
Abstract
Two subspecies of the meadow grasshopper Chorthippus parallelus meet in a secondary hybrid zone in the Pyrenees. We have recently detected Wolbachia infection in this extensively studied species. C. p. parallelus (Cpp) and C. p. erythropus (Cpe) harbour bacteria from the B and F supergroups, but they differ noticeably in the incidence and type of infection present in their populations. We can distinguish different regional infection patterns that are associated with the distribution of pure and hybrid C. parallelus individuals. The northern pattern of low-level infection characterizes Cpp populations, whereas the southern pattern of high-level infection affects Cpe locations. These patterns converge in the hybrid zone and generate a third infection pattern featuring an extremely high degree of co-infection with the two Wolbachia types in hybrids. The distribution of Wolbachia among the studied populations encourages us to consider the possibility that this bacterium has a significant influence on the origin, maintenance and dynamics of this hybrid zone, given the reproductive alterations that are often associated with Wolbachia.
Similar content being viewed by others


Introduction
A hybrid zone is a region where two taxa meet, mate and leave hybrid offspring. These ‘natural laboratories’ allow us to study evolutionary agents, gene-flow barriers and the genetic divergence underlying them (Barton and Hewitt, 1985; Hewitt, 1988).
Two subspecies of the meadow grasshopper Chorthippus parallelus that arose in allopatry, C. parallelus parallelus (Cpp) and C. parallelus erythropus (Cpe), form a secondary hybrid zone in the Pyrenees. These grasshoppers diverged as a result of the geographic isolation that existed during and after the last Ice Age, before the ice finally disappeared from the mountain passes. Cpp is widely distributed throughout most of continental Europe, but in the Iberian Peninsula it is replaced by the endemic Cpe, which exhibits morphological, ethological, chromosomal and molecular peculiarities (reviewed in Shuker et al., 2005a).
The ecological requirements of these grasshoppers constrain their habitat to alpine-like prairies and this restricts their distribution in the Iberian Peninsula to the northern third and to certain mountain regions. The two subspecies meet and hybridize in transverse Pyrenean valleys, such as the Vall d’Ossau–Valle de Tena, in which contact between the endemic Iberian and the continental subspecies is possible.
Many studies have showed clines in this hybrid zone of morphological, ethological, chromosomal and molecular markers that differentiate the subspecies (Butlin and Hewitt, 1985a, 1985b; Gosálvez et al., 1988; Ritchie, 1990; Cooper et al., 1995; Lunt et al., 1998; Ibrahim et al., 2002; Bella et al., 2007). Furthermore, laboratory crosses between these subspecies produce male F1 hybrids that are sterile, exhibiting testicular dysfunction and meiotic anomalies (Hewitt et al., 1987; Bella et al., 1990; Virdee and Hewitt, 1994; Shuker et al., 2005b). These observations are consistent with an explanation of selection against natural hybrids, suggesting that this hybrid zone is a tension zone.
In this study, we describe the presence of a new factor that may be relevant to the origin, dynamics and maintenance of this hybrid zone. The genus Wolbachia (Hertig and Wolbach, 1924) is one of a group of alpha proteobacteria belonging to the order Rickettsiales. They have a strictly intracellular lifecycle and have been detected in a wide range of arthropods and nematodes (Jeyaprakash and Hoy, 2000; Hilgenboecker et al., 2008). They infect the germinal cell line by vertical transfer through a pattern of mainly maternal inheritance. Several different supergroups (clades A–H) have been defined on the basis of differences in various genes, such as 16S rDNA, ftsZ, wsp, and so on (Werren et al., 1995; Baldo et al., 2006).
These bacteria have been associated with certain reproductive disorders induced in parasitized arthropods (Werren, 1997; Weeks et al., 2002; Serbus et al., 2008) and with effects of Wolbachia strains that have a mutualistic lifestyle in nematodes (Bandi et al., 1999; Langworthy et al., 2000; Casiraghi et al., 2002) as side effects of their maintenance and transmission in populations. These disorders involve male feminization, parthenogenesis induction, male killing and cytoplasmic incompatibility (CI), the latter being the most common reproductive disorder in insects induced by Wolbachia. It is manifested as the reduction in the number of viable descendants from crosses between infected males and uninfected females, or between individuals infected by different strains (unidirectional and bidirectional CI, respectively). This may generate a significant reproductive barrier underlying the origin, maintenance and/or reinforcement of genetic differences among host subpopulations. This situation has led to a controversial model of Wolbachia-assisted speciation being proposed (Werren, 1998; Bordenstein, 2003; Telschow et al., 2005). The general assumption is that Wolbachia cannot be the primary cause of speciation events. However, there is recent strong evidence in Nasonia that reproductive isolation between its species is driven by Wolbachia (by bidirectional CI), mediated by some nuclear–cytoplasmic incompatibilities. Nevertheless, this evidence does not rule out the possibility that other pre- and post-zygotic isolation mechanisms also operate in this wasp (Gadau et al., 1999; Bordenstein et al., 2000; Beukeboom and van den Assem, 2001). This suggests that this alternative model of speciation should be examined in other well-studied complex systems, such as that of C. parallelus. The initial steps would involve establishing whether Wolbachia infection is present in populations inside and outside the hybrid zone, then examining whether they show similar levels and types of infection (which would indicate an analogous influence of the bacterium in all of them, and would discount any specific influence in the hybrid zone) and ruling out differences between the sexes and other factors that may mask the real influence of Wolbachia. Should differences among populations be found and potential confounding variables be assessed and discounted, further experiments would be required to test the role played by Wolbachia as a reproductive barrier and a speciation agent.
