank Dr Michael Betts, Jay Gardner, and Dr Morgan Reuter-Moslow for guidance with the Amnis Image Stream. This work was funded by the Bill and Melinda Gates Foundation (OPP1117728), the Medical Research Council and FAPESP (MR/K01188X/1) grants awarded to DMF and SBG, and NIH grants (AI038446 and AI105168) to JNW.
Author contribution
Conception and design: EM, AMR, JR, SHP, MIJ, SBJ, JNW, DMF. Analysis and interpretation: EM, AMR, JR, TZ, JTO, SHP, JFG, ADW, AMC, SS, MIJ, SBG, JNW, DMF. Drafting the manuscript for important intellectual content: EM, AMR, JR, SHP, JNW, DMF. The authors acknowledge support from Wellcome Trust Multi-User Equipment Grant (104936/Z/14/Z).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
JNW receives royalties from GlaxoSmithKline for a pneumococcal vaccine. The remaining authors declare no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Mitsi, E., Roche, A., Reiné, J. et al. Agglutination by anti-capsular polysaccharide antibody is associated with protection against experimental human pneumococcal carriage. Mucosal Immunol 10, 385–394 (2017). https://doi.org/10.1038/mi.2016.71
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2016.71
This article is cited by
-
Pneumococcal within-host diversity during colonization, transmission and treatment
Nature Microbiology (2022)
-
Pneumococcal capsule blocks protection by immunization with conserved surface proteins
npj Vaccines (2021)
-
Microinvasion by Streptococcus pneumoniae induces epithelial innate immunity during colonisation at the human mucosal surface
Nature Communications (2019)
-
Streptococcus pneumoniae: transmission, colonization and invasion
Nature Reviews Microbiology (2018)
