- NEWS AND VIEWS
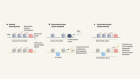
Protein complexes assemble as they are being made
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 561, 186-187 (2018)
doi: https://doi.org/10.1038/d41586-018-05905-4
References
Shiber A. et al. Nature 561, 268–272 (2018).
Ingolia, N. T., Ghaemmaghami, S., Newman, J. R. S. & Weissman, J. S. Science 324, 218–223 (2009).
Becker, A. H., Oh, E., Weissman, J. S., Kramer, G. & Bukau, B. Nature Protocols 8, 2212–2239 (2013).
Halbach, A. et al. EMBO J. 28, 2959–2970 (2009).
Duncan, C. D. S. & Mata, J. PLoS Genet. 7, e1002398 (2011).
Ellis, R. J. Adv. Exp. Med. Biol. 594, 1–13 (2007).
Liu, F., Jones, D. K., de Lange, W. J. & Robertson, G. A. Proc. Natl Acad. Sci. USA 113, 4859–4864 (2016).
Sugimoto, Y. et al. Nature 519, 491–494 (2015).
Yasuda, K. et al. J. Cell Biol. 203, 737–746 (2013).
Berkovits, B. D. & Mayr, C. Nature 522, 363–367 (2015).
Chartron, J. W., Hunt, K. C. L. & Frydman, J. Nature 536, 224–228 (2016).
Shieh, Y.-W. et al. Science 350, 678–680 (2015).
Wells, J. N., Bergendahl, L. T. & Marsh, J. A. Cell Rep. 14, 679–685 (2016).
Mayr, C. Annu. Rev. Genet. 51, 171–194 (2017).
 Read the paper: Cotranslational assembly of protein complexes in eukaryotes revealed by ribosome profiling
Read the paper: Cotranslational assembly of protein complexes in eukaryotes revealed by ribosome profiling
 Mature proteins braced by a chaperone
Mature proteins braced by a chaperone
 Enzymes can adapt to cold by wiggling regions far from their active site
Enzymes can adapt to cold by wiggling regions far from their active site