In this preliminary study, we describe B/F supergroup Wolbachia infection in C. parallelus and report differences between Cpp and Cpe populations, which have, respectively, low and high incidences of infection. The incidence of Wolbachia types differs between subspecies: the B supergroup is better represented in Cpp, whereas the F supergroup is widespread in Cpe. These differences generate a cline of infection through the Pyrenean hybrid zone, which we have described here. We have also been able to distinguish three patterns of infection: (i) northern: typically of Cpp and northern populations of the hybrid zone; (ii) southern: characteristic of Cpe populations and the southern edge of the hybrid zone; (iii) Sallent de Gállego: apparently a consequence of the contact between (i) and (ii) in the Pyrenees, and named after the location of the population in which it was first observed. Considering Wolbachia infection in the context of earlier studies of the C. parallelus hybrid zone leads us to propose that Wolbachia may be an important factor in the hybrid zone that structures populations and affects gene flow between subspecies.
Materials and methods
Sampling and DNA isolation
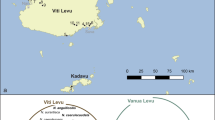
A 25-km transect across the entire width of the hybrid zone, running north-south through the Vall d’Ossau (France)–Valle de Tena (Spain) in the central Pyrenees, was selected for the study. Nine populations were sampled at points along the transect: Gabas, L’Hermine, Refugio, Tourmont, Cabane de l’Araille, Portalet, Corral de Mulas, Sallent de Gállego and Escarrilla. The population at Gabas, at the northern limit of the hybrid zone, is considered as pure Cpp (that is, the European continental subspecies of C. parallelus), while that at Escarrilla, at the southern limit, is comprised of pure Cpe (the endemic Iberian subspecies of C. parallelus). The mountains on either side of the valley exceed 2000 m, preventing the immigration of grasshoppers from neighbouring valleys. Other collection sites situated at various distances from the hybrid zone, towards the north or south of Europe, are considered as reference populations (Figure 1). Some populations were collected in more than 1 year to assess the infection over time. Otherwise, sampling effort was shared more broadly but less intensively among other populations in order to establish the cline from more points through the hybrid zone, which was the main aim of this research, or to determine the type of infection present in remote sites. Approximately 4700 individuals were analysed, similar proportions of males and females being collected from each population (see Tables 1 and 2 for sample sizes and locations).

Map of Wolbachia infections in the field. The circles show the average percentage of infection categories observed in each sampled population. (a) Populations outside the hybrid zone: ENG, England; MC, Massif Central; ARU, Arudy; BIE, Biescas; SAB, Sabiñánigo; NAV, Navafría; and BUB, Bubión. The solid line shows the border of C. p. parallelus (Cpp) and C. p. erythropus (Cpe) distributions. (b) Populations from the hybrid zone: GAB, Gabas; HER, L’Hermine; REF, Refugio; TOU, Tourmont; CA, Cabane de L’Araille; POR, Portalet; CM, Corral de Mulas; SAL, Sallent de Gállego and ESC, Escarrilla. The shading indicates mountains of more than 2000 m adjacent to the hybrid zone.
The gonads were extracted and preserved in ethanol at −20 °C until independent processing with the standard phenol–chloroform procedure for DNA isolation. The DNA was precipitated with ethanol and resuspended in TE buffer (Tris-HCl 1 mM, pH 8.0; EDTA 1 mM, pH 8.0), and then treated with RNAse H for 1 h at 37 °C.
PCR amplification, sequencing and phylogenetic analysis
We used the PCR to detect the presence of Wolbachia in C. parallelus. PCR was carried out with specific primers that amplify a fragment of ∼1400 bp from the 16S rDNA bacterial gene (Giordano et al., 1995). Each reaction mixture contained 200 ng extracted DNA, 5 μl reaction buffer, 1.5 mM MgCl2, 0.2 mM dNTPs (each nucleotide), 30 pmol each primer and 1 U Taq DNA polymerase in a 50-μl final volume. PCR cycling conditions consisted of 3 min at 95 °C, followed by 30 cycles of 30 s at 95 °C, 1 min at 54 °C, 2 min at 72 °C and a final cycle of 10 min at 72 °C.
A total of 50 amplified fragments from six representative populations (England, Arudy, Gabas, Portalet, Escarrilla and Navafría) were purified on columns (GFX PCR DNA Purification Kit, Amersham Biosciences, Freiburg, Germany) and sequenced using an Applied Biosystems 3730 Analyzer (Applied Biosystems, Foster City, CA, USA). Forward and reverse sequences were aligned in ClustalX 1.83 (Thompson et al., 1997) and the alignment manually inspected. Six variants of the bacterial gene were identified and deposited in GenBank (accession numbers: FJ438533, FJ438534, FJ438535, FJ438536, FJ438537 and FJ438538). Searching for homologous sequences in BLAST returned other Wolbachia 16S rDNA fragments, some of which (Table 3) were used for the phylogenetic analysis. With the 20 previously published sequences and the six sequences described here, an alignment of 1208 bp was obtained after removing non-overlapping tails in order to obtain similar fragment lengths.
The topology of the phylogenetic tree was constructed by Bayesian inference using the MrBayes 3.1 program (Ronquist and Huelsenbeck, 2003). The HKY+G model of sequence evolution was selected using hierarchical likelihood ratio tests available in the Modeltest 3.7 program (Posada and Crandall, 1998). The two independent runs performed, each with four chains, reached convergence of the split frequencies after 200 000 generations. Trees were sampled every 100 generations, giving a total of 4000 trees. The first 500 (25%) trees produced in each run were discarded as burn-in, leaving 3000 trees from which to estimate the 50% majority rule consensus and the Bayesian posterior probabilities.
