
Abstract
Regulatory T cells (Tregs) and CD4+ T helper (Th) cells have important roles in bile duct injury of biliary atresia (BA). However, their impacts on liver fibrosis are undefined. Between 2013 and 2016, 146 patients with various stages of BA were enrolled in this study. Peripheral blood, liver biopsy and lymph node samples were collected. Flow cytometry, magnetic cell sorting and immunostaining were used to characterize lymphocytes from BA patients. Deficiency of Tregs was observed along with increased Th1, Th2 and Th17 frequencies in the peripheral blood and livers of BA patients. The levels of peripheral and intrahepatic Th1 cells positively correlated with the stage of liver fibrosis. Furthermore, Th1 cells were located in close proximity to activated hepatic stellate cells (HSCs) and areas of fibrosis in BA livers. In culture, Th1 cells accelerated the proliferation and secretion of profibrogenic markers of HSCs through the IFN-γ/STAT1 pathway. Of note, Tregs blocked the Th1-stimulated effects on HSCs by inhibiting Th1-induced activation of STAT1. Consistent with the results of in vitro study, intrahepatic IFN-γ/STAT1 levels increased in relation to the severity of liver fibrosis in BA patients, and the altered balance between MMP2 and TIMP1 expressions in livers may contribute to increased deposition of extracellular matrix and fibrosis. Finally, to identify the effects of Th1 cells on Tregs, we demonstrated that Th1 cells upregulated the proportion of aTreg cells by secreting IFN-γ cytokine. Thus, aberrant Th1 immune responses in BA promote the proliferation and secretion of HSCs through the IFN-γ/STAT1 pathway. The regulation of HSCs by the interactions between Tregs and Th1 cells might be part of the mechanism underlying progressive liver fibrosis and may be a suitable target for therapy.
Similar content being viewed by others
Main
Biliary atresia (BA) is a rare pediatric liver disease characterized by progressive inflammation and fibrosis of both intrahepatic and extrahepatic bile ducts.1, 2 The etiology of BA is largely unknown. A current view of the pathogenesis of BA suggests the involvement of both primary perinatal hepatobiliary virus infection and secondary generation of exaggerated inflammatory or autoimmune-mediated bile duct injury.3, 4 One potential mechanism to explain the abnormal inflammatory and immune response is the loss of functional regulatory T cells (Tregs), defined by the expression of the surface markers CD4 and CD25 and the transcription factor FoxP3, which are indispensable for the maintenance of self tolerance and immune homeostasis.5, 6 Dysfunction of Treg has been associated with an abnormal increase in CD4+ T helper (Th) cell numbers, which has been reported to have critical roles in bile duct injury of BA.5, 6,7, 8 CD4+ Th cells can be subdivided into three different types based on their distinctive patterns of cytokine secretion: IFN-γ-secreting CD4+ T cells (Th1), IL-4-secreting CD4+ T cells (Th2) and IL-17- producing CD4+ T cells (Th17).6 In addition, human FoxP3+CD4+ T cells can be divided into three phenotypically and functionally distinct subpopulations based on their CD45RA and FoxP3 expression levels: CD45RA+FoxP3lo resting Treg cells (rTregs) and CD45RA−FoxP3hi activated Treg cells (aTregs) that manifest suppressive phenotype in vitro, and cytokine-secreting CD45RA−FoxP3lo nonsuppressive T cells.9 The classification of the FoxP3+ T cell into three different cell subsets allowed experimental analyses aimed at understanding Tregs differentiation dynamics, involvement in disease states and the regulation of immune responses through manipulating particular subsets.
