- TECHNOLOGY FEATURE
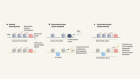
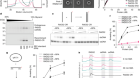
Got mutation? ‘Base editors’ fix genomes one nucleotide at a time
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 575, 553-555 (2019)
doi: https://doi.org/10.1038/d41586-019-03536-x
References
Komor, A. C., Kim, Y. B., Packer, M. S., Zuris, J. A. & Liu, D. R. Nature 533, 420–424 (2016).
Gaudelli, N. M. et al. Nature 551, 464–471 (2017).
Liu, Z. et al. Nature Commun. 9, 2338 (2018).
Liu, Z. et al. Nature Commun. 9, 2717 (2018).
Kim, K. et al. Nature Biotechnol. 35, 435–437 (2017).
Ryu, S.-M. et al. Nature Biotechnol. 36, 536–539 (2018).
Kuscu, C. et al. Nature Methods 14, 710–712 (2017).
Billon, P. et al. Mol. Cell 67, 1068–1079 (2017).
Grünewald, J. et al. Nature 569, 433–437 (2019).
Zuo, E. et al. Science 364, 289–292 (2019).
Jin, S. et al. Science 364, 292–295 (2019).
Rees, H. A. et al. Nature Commun. 8, 15790 (2017).
Kim, Y. B. et al. Nature Biotechnol. 35, 371–376 (2017).
Jiang, W. et al. Cell Res. 28, 855–861 (2018).
Kim, H. S., Jeong, Y. K., Hur, J. K., Kim, J.-S. & Bae, S. Nature Biotechnol. 37, 1145–1148 (2019).
Anzalone, A. V. et al. Nature https://doi.org/10.1038/s41586-019-1711-4 (2019).
Rees, H. A. & Liu, D. R. Nature Rev. Genet. 19, 770–788 (2018).
 CRISPR gene editing produces unwanted DNA deletions
CRISPR gene editing produces unwanted DNA deletions
 David Liu: Gene corrector
David Liu: Gene corrector
 Super-precise new CRISPR tool could tackle a plethora of genetic diseases
Super-precise new CRISPR tool could tackle a plethora of genetic diseases
 CRISPR hacks enable pinpoint repairs to genome
CRISPR hacks enable pinpoint repairs to genome