Determination of infection frequencies
Our previous phylogenetic analysis indicated the existence in our populations of bacteria belonging to the B and F supergroups of Wolbachia; therefore, we adopted a nested PCR approach to determine accurately the infection frequencies of Wolbachia in C. parallelus populations. Nested PCR was performed with the aforementioned specific external primers. The products of general PCR were subjected to a second round of amplification with specific internal primers. This two-part specific amplification approach is useful for detecting Wolbachia endoparasites because it improves specificity and avoids false negatives arising from low bacterial density. The combination of F6 (5′-GTGAAGAAGGCCTTTGGGTTGTA-3′) and R10B (5′-GATTAGCTTAGGCTTGCGCACC-3′) oligomers enables B supergroup strains to be amplified, whereas the F6 and R11F pair (5′-GATTAGCTTAGCCTCGCGACTT-3′) detects F supergroup bacteria. These two combinations result in an approximately 850-bp fragment length and enable B and F supergroup infections to be discriminated. The reaction conditions were largely the same as those outlined above except that 1 μl PCR product was added instead of 200 ng DNA. The PCR cycling conditions consisted of 3 min at 95 °C followed by 30 cycles of 30 s at 95 °C, 1 min at 61 °C, 1 min at 72 °C, and a final cycle of 10 min at 72 °C. In addition, the procedure was tested with closely related bacteria, such as Rickettsia sp., Rhizobium sp., Pseudomonas fluorescens, Escherichia coli, as well as chicken, turkey and human DNA, none of which yielded positive results. In all cases and analyses, PCR was performed with a known infected DNA sample as a positive control and water as a blank.
Statistical analyses
Differences in infection between sexes
The incidence of infection in males and females was compared in each of the 45 samples by analysing the cross-tabulated frequencies of sex (male/female) by infection category (uninfected/B-infected/F-infected/co-infected). Likelihood ratios were calculated on the basis of these tables using the G-test. As 45 tests were carried out, we applied a Bonferroni correction. In this way, at the experiment-wide 95% confidence level, the critical probability for rejecting the null hypothesis in an individual test was set at α=0.001.
Temporal differences in the infection of populations
The homogeneity of the infection frequencies over four generations (2002–2006) was examined by analysing the cross-tabulated frequencies of the four categories of infection observed in a population over distinct sampling years. Once again, likelihood ratios were calculated using the G-test.
Determining the different patterns of infection
A hierarchical cluster analysis was carried out in which the 45 samples in this study were considered independently of their site of origin and year of sampling. To this end, a distance matrix was constructed with the values of χ2 obtained for each possible pair of samples using the frequencies of the infection categories found in each sample. Applying the furthest-neighbour method (among others) to this matrix, a dendrogram was generated from which we identified the various possible groupings. To take into account the possible undesirable influence on the results of some small sample sizes, the analysis was repeated excluding those with fewer than 40 or 80 individuals. Outcomes were not substantially altered, and so it is most informative to include all samples in the analyses reported here. Non-parametric tests (Kruskal–Wallis test and Mann–Whitney U-test) were used to compare the distribution of proportions of each infection category (uninfected, B-infected, F-infected and co-infected) between the patterns described below (see the Results section). Once again, the exclusion of small samples produced no substantial differences in the results and these have therefore been retained.
All statistical procedures were carried out using SPSS 15.0.
Results
Wolbachia strains in C. parallelus
We detected six 16S rDNA variants in C. parallelus (wCpar_01, wCpar_02, wCpar_03, wCpar_04, wCpar_05 and wCpar_06). They featured 52 variable positions over 1305 bp that allowed them to be grouped into two sets, the first including wCpar_01 and wCpar_02, with only two variable positions, and the second including wCpar_03, wCpar_04, wCpar_05 and wCpar_06, with 13 variable positions. There was more than 99% homology within groups, but sequence similarity was <97% between groups.
The Bayesian inference tree based on the 1208-bp alignment obtained with published sequences and our sequences showed that wCpar_01 and wCpar_02 variants belong to the B supergroup, whereas wCpar_03, wCpar_04, wCpar_05 and wCpar_06 are included in the F supergroup (Figure 2) (see Werren et al., 1995; Lo et al., 2002; Serbus et al., 2008). The Bayesian posterior probabilities of these results are 100 and 80–87% for the B and F supergroup nodes, respectively. The topology is consistent with that of previous phylogenies (Lo et al., 2002).

Unrooted phylogenetic tree based on 16S rDNA gene of Wolbachia, derived using the Bayesian inference method. The letters A–F indicate supergroups and the numbers are the Bayesian posterior probabilities. The names of taxa are those of the host and are accompanied by the accession number of their bacterial 16S rDNA sequences. wCpar and Wolbachia strains of C. parallelus found in this study.
Wolbachia infection in C. parallelus
All the populations of C. parallelus analysed in this study were infected with Wolbachia, although there were differences in infection type and incidence between northern and southern populations. Northern European locations had a low incidence of infection, whereas most grasshoppers in Spanish populations carried Wolbachia. Through the hybrid zone, there was a north–south gradient from low to high levels of infection (Figure 1).