Because of the progressive nature of BA, up to 80% of patients eventually require liver transplantation. For reasons not clear, liver fibrosis continues to progress even after adequate bile flow is restored through a surgical Kasai portoenterostomy intervention.10, 11, 12, 13 Mechanisms leading to liver fibrosis are largely unknown, but accumulating evidence suggests that immune cells are critical in modulating fibrogenesis.14, 15 For example, in chronic hepatitis C, Tregs have acquired differentiation as regulators of fibrogenesis in addition to suppressing local immune responses.16 Also, it has been reported that Tregs can interfere with the regulation of fibrogenesis by modifying the functions of NK cells.17 Tregs and Th cells are key members of the immune system. Over the past decade, studies on the role of Tregs or Th cells in BA have focused on inflammatory or immune-mediated bile duct injury.18, 19 However, the impact of Tregs or Th cells on liver fibrosis, and whether interactions between Tregs and Th cells affect the regulation of hepatic fibrosis in BA have not been adequately studied. In this study, we analyzed the dynamics of Tregs and Th cells in various stages of BA to determine whether dysfunction of Tregs and aberrant Th responses persist over time. We also sought to determine the impact of these cells on progressive liver fibrosis in BA patients.
Results
Persistent deficiency of Tregs combined with aberrant Th1, Th2 and Th17 frequencies in BA patients
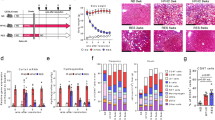
A decreased frequency of Tregs has been reported in BA.17, 20 However, it is unclear, whether deficiency of Tregs is persistent over the duration of disease and Treg function is altered.20 To examine the dynamics of Tregs in BA, we analyzed the surface expression of CD4 and CD25 and the intracellular expression of FoxP3 transcription factor to determine the proportion of Tregs in peripheral blood from BA patients at various stages of disease. A deficiency of Tregs was detected in both early-stage (average 4.14±1.03%) and late-stage BA (3.40±1.01%) groups compared with the controls (5.18±0.53%; Figure 1a) and the controls (5.40±0.50%) as reported;20 however, 2 weeks after LT, the percentage of Tregs in peripheral blood mononuclear cells (PBMCs) of BA patients at the late stage of disease increased compared with preoperative values (4.44±1.49% versus 3.40±1.02%; P<0.05; Figure 1a).

Proportions of Tregs, Th1, Th2 and Th17 cells in BA patients at various stages and their correlations with liver fibrosis. (a) Flow cytometry of PBMCs gated on CD4+ T cells. Numbers circled in red indicate percentages of CD25+FoxP3+ Treg in CD4+ T cells. The results of representative patients are shown in the first four panels, and the compiled results from all patients are shown in the last panel. Horizontal bars indicate mean±S.D. (b) Flow cytometry of PBMCs gated on CD4+ T cells. Left panels: detection of IFN-γ-producing (Th1) or IL-4-producing (Th2) cells in representative patients (top row); detection of Th1 or IL-17-producing (Th17) cells in representative patients (bottom row). Numbers indicate percentages of Th1 (red circle), Th2 (blue circle) and Th17 (purple circle) cells. Right panels: compiled results from all patients. Horizontal bars indicate mean±S.D. *P<0.05; **P<0.01. (c) Pearson correlation was performed between proportions of peripheral Th1, Th2, Th17 or Tregs in CD4+ T cells and the stage of liver fibrosis in 56 early-stage BA patients
Next, we examined the percentage of Th1, Th2 and Th17 cells in peripheral blood by flow cytometry. The results showed that there was an increased proportion in both early-stage and late-stage BA groups compared with control groups of Th1 cells (early stage: 15.73%±14.62, late stage: 10.78±8.50%, control: 1.39±0.93%), Th2 cells (early stage: 2.11%±1.39, late stage: 2.39±1.52%, control: 0.78±0.51%) and Th17 cells (early stage: 0.43%±0.47, late stage: 0.62±0.75%, control: 0.11±0.07%; Figure 1b). The increased frequencies in late-stage BA patients declined after LT in Th1 cells (6.97±3.34% versus 10.78±8.49%), Th2 cells (1.80±1.19% versus 2.39±1.52%) and Th17 cells (0.21±0.13% versus 0.62±0.75%; Figure 1b).These results demonstrate that BA patients manifest persistent deficiency of Tregs and increased Th1, Th2 and Th17 frequencies in peripheral blood until after LT.
