
Abstract
Proteorhodopsin (PR) is a wide-spread protein found in many marine prokaryotes. PR allows for the potential conversion of solar energy to ATP, possibly assisting in cellular growth and survival during periods of high environmental stress. PR utilises either blue or green light through a single amino acid substitution. We incubated the PR-bearing bacterium Psychroflexus torquis 50 cm deep within Antarctic sea ice for 13 days, exposing cultures to diurnal fluctuations in light and temperature. Enhanced growth occurred most prominently in cultures incubated under irradiance levels of ∼50 μmol photons m−2 s−1, suggesting PR provides a strong selective advantage. In addition, cultures grown under blue light yielded over 5.5 times more live cells per photon compared to green-light incubations. Because P. torquis expresses an apparently ‘green-shifted’ PR gene variant, this finding infers that the spectral tuning of PR is more complex than previously thought. This study supports the theory that PR provides additional energy to bacteria under sub-optimal conditions, and raises several points of interest to be addressed by future research.
Similar content being viewed by others


Introduction
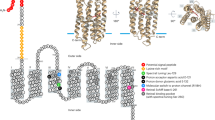
The protein proteorhodopsin (PR) utilises solar irradiance to translocate protons across cell membranes of many prokaryotes (Béjà et al., 2000, 2001; Finkel et al., 2013), which allows for ATP synthesis. Consequently, the growth-rate under visible light increases for some PR-containing bacteria (Gómez-Consarnau et al., 2007; Palovaara et al., 2014; Gomez-Consarnau et al., 2016), but, surprisingly, not for others (Giovannoni et al., 2005; Lami et al., 2009; Steindler et al., 2011). This variation in response led Fuhrman et al. (2008) to suggest that PR may be utilised to buffer environmental stress. Antarctic sea ice harbours wide gradients of temperature (Petrich and Eicken, 2009), salinity (Kottmeier and Sullivan, 1988) and (predominantly blue) light (Petrich and Eicken, 2009; Ryan et al., 2009). When cultured under salinity stresses similar to those in sea-ice brine channels (Arrigo et al., 1997; Petrich and Eicken, 2009), the growth of the Antarctic sea-ice PR-bacterium Psychroflexus torquis (Bowman et al., 1998) was stimulated by light (Feng et al., 2013).
PR can be spectrally tuned through the substitution of a single amino acid at PR-position 105 (Béjà et al., 2001; Man et al., 2003). A glutamine ensures maximum absorption at 490 nm, whereas other amino acids shift it to 530 nm (Béjà et al., 2001; Man et al., 2003; Riedel et al., 2010). In sea ice, blue light in upper sections typically favour blue-shifted PR-bacteria (Koh et al., 2010), while green-shifted species populate lower sections, where high chlorophyll concentrations ensure the only available light is green (Koh et al., 2010). To assess how in situ conditions affect the PR-response of P. torquis, we incubated monocultures within Antarctic sea ice, exposing them to various wavebands and intensities of visible light under natural diurnal variations in temperature and solar irradiance. We expected to observe an increase in growth under illumination, and as P. torquis possesses a methionine at position 105 (Bowman et al., 1998), we expected the highest light-induced growth under green light.
Materials and methods
P. torquis ATCC 700755T was cultured within annual sea ice at Cape Evans, Antarctica (77°38′S, 166°24′E). A 100 μl aliquot of stock culture was inoculated into 10 ml of sterile Marine Broth 2216 (Becton Dickinson, Auckland, New Zealand) in 15 ml tubes wrapped with polycarbonate filters (Wellington Photographic Supplies, Wellington, New Zealand). Treatments were: ambient photosynthetically active radiation (PAR; no filter), reduced PAR (50% Neutral Density), blue (141-Bright Blue), green (122-Fern Green), red (021-Gold Amber) or darkness (aluminium foil). A series of 5 cm-diameter holes were drilled in 1.7 m-thick annual ice (Kovacs, Roseburg, OR, USA). Each sample was randomly allocated to a single hole and incubated at 50 cm depth within the sea ice for up to 13 days (N=5 per treatment). A one-way analysis of variance (ANOVA; α=0.05) was used to compare each treatment and Tukeyb post hoc tests differentiated subgroups within the population. Subgroups were compared again using a secondary series of one-way analysis of variances, thus avoiding the use of harmonic mean group sizes (Smith, 1971).