The infection type and frequencies in populations outside the hybrid zone are shown in Table 1. There was no population without some kind of Wolbachia infection, and it was also possible to find variable proportions of individuals co-infected with B and F bacterial supergroups. This allows individuals to be classified in one of four infection categories: uninfected, B-infected, F-infected and co-infected. Infection is widespread in Cpe populations, where most of the samples featured 76–100% infected individuals. However, this percentage was <43% in all Cpp locations except for England, where 60% of the individuals were infected. Iberian populations were mainly infected by F supergroup strains; 1.5–15 times as many individuals harboured these bacteria as did those of the B supergroup. In contrast, Cpp locations featured individuals that were equally or up to three times as likely to harbour B supergroup strains as F supergroup bacteria. Our results do not allow us to confirm the independence of infection by the two Wolbachia strains. In general, there was a higher rate of co-infection overall than would be expected from the separate frequencies of B and F infection, although this pattern was not seen in all years or in all the populations considered.
We checked for differences in the incidence of infection between the sexes in all 45 samples included in this study (Tables 1 and 2). There were no statistically significant differences between the sexes in any sample except for those from Portalet (2002) and Sallent de Gállego (2006) (G=16.08, d.f.=2, P<0.001; G=15.52, d.f.=2, P<0.001, respectively). In both cases, these differences consisted of a lower than expected incidence of B infection and an excess of uninfected males with respect to females.
Some representative populations inside and outside the hybrid zone were monitored in consecutive years from 2002 to 2006 (Tables 1 and 2) in order to detect annual variation in infection frequencies. With the exception of the Sabiñánigo population, where the F infection was almost universal, all populations exhibited statistically significant differences between years, characterized by erratic rises and falls, rather than any consistent trend. In spite of infection heterogeneity between years, there were no infection differences between the first and the last year in the extremes of the hybrid zone after four generations (Gabas: G=2.23, d.f.=3, P=0.527; Escarrilla: G=5.47, d.f.=3, P=0.141). However, there were differences in intermediate populations during the same period (Portalet: G=18.87, d.f.=3, P<0.001; Sallent de Gállego: G=15.80, d.f.=3, P<0.001). We inspected the adjusted standardized residuals of the contingency cross-tabulations to determine which infection category contributed most to the significant differences within each population and its direction. The proportion of uninfected individuals increased by 17.2% between the first and fourth years of sampling in Portalet, whereas that of co-infected individuals decreased by 9.9% in the same population. In Sallent de Gállego, individuals harbouring both strains became proportionally more common (an increase of 14.2%) over the 4 years of sampling, whereas the incidence of F infection decreased by 11.3%.
Patterns of Wolbachia infection and their spatial distribution
The different infection frequencies between and within populations prompted us to conduct a blind analysis in order to classify independently all the 45 samples collected over the years according to their infection category frequencies. We carried out a hierarchical cluster analysis, generating a distance matrix for all possible sample pairs and a dendrogram was produced (Figure 3). It was possible to distinguish three clusters corresponding to different infection patterns. Differences between samples pooled in the three groups (Gpooled=2520.903, d.f.=6, P<0.001) accounted for 74.7% of the total entropy observed across all samples (Gtotal=3376.757, d.f.=132 P<0.001), whereas heterogeneity within clusters (Gheterogenity=855.855, d.f.=126, P<0.001) accounted for only 25.3% of the total differences among sample distributions. Indeed, non-parametric analysis of variance showed differences between the three patterns in ranked proportion for all infection classes (Kruskal–Wallis test: uninfected, H=31.668, d.f.=2, P<0.001; B-infected, H=21.104, d.f.=2, P<0.001; F-infected, H=28.328, d.f.=2, P<0.001; co-infected, H=19.075, d.f.=2, P<0.001). Populations sampled in several years were always classified in the same cluster irrespective of the sampling year. One cluster included populations from the north of Europe and almost the entire transect except for those from Sallent de Gállego and Escarrilla at its southern end. Another cluster was composed of the south and central Spanish populations, Escarrilla and populations from the Spanish Pyrenees outside the transect. The third cluster featured solely Sallent de Gállego samples. The distributions of infection proportions for the three clusters are shown in Figure 4. Populations included in the northern pattern of infection showed a great predominance of uninfected individuals. In contrast, the most abundant category in the southern infection pattern was of F-infected individuals, and co-infected individuals were in the majority in the Sallent de Gállego pattern. B and F infection groups were similarly represented in northern and Sallent de Gállego populations (Mann–Whitney U-test: B-infected, U=45, P=0.243; F-infected, U=38, P=0.126), that is, a slightly higher incidence of B infection and a much lower incidence of F infection than in the southern pattern. By contrast, the Sallent and southern patterns had a similar average proportion of uninfected individuals (Mann–Whitney U-test: uninfected, U=21, P=0.256) that was very low in comparison with the northern pattern (Figure 4).

Dendrogram obtained from hierarchical cluster analysis using a furthest-neighbour method. Three clusters are differentiated, indicated by the solid, dashed and pale lines. The samples are listed by their population abbreviation and the year of sampling. ENG, England; MC, Massif Central; ARU, Arudy; BIE, Biescas; SAB, Sabiñánigo; NAV, Navafría; and BUB, Bubión. Populations from the hybrid zone: GAB, Gabas; HER, L’Hermine; REF, Refugio; TOU, Tourmont; CA, Cabane de L’Araille; POR, Portalet; CM, Corral de Mulas; SAL, Sallent de Gállego and ESC, Escarrilla.

Box graphs of infection category proportions for the three infection patterns identified by the hierarchical cluster analysis. The rectangles indicate the 50% of values between the 25th and 75th percentiles. The lines inside the rectangles indicate medians. Whiskers show the lowest and highest values, although outliers (cases with a value >1.5 times the box length) are indicated outside the rectangles by circles.