For decades, studies have focused mainly on the effects of Th cells and Tregs in bile duct injury of BA.21, 22 However, the role of Th cells and Tregs in progressive liver fibrosis has remained undefined. To examine the involvement of T-cell subsets in hepatic fibrosis, we performed Pearson correlation between the proportions of peripheral Th1, Th2, Th17 or Tregs and histological stage of liver fibrosis in 56 early-stage BA patients. The results showed that the proportion of Th1 cells, but not Tregs, Th2 or Th17 cells, was positively correlated with the stage of liver fibrosis (r=0.422; P=0.001) in BA (Figure 1c).
The proportions of Tregs and Th cells in BA livers compare with those in lymph nodes and PBMCs
We next analyzed Tregs and Th cells in livers and porta hepatis lymph nodes from 24 early-stage BA patients, and compared with the frequencies in PBMCs from matched BA patients. The results showed that the percentage of Tregs in BA livers (2.31±0.74%) was lower than that in PBMCs (3.93±1.20%) and lymph nodes (7.30±1.86%; Figures 2a and c). Interestingly, an apparently higher frequency of Th1 cells was observed in BA livers (30.26±21.48%) than in BA lymph nodes and PBMCs (9.91±6.94% and 16.04±15.81%). There was also an increased proportion of Th2 and Th17 cells in BA livers compared with corresponding values in BA lymph nodes and PBMCs (Figures 2b and c). Subset analyses showed that the increased proportions of Tregs from PBMCs to lymph nodes, with the lowest level in livers was associated with a reverse pattern of decreased Th cells from PBMCs to lymph nodes, with the highest level in livers (Figure 2c, third row). Pearson correlation was performed between the proportions of intrahepatic Th1, Th2, Th17 or Tregs and histological stage of liver fibrosis in 24 early-stage BA patients. The results revealed that the frequency of intrahepatic Th1 cells, but not intrahepatic Tregs, Th2 or Th17 cells, was positively correlated with the stage of liver fibrosis (r=0.752; P<0.001; Figure 2d, first panel). Therefore, these results demonstrate that deficiency of Tregs is associated with increased Th1, Th2 and Th17 frequencies in BA livers. The accumulated Th1 cells in livers may be related with the progressive liver fibrosis of BA.

Comparison of the proportions of Tregs and Th cells in BA livers, lymph nodes and PBMCs. (a) Expression of CD25+FoxP3+ Treg in CD4+ T cells. (b) Detection of Th1 or Th2 cells in representative patients (top row); detection of Th1 or Th17 cells in representative patients (bottom row). Numbers indicate percentages of Th1 (red circle), Th2 (blue circle) and Th17 (purple circle) cells. (c) First and second rows: compiled results from 24 early-stage BA patients. Horizontal bars indicate mean±S.D. Third row: changing trends of Tregs (left panel) and Th cells (right panel). *P<0.05; **P<0.01. (d) Pearson correlation was performed between proportions of intrahepatic Th1, Th2, Th17 or Tregs in CD4+ T cells and the stage of liver fibrosis in 24 early-stage BA patients
Intrahepatic T-bet+Th1 cell numbers correlate with the stage of liver fibrosis in BA
On the basis of these findings, we then analyzed successive liver sections for Th1 cells by immunohistochemical and multiple immunofluorescence staining (Figure 3). The grades of liver fibrosis were classified as I, II, III and IV according to Masson's trichrome staining (Figure 3a). Immunohistochemical staining for T-bet, which specifically identifies Th1 cells, showed that intrahepatic T-bet+ Th1 cell populations increased over the progression of liver fibrosis. Furthermore, T-bet+ Th1 cells were located in close proximity to areas of fibrosis (Figure 3b). To clarify the spatial relationships between HSCs and Th1 cells, we performed immunofluorescence staining for alpha-SMA+ HSCs and T-bet+ Th1 cells. We found that alpha-SMA+ HSCs were surrounded closely by multiple T-bet+ Th1 cells (Figure 3c). Next, we quantified the number of T-bet+ Th1 cells (median 10 cells, range 2–72, per 200 × 200 μm visual field). Pearson correlation analysis confirmed that the histological stage of liver fibrosis positively