Results and discussion
The mean surface PAR at midday was ∼1150 μmol photons m−2 s−1, although irradiances fell below this value over daily cycles. The estimated mean midday irradiance (μmol photons m−2 s−1) at 50 cm was 103.5 (ambient), 48.6 (reduced), 45.6 (blue), 54.4 (green), 9.5 (red) and 0 (dark), taking account of the differential attenuation of these wavelengths in ice, and the filter transmission properties (Table 1). Thus, reduced PAR, blue and green treatments provided approximately equal photon fluxes at 50 cm depth (analysis of variance F2,39=1.55, P=0.22), at about half that of the ambient treatment (F1,54=96.58, P<0.001), and over five times greater than the red treatment (F1,54=124.32, P<0.001).






The number of live cells in cultures exposed to reduced PAR increased linearly throughout the incubation (Figure 1a), whereas those under ambient light had no additional growth after 10 days (F1,9=2.198, P=0.177), thus yielding ∼35% less live cells than reduced PAR treatments at day 13 (F1,8=11.6257, P=0.011). As these treatments had equal abundances at day 10, their nutrient consumption and rates of gas exchange would be similar. Therefore, the increase in light exposure must account for the lower abundance in ambient PAR treatments indicating a photo-inhibitory response somewhere above 48.6 μmol photons m−2 s−1. Similarly, Feng et al. (2013) observed photo-oxidative stress on P. torquis at 27.7 μmol photons m−2 s−1. At day 13, reduced PAR treatments contained over 40% more live cells (F1,12=11.224, P=0.006) than in ambient or blue treatments (F1,8=0.794, P=0.399). Green treatments differed from ambient and blue (F1,13=29.289, P<0.001), with only one third of the live cells in reduced PAR samples. Red exposed cultures were similar to dark treatments (F1,8=2.730, P=0.137), both of which yielded even fewer live cells than in green treatments (F1,13=35.758, P<0.001).

(a) Mean number (±s.e.m.) of live in situ incubated P. torquis cells. Treatments include exposure to ambient photosynthetically active radiation (), complete darkness (), reduced photosynthetically active radiation (), blue (), green () or red light (). Data points are offset (±3 h) for ease of viewing. Following incubation for 6, 10 or 13 days, 2 ml samples were fixed in 1% formaldehyde (Sigma-Aldrich, Auckland, New Zealand) and stored at −20 °C. After thawing, 1 ml of each was centrifuged (5000 g, 12 min), and the pellets air dried for ∼10 min. Cells were re-suspended in 350 μl of 0.22 μm filtered phosphate buffered saline (Life Technologies, Auckland, New Zealand) diluted 10 × and 350 μl was transferred to a 5 ml tube containing 35 μl of TruCount bead solution (Becton Dickinson). Each sample was incubated at room temperature for 15 min in 3 μm propidium iodide (Life Technologies). Prepared samples were analysed using a FACSCanto II flow cytometer (Becton Dickinson), acquiring data until 1000 TruCount bead events were recorded. Total cell counts were obtained through particle sizing, and intact and membrane-permeablised cells (live and dead at the time of fixation) were differentiated via the uptake of propidium iodide. (b) Mean number (±s.e.m.) of light-generated live cells per μmol photon present in each waveband treatment on day 13. The number of light-generated live cells of each sample was calculated by subtracting the number of live cells present in each light-exposed sample from the mean number of live cells present in dark incubations. The number of live cells under red light was similar to that in darkness, and so the red-light treatment was not included in this analysis. This value was then divided by the irradiance of their corresponding treatment to provide an estimate of the light-generated live cells per μmol photon for each light treatment.