Infection distribution in the hybrid zone
There was a change through the transect, from the northern infection pattern (from Gabas to Corral de Mulas) to the southern one (Escarrilla, see above), whereas a new pattern in Sallent de Gállego, with its own features, connected the northern and southern patterns (Figure 5). Uninfected individuals were the predominant category from Gabas to Corral de Mulas (Figure 5a), but their proportion began to fall between Corral de Mulas and Sallent de Gállego, where uninfected individuals made up only about 4% of the population. In Escarrilla, the uninfected class accounted for 23% of the individuals sampled. The incidence of the B-infection category (Figure 5b) was around 6% in Gabas. This value was higher at L’Hermine, where the average was around 20%. Similar values were found until Sallent de Gállego. About 6% of individuals were B-infected in Escarrilla. The incidence of F infection (Figure 5c) was very similar from Gabas to Sallent de Gállego, at around 10%, but the average reached 45% in Escarrilla. Co-infection was most common in Sallent de Gállego (Figure 5d), where individuals harbouring both supergroup strains accounted for about 68% of the total. The average was lower towards the north and south (∼12 and 26%, respectively).

Spatial distribution of infection frequencies through the hybrid zone, following the four infection categories described in the text. (a) Uninfected, (b) B-infected, (c) F-infected and (d) co-infected individuals. Several years of sampling may be represented for particular locations. The solid line shows the average proportion observed in each population. G, Gabas; H, L'Hermine; R, Refugio; T, Tourmont; CA, Cabane de L'Araille; P, Portalet; CM, Corral de Mulas; S, Sallent de Gállego; and E, Escarrilla. The vertical axis shows the proportions and horizontal axis shows the distance in kilometres.
Discussion
There were moderate to severe Wolbachia infections in the populations of pure and hybrid individuals of the grasshopper C. parallelus included in this study. Molecular screening of distinct samples showed variability in the 16S rDNA sequence of the Wolbachia gene, allowing us to distinguish two groups belonging to the B and F Wolbachia supergroups. The B supergroup strains are well characterized and are known to infect other arthropods. Their parasitic lifestyle often induces alterations in host reproduction, such as CI. However, F supergroup strains have been found in arthropods and nematodes, where their lifestyle and the effects on their hosts are uncertain (Werren, 1997; Lo et al., 2002; Rasgon and Scott, 2004; Casiraghi et al., 2005). C. parallelus are often parasitized by nematodes and nematomorphs. Risk of contamination in our study was minimized using only the DNA from the gonads for PCR analysis.
There were no sex-related differences in Wolbachia infection in the populations analysed, with the exception of two samples. An excess of infected females might be expected in the case of a system with male feminization, selective male killing or parthenogenetic effects produced by Wolbachia. Therefore, we conclude that this bacterium does not promote these effects in the populations of C. parallelus studied here. This is supported by direct observations in natural and laboratory populations of this grasshopper, where such effects have not been described in any previous study. Wolbachia infection appears not to show stable temporal trends in this species, but this might be expected as this bacterium seems to be affected by various temporal and environmental characteristics: infection density, temperature, immune defence of the host, seasonal and ecological factors, and so on (Werren, 1997; Serbus et al., 2008). We recognize that, as these grasshoppers have an annual life cycle, we have not studied sufficient generations to be able to discern any clear trends due to bacterial effects amidst the relatively large and erratic shifts in infection proportions.
Infection incidence differs between Cpp and Cpe, but there are similarities within populations of the pure subspecies (locations outside the hybrid zone; Figure 1). Thus, the infections observed in Cpp and Cpe, respectively, characterize a northern and a southern infection pattern (Figure 3). We do not know whether this distribution is a consequence of a latitudinal gradient (reflecting those of humidity, temperature, insolation, etc.) or whether it is the consequence of independent infection events. We intend to investigate the origin of Wolbachia infection in these grasshoppers and to determine whether the infection arose before or after the geographical isolation of the taxa forming the hybrid zone; our current data do not enable us to draw any conclusions about this. However, given that Iberian Cpe and continental Cpp differ in Wolbachia infection pattern, we do not think it likely to be ancestral (that is, before divergence in allopatry), which makes it possible that it may have affected even the origin of the hybrid zone.
The populations from the hybrid zone describe a transitional route between the northern and southern infection patterns. However, the infection shifts were mainly found on the Iberian side of the Pyrenees, in the segment between Escarrilla and Corral de Mulas (Figure 5). Furthermore, Sallent de Gállego is the place where both infection patterns coincide to generate a third pattern in which the level of co-infection was the highest recorded. As Wolbachia has maternally inherited transmission, the co-infection peak at Sallent de Gállego implies strong positive selection for individuals harbouring both bacterial supergroup infections. Co-infected females could have more viable offspring in a two-strain model with bidirectional CI, thus the proportion of co-infected individuals would increase generation after generation until the infection frequency reaches a stable equilibrium (see Frank, 1998, for this and other hypotheses). This equilibrium is generated by two opposite forces: the fertility advantage of co-infected females and the cost to fecundity of being infected (Turelli, 1994). Furthermore, co-infected females would only be favoured under specific conditions of infection frequencies. Those conditions could have prevailed in Sallent de Gállego but not in the rest of populations, where most individuals were characterized by the absence of infection or by infection with the F strain. This increase may also be explained by paternal and horizontal transmission but, although this has been described and cannot be definitively ruled out in this case, it is unlikely to be a sufficiently common event to explain the observed co-infection frequencies (Werren, 1997; Baldo et al., 2008; Serbus et al., 2008).
