
- NEWS AND VIEWS
Forests in flux as climate varies
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 556, 35-37 (2018)
doi: https://doi.org/10.1038/d41586-018-02858-6
References
Parmesan, C. & Yohe, G. Nature 421, 37–42 (2003).
Zhang, T., Niinemets, Ü., Sheffield, J. & Lichstein, J. W. Nature 556, 99–102 (2018).
Kunstler, G. et al. Nature 529, 204–207 (2016).
Bechtold, W. A. & Patterson, P. L. The Enhanced Forest Inventory and Analysis Program — National Sampling Design and Estimation Procedures. Gen. Tech. Rep. SRS–80 (US Department of Agriculture Forest Service, 2005).
McDowell, N. G. & Allen, C. D. Nature Clim. Change 5, 669–672 (2015)
Ruiz-Benito, P. et al. Glob. Change Biol. 23, 4162–4176 (2017).
Smith, S. E. & Read, D. J. Mycorrhizal Symbiosis (Elsevier, 1997).
Phillips, R. P., Brzostek, E. & Midgley, M. G. New Phytol. 199, 41–51 (2013).
Soudzilovskaia, N. A. et al. Glob. Ecol. Biogeogr. 24, 371–382 (2015).
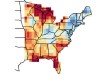 Read the paper: Shifts in tree functional composition amplify the response of forest biomass to climate
Read the paper: Shifts in tree functional composition amplify the response of forest biomass to climate
 Drought in the Congo Basin
Drought in the Congo Basin
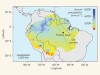 The resilience of Amazonian forests
The resilience of Amazonian forests