The efficiency of each wavelength in promoting cellular growth was visualised by estimating the number of light-generated live cells per μmol photon (Figure 1b). Reduced PAR and blue light were the most efficient treatments (F1,7=3.827, P=0.091) with over three times more light-generated live cells than samples exposed to ambient PAR (F1,12=24.476, P<0.001). Green light however, was much less efficient, with 5.5 × fewer cells per photon than reduced PAR and blue-light samples and ∼45% fewer than ambient PAR treatments (F1,8=7.856, P=0.023). This indicates that cellular growth was not directly correlated with spectral intensity.
P. torquis PR contains methionine in position 105 as in other Bacteroidetes (Bowman et al., 1998; Koh et al., 2010), suggesting it should preferentially absorb green light (Riedel et al., 2010). Our results are therefore surprising. Blue and green treatments were exposed to a similar number of photons, removing differential photon exposure as a potential contributing factor. Furthermore, reduced PAR incubations were exposed to the same level of irradiation, and yielded a similar number of light-generated live cells as blue-light treatments. Because P. torquis is present throughout the entire ice column (Bowman et al., 1998) the utilisation of blue light rather than green would be more beneficial. The preference of this bacterium for blue light suggests that the capacity for PR to absorb a particular wavelength of light (Béjà et al., 2001; Man et al., 2003; Riedel et al., 2010; Mao et al., 2014), is controlled by more factors than this single amino acid shift. One such factor may be the abundance of carotenoids (such as β-carotene and zeaxanthin) within P. torquis (Bowman et al., 1998). These pigments have a major spectral absorbance between 425 and 525 nm (Feng et al., 2013). P. torquis carotenoids increase under light exposure (Feng et al., 2013), and PR function is dependent on the carotenoid derived chromophore, retinal (Béjà et al., 2001). P. torquis possesses both an unidentified carotenoid and retinal (Feng et al., 2013), similar to the retinal protein-carotenoid complex, xanthorhodopsin (Balashov et al., 2005). However, xanthorhodopsin has not been reported in P. torquis. Therefore, the carotenoids present in P. torquis may have a functional effect on the wavelength tuning of PR and merit further investigation.
The vast distribution of PR-bacteria indicates that PR may confer a selective advantage. However, their varied light responses suggest that this advantage is not as simple as increased growth under higher irradiances. The rate of cellular growth and the light–dark differences observed are similar to the experimental work performed by Feng et al. (2013). However, the growth differences in light and dark treatments were more extreme than expected, which indicates that environmental stressors such as temperature, may be having a larger influence on PR than initially thought. This work supplements the growing volume of evidence that PR provides additional energy to bacteria growing under sub-optimal conditions (Fuhrman et al., 2008; Feng et al., 2013). To our knowledge, this is the first study to demonstrate light-enhanced growth of a PR-bacterium using a realistic natural habitat. This insight into spectral tuning provides an exciting area for future research, potentially shifting the current understanding of the ecological significance of PR-bacteria.
References
Arrigo KR, Worthen DL, Lizotte MP, Dixon P, Dieckmann G . (1997). Primary production in Antarctic sea ice. Science 276: 394–397.
Balashov SP, Imasheva ES, Boichenko VA, Antón J, Wang JM, Lanyi JK . (2005). Xanthorhodopsin: a proton pump with a light-harvesting carotenoid antenna. Science 309: 2061–2064.
Béjà O, Aravind L, Koonin EV, Suzuki MT, Hadd A, Nguyen LP et al. (2000). Bacterial rhodopsin: evidence for a new type of phototrophy in the sea. Science 289: 1902–1906.
Béjà O, Spudich EN, Spudich JL, Leclerc M, DeLong EF . (2001). Proteorhodopsin phototrophy in the ocean. Nature 411: 786–789.
Bowman JP, McCammon SA, Lewis T, Skerratt JH, Brown JL, Nichols DS et al. (1998). Psychroflexus torquis gen. nov., sp. nov. a psychrophilic species from Antarctic sea ice, and reclassification of Flavobacterium gondwanense (Dobson et al. 1993) as Psychroflexus gondwanense gen. nov., comb. nov. Microbiology 144: 1601–1609.
Feng S, Powell SM, Wilson R, Bowman JP . (2013). Light-stimulated growth of proteorhodopsin-bearing sea-ice psychrophile Psychroflexus torquis is salinity dependent. ISME J 7: 2206–2213.