The main aim of this study was to determine whether Wolbachia could contribute a role in the dynamics of the C. parallelus hybrid zone in the Pyrenees. It is difficult to predict what one might expect from Wolbachia following secondary contact. We have already noted the evidence from Nasonia of reproductive isolation among species driven by Wolbachia. In this case, other reproductive barriers also operate, indicating that simple hypotheses are not adequate to explain complicated interactions with selection operating on phenotypes resulting from nuclear–nuclear, nuclear–cytoplasmic and cytoplasmic–cytoplasmic interactions (Bordenstein et al., 2003; Opijnen et al., 2005; Ellison et al., 2008; Oliveira et al., 2008). The clinal distribution of Wolbachia strain infections along a north–south gradient in the populations here studied and the overall decrease of the infection levels across this transect (Figure 1) are striking. Nevertheless, these alone are not evidence that Wolbachia acts as a reproductive barrier in this system, although it is very intriguing that the peak incidence of co-infection in Sallent de Gállego coincides with those of some cytogenetic markers described in previous studies (Gosálvez et al., 1988; Serrano et al., 1996), which reported a shift from 100 to 0% in the frequency of a distal heterochromatic band in the X chromosome (P-band) characteristic of Cpp between Corral de Mulas and Sallent de Gállego. Another study proposed that there was an incompatibility between this P-band region and cytoplasmic factors presumed to be present in Cpe cells (Virdee and Hewitt, 1992). If we assume preferential maternal inheritance of Wolbachia, the transmission of bacteria would be associated with the transmission of the cytoplasm and its factors from the mother. Some studies have shown that a Wolbachia-induced population sweep could reduce mitochondrial variability (Turelli et al., 1992; Dean et al., 2003). Furthermore, the Sallent de Gállego co-infection peak could have brought about the near fixation of the Cpe cytotype in that population and might constitute a barrier to the introgression of Cpp traits, as suggested by the behaviour of the P-band in this X-chromosome. Further experiments will distinguish which of these hypotheses provides the best explanation of the patterns observed.
As stated above, in Nasonia standard forms of reproductive isolation do not preclude a role for Wolbachia as a strong reproductive barrier (Bordenstein et al., 2001). Indeed, Wolbachia might even precede them, not discounting the possibility that once bidirectional CI and/or nuclear–cytoplasmic incompatibility due to Wolbachia has arisen, it could also affect those ‘orthodox’ mechanisms themselves, influencing patterns of nuclear–nuclear incompatibility and mediating the selection against deleterious genotypes (D Shuker, personal communication). This is particularly relevant here as genetic variation in post-zygotic incompatibilities among C. parallelus subspecies was attributed to neutral alleles, or those nearly neutral against their own genetic background, that were responsible for incompatibilities and hybrid sterility (Shuker et al., 2005b). This pattern might also be explained by Wolbachia acting as a reproductive isolating mechanism, or promoting nuclear–cytoplasmic interactions (see Bordenstein et al., 2003; Jaenike et al., 2006).
In addition, asymmetrical homogamy has been observed in laboratory crosses involving sequential matings with males of both pure C. parallelus subspecies (Bella et al., 1992). An excess of pure-like descendants was obtained in crosses with Cpp mothers, whereas approximately equal proportions of pure and hybrid descendants were seen in crosses with Cpe mothers. The Cpp individuals used in these crosses were from Gabas, where currently 70% of the individuals are not infected, whereas Cpe grasshoppers were from Escarrilla, where 77% of individuals are currently infected. This excess of pure-like descendants may reflect Wolbachia-induced CI because of the crossing of infected Cpe males and uninfected Cpp females (assuming similar types and levels of Wolbachia infection).
Our results do not represent evidence that Wolbachia acts as a barrier to reproduction in the hybrid zone of C. parallelus, although overall they paint an intriguing picture that is consistent with such a possibility. Our continuing research includes a programme of crosses between infected and uninfected individuals from the hybrid zone, which should enable us to resolve this matter. We also hope to be able to determine the consequences of Wolbachia infection on reproduction in this grasshopper, its transmission rate and the correlations with other variables and traits (for example, mitochondrial phylogeography, chromosomal, morphometric and behavioural traits) involved in its hybrid zone.
Our results show the distribution of Wolbachia in this insect and reveal geographical patterns of distribution of the bacterial strains that may also appear in other organisms. This study also provides the first evidence of the hitherto undescribed presence of the Wolbachia F strain in orthopteroids and enables us to derive an informative new phylogenetic tree.
References
Baldo L, Dunning Hotopp JC, Jolley KA, Bordenstein SR, Biber SA, Choudhury RR et al. (2006). Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl Environ Microbiol 72: 7098–7110.
Baldo L, Ayoub NA, Hayashi CY, Russell JA, Stahlhut JK, Werren JH (2008). Insight into the routes of Wolbachia invasion: high levels of horizontal transfer in the spider genus Agelenopsis revealed by Wolbachia strain and mitochondrial DNA diversity. Mol Ecol 17: 557–569.
Bandi C, McCall JW, Genchi C, Corona S, Venco L, Sacchi L (1999). Effects of tetracycline on the filarial worms Brugia phangi and Diofilaria immitis and their bacterial endosymbionts Wolbachia. Int J Parasitol 29: 357–364.
Barton NH, Hewitt GM (1985). Analysis of hybrid zones. Ann Rev Ecol Syst 16: 113–148.
Bella JL, Hewitt GM, Gosálvez J (1990). Meiotic imbalance in laboratory produced hybrid males of Chorthippus parallelus parallelus and Chorthippus parallelus erythropus. Genet Res 56: 43–48.
Bella JL, Butlin RK, Ferris C, Hewitt GM (1992). Asymmetrical homogamy and unequal sex ratio from reciprocal mating order crosses between Chorthippus parallelus subspecies. Heredity 68: 345–352.
