- TECHNOLOGY FEATURE
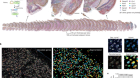
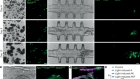
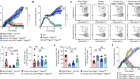
Single-cell approaches to immune profiling
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 557, 595-597 (2018)
doi: https://doi.org/10.1038/d41586-018-05214-w
References
Bandura, D. R. et al. Anal. Chem. 81, 6813–6822 (2009).
Bendall, S. C. et al. Science 332, 687–696 (2011).
Korin, B., Dubovik, T. & Rolls, A. Nature Protoc. 13, 377–391 (2018).
Korin, B. et al. Nature Neurosci. 20, 1300–1309 (2017).
Cheung, P. et al. Cell https://doi.org/10.1016/j.cell.2018.03.079 (2018).
Shahi, P., Kim, S. C., Haliburton, J. R., Gartner, Z. J. & Abate, A. R. Sci. Rep. 7, 44447 (2017).
Klein, A. M. et al. Cell 161, 1187–1201 (2015).
Macosko, E. Z. et al. Cell 161, 1202–1214 (2015).
Stephenson, W. et al. Nature Commun. 9, 791 (2018).
Gierahn, T. M. et al. Nature Meth. 14, 395–398 (2017).
Peterson, V. M. et al. Nature Biotechnol. 35, 936–939 (2017).
Stoeckius, M. et al. Nature Meth. 14, 865–868 (2017).
 How to build a human cell atlas
How to build a human cell atlas
 Single-cell sequencing made simple
Single-cell sequencing made simple