Finkel OM, Béjà O, Belkin S . (2013). Global abundance of microbial rhodopsins. ISME J 7: 448–451.
Fuhrman JA, Schwalbach MS, Stingl U . (2008). Proteorhodopsins: an array of physiological roles? Nature Rev Microbiol 6: 488–494.
Giovannoni SJ, Bibbs L, Cho J-C, Stapels MD, Desiderio R, Vergin KL et al. (2005). Proteorhodopsin in the ubiquitous marine bacterium SAR11. Nature 438: 82–85.
Gomez-Consarnau L, Gonzalez JM, Riedel T, Jaenicke S, Wagner-Dobler I, Sanudo-Wilhelmy SA et al. (2016). Proteorhodopsin light-enhanced growth linked to vitamin-B1 acquisition in marine Flavobacteria. ISME J 10: 1102–1112.
Gómez-Consarnau L, González JM, Coll-Lladó M, Gourdon P, Pascher T, Neutze R et al. (2007). Light stimulates growth of proteorhodopsin-containing marine Flavobacteria. Nature 445: 210–213.
Koh EY, Atamna-Ismaeel N, Martin A, Cowie ROM, Béjà O, Davy SK et al. (2010). Proteorhodopsin-bearing bacteria in Antarctic sea ice. Appl Environ Microbiol 76: 5918–5925.
Kottmeier ST, Sullivan CW . (1988). Sea ice microbial communities (SIMCO). Polar Biol 8: 293–304.
Lami R, Cottrell MT, Campbell BJ, Kirchman DL . (2009). Light‐dependent growth and proteorhodopsin expression by Flavobacteria and SAR11 in experiments with Delaware coastal waters. Environ Microbiol 11: 3201–3209.
Man D, Wang W, Sabehi G, Aravind L, Post AF, Massana R et al. (2003). Diversification and spectral tuning in marine proteorhodopsins. EMBO J 22: 1725–1731.
Mao J, Do N-N, Scholz F, Reggie L, Mehler M, Lakatos A et al. (2014). Structural Basis of the Green–Blue Color Switching in Proteorhodopsin as Determined by NMR Spectroscopy. J Am Chem Soc 136: 17578–17590.
Palovaara J, Akram N, Baltar F, Bunse C, Forsberg J, Pedrós-Alió C et al. (2014). Stimulation of growth by proteorhodopsin phototrophy involves regulation of central metabolic pathways in marine planktonic bacteria. Proc Natl Acad Sci USA 111: E3650–E3658.
Petrich C, Eicken H . (2009). Growth, structure and properties of sea ice. In: Thomas DN, Dieckmann GS (eds), Sea Ice. 2nd edn. John Wiley & Sons: Oxford.
Riedel T, Tomasch J, Buchholz I, Jacobs J, Kollenberg M, Gerdts G et al. (2010). Constitutive expression of the proteorhodopsin gene by a flavobacterium strain representative of the proteorhodopsin-producing microbial community in the North Sea. Appl Environ Microbiol 76: 3187–3197.
Ryan KG, Cowie ROM, Liggins E, McNaughtan D, Martin A, Davy SK . (2009). The short-term effect of irradiance on the photosynthetic properties of Antarctic fast-ice microbial communities. J Phycol 45: 1290–1298.
Smith RA . (1971). The effect of unequal group size on Tukey's HSD procedure. Psychometrika 36: 31–34.
Steindler L, Schwalbach MS, Smith DP, Chan F, Giovannoni SJ . (2011). Energy starved Candidatus Pelagibacter ubique substitutes light-mediated ATP production for endogenous carbon respiration. PLoS One 6: e19725.
Acknowledgements
We thank Associate Professor John Bowman (U. Tasmania) for supplying cultures of P. torquis, and acknowledge VUW Grant 100241 and Antarctica New Zealand for funding and logistical support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Burr, D., Martin, A., Maas, E. et al. In situ light responses of the proteorhodopsin-bearing Antarctic sea-ice bacterium, Psychroflexus torques. ISME J 11, 2155–2158 (2017). https://doi.org/10.1038/ismej.2017.65
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2017.65