Bella JL, Serrano L, Orellana J, Mason PL (2007). The origin of the Chorthippus parallelus hybrid zone: chromosomal evidence of multiple refugia for Iberian populations. J Evol Biol 20: 568–576.
Beukeboom LW, van den Assem J (2001). Courtship and mating behaviour of interspecific Nasonia hybrids (Hymenoptera: Pteromalidae): a grandfather effect. Behav Genet 31: 167–177.
Bordenstein SR, Drapeau MD, Werren JH (2000). Intraspecific variation in sexual isolation in the jewel wasp Nasonia. Evolution 54: 567–573.
Bordenstein SR, O’Hara FP, Werren JH (2001). Wolbachia-induced incompatibility precedes other hybrid incompatibilities in Nasonia. Nature 409: 707–710.
Bordenstein SR (2003). Symbiosis and the origin of species. In: Bourtizs K, Miller T (eds). Insect Symbiosis. CRC Press: Boca Raton, Florida, pp 283–304.
Bordenstein SR, Uy JJ, Werren JH (2003). Host genotype determines cytoplasmic incompatibility type in the haplodiploid genus Nasonia. Genetics 163: 223–233.
Butlin RK, Hewitt GM (1985a). A hybrid zone between Chorthippus parallelus parallelus and Chorthippus parallelus erythropus (Orthoptera: Acrididae): morphological and electrophoretic characters. Biol J Linn Soc 26: 269–285.
Butlin RK, Hewitt GM (1985b). A hybrid zone between Chorthippus parallelus parallelus and Chorthippus parallelus erythropus (Orthoptera: Acrididae): behavioural characters. Biol J Linn Soc 26: 287–299.
Casiraghi M, McCall JW, Simoncini L, Kramer LH, Sacchi L, Genchi C et al. (2002). Tetracycline treatment and sex–ratio distortion: a role for Wolbachia in the moulting of filarial nematodes? Int J Parasitol 32: 1457–1468.
Casiraghi M, Bordenstein SR, Baldo L, Lo N, Beninati T, Wernegreen JJ et al. (2005). Phylogeny of Wolbachia pipientis based on gltA, groEL and ftsZ gene sequences: clustering of arthropod and nematode symbionts in the F supergroup, and evidence for further diversity in the Wolbachia tree. Microbiology 151: 4015–4022.
Cooper SJ, Ibrahim KM, Hewitt GM (1995). Postglacial expansion and genome subdivision in the European grasshopper Chorthippus parallelus. Mol Ecol 4: 49–60.
Dean MD, Ballard KJ, Glass A, Ballard JWO (2003). Influence of two Wolbachia strains on population structure of East African Drosophila simulans. Genetics 165: 1959–1969.
Ellison CK, Niehuis O, Gadau J (2008). Hybrid breakdown and mitochondrial dysfunction in hybrids of Nasonia parasitoids wasps. J Evol Biol 21: 1844–1851.
Frank SA (1998). Dynamics of cytoplasmic incompatibility with multiple Wolbachia infections. J Theor Biol 192: 213–218.
Gadau J, Page Jr RE, Werren JH (1999). Mapping of hybrid incompatibility loci in Nasonia. Genetics 153: 1731–1741.
Giordano R, O’Neill SL, Robertson HM (1995). Wolbachia infections and the expression of cytoplasmic incompatibility in Drosophila sechellia and D. Mauritania. Genetics 140: 1307–1317.
Gosálvez J, López-Fernández C, Bella JL, Butlin RK, Hewitt GM (1988). A hybrid zone between Chorthippus parallelus parallelus and Chorthippus parallelus erythropus (Orthoptera: Acrididae): chromosomal differentiation. Genome 30: 656–663.
Hertig M, Wolbach SB (1924). Studies on Rickettsia-like microorganism in insect. J Med Res 44: 329–374.
Hewitt GM (1988). Hybrid zones: natural laboratories for evolutionary studies. Trends Ecol Evol 3: 158–167.
Hewitt GM, Butlin RK, East TM (1987). Testicular dysfunction in hybrids between parapatric subspecies of the grasshopper Chorthippus parallelus. Biol J Linn Soc 31: 25–34.
Hilgenboecker K, Hammerstein P, Schlattmann P, Telschow A, Werren JH (2008). How many species are infected with Wolbachia? A statistical analysis of current data. FEMS Microbiol Lett 281: 215–220.
Ibrahim KM, Cooper SJ, Hewitt GM (2002). Testing for recombination in a short nuclear DNA sequence of the European meadow grasshopper Chorthippus parallelus. Mol Ecol 11: 583–590.
Jaenike J, Dyer KA, Cornish C, Minhas MS (2006). Asymmetrical reinforcement and Wolbachia infection in Drosophila. PLoS Biol 4: e325.
Jeyaprakash A, Hoy MA (2000). Long PCR improves Wolbachia DNA amplification: wsp sequences found in 76% of sixty-three arthropod species. Insect Mol Biol 9: 393–405.
Langworthy NG, Renz A, Mackenstedt U, Henkle-Dührsen K, de Bronsvoort MB, Tanya VN et al. (2000). Macrofilaricidal activity of tetracycline against the filarial nematode Onchocerca onchengi: elimination of Wolbachia precedes worm death and suggests a dependent relationship. Proc Biol Sci 267: 1063–1069.
Lo N, Casiraghi M, Salati E, Bazzochi C, Bandi C (2002). How many Wolbachia supergroups exist? Mol Biol Evol 19: 341–346.
Lunt DH, Ibrahim KM, Hewitt GM (1998). mtDNA phylogeography and postglacial patterns of subdivision in the meadow grasshopper Chorthippus parallelus. Heredity 80: 633–641.
Oliveira DC, Raychoudhury R, Lavrov DV, Werren JH (2008). Rapidly evolving mitochondrial genome and directional selection in mitochondrial genes in the parasitic wasp Nasonia (Hymenoptera: Pteromalidae). Mol Biol Evol 25: 2167–2180.
Opijnen T, Baudry E, Baldo L, Bartos J, Werren JH (2005). Genetic variability in the three genomes of Nasonia: nuclear, mitochondrial and Wolbachia. Insect Mol Biol 14: 653–663.
Posada D, Crandall KA (1998). Modeltest: testing the model of DNA substitution. Bioinformatics 14: 817–818.
Rasgon JL, Scott TW (2004). Phylogenetic characterization of Wolbachia symbionts infecting Cimex lectularius L. and Oeciacus vicarius Horvath (Hemiptera: Cimicidae). J Med Entomol 41: 1175–1178.
Ritchie MG (1990). Does song contribute to assortative mating between subspecies Chorthippus parallelus (Orthoptera: Acrididae)? Anim Behav 39: 685–691.
Ronquist F, Huelsenbeck JP (2003). MrBayes3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572–1574.
Serbus LR, Casper-Lindley C, Landmann F, Sullivan W (2008). The genetics and cell biology of Wolbachia–host interactions. Annu Rev Genet 42: 683–707.
Serrano L, García de la Vega C, Bella JL, López-Fernández C, Hewitt GM, Gosálvez J (1996). A hybrid zone between two subspecies of Chorthippus parallelus. X-chromosome variation through a contact zone. J Evol Biol 9: 173–184.
Shuker DM, King TM, Bella JL, Butlin RK (2005a). The genetic basis of speciation in a grasshopper hybrid zone. In: Fellowes M, Holloway G, Roff J (eds). Insect Evolutionary Biology. CABI Publishing, Oxford University Press: Wallingford, Oxon, UK, pp 427–454.
Shuker DM, Underwood K, King TM, Butlin RK (2005b). Patterns of male sterility in a grasshopper hybrid zone imply accumulation of hybrid incompatibilities without selection. Proc R Soc B 272: 2491–2497.
Telschow A, Hammerstein P, Werren JH (2005). The effects of Wolbachia versus genetic incompatibilities on reinforcement and speciation. Evolution 59: 1607–1619.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997). The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 24: 4876–4882.
Turelli M (1994). Evolution of incompatibility-inducing microbes and their hosts. Evolution 48: 1500–1513.
Turelli M, Hoffmann AA, McKechnie SW (1992). Dynamics of cytoplasmic incompatibility and mtDNA variation in natural Drosophila simulans populations. Genetics 132: 713–723.
Virdee SR, Hewitt GM (1992). Postzygotic isolation and Haldane's rule in a grasshopper. Heredity 69: 527–538.
Virdee SR, Hewitt GM (1994). Clines for hybrid dysfunction in a grasshopper hybrid zone. Evolution 48: 392–407.
Weeks AR, Reynolds KT, Hoffmann AA (2002). Wolbachia dynamics and host effects: what has (and has not) been demonstrated? Trends Ecol Evol 17: 257–262.
Werren JH (1997). Biology of Wolbachia. Annu Rev Entomol 42: 587–609.
Werren JH (1998). Wolbachia and speciation. In: Howard D, Berlocher L (eds). Species and Speciation. Oxford University Press: Oxford. pp 245–260.
Werren JH, Zhang W, Guo LR (1995). Evolution and phylogeny of Wolbachia: reproductive parasites of arthropods. Proc Biol Sci 261: 55–63.
Acknowledgements
We are grateful to the many people who helped in the collection and handling of the grasshoppers and in the analyses of the data: PL Mason, C López-Fernández, J Orellana, J García-Hurtado, J Sarasa, P Martínez-Rodríguez, E Rolán-Alvarez, RK Butlin, JR Dagley, M Charalambous, RM Santos, J Bella and the other members of our group in the Genetics Unit at the Universidad Autónoma de Madrid. A Colas and A Peire collaborated in the earliest stages of this study. We thank D Shuker for his insightful comments on a previous version of our paper. We also express gratitude to the Spanish Junta de Andalucía, the Comunidad de Madrid, the Gobierno de Aragón and the French Parc National des Pyrénées (especially its Director, R Kbaier) for permitting to collect the grasshoppers. This work has been supported by the Spanish Grants BOS2002/0232 and CGL2005-02898.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zabal-Aguirre, M., Arroyo, F. & Bella, J. Distribution of Wolbachia infection in Chorthippus parallelus populations within and beyond a Pyrenean hybrid zone. Heredity 104, 174–184 (2010). https://doi.org/10.1038/hdy.2009.106
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/hdy.2009.106
Keywords
This article is cited by
-
16S rRNA Sequencing Detected Profftella, Liberibacter, Wolbachia, and Diplorickettsia from Relatives of the Asian Citrus Psyllid
Microbial Ecology (2020)
-
Transcriptome assembly for a colour-polymorphic grasshopper (Gomphocerus sibiricus) with a very large genome size
BMC Genomics (2019)
-
Widespread hybridization among native and invasive species of Operophtera moths (Lepidoptera: Geometridae) in Europe and North America
Biological Invasions (2019)
-
Geographic and Temporal Variation of Distinct Intracellular Endosymbiont Strains of Wolbachia sp. in the Grasshopper Chorthippus parallelus: a Frequency-Dependent Mechanism?
Microbial Ecology (2019)
-
Differential immunity as a factor influencing mussel hybrid zone structure
Marine Biology (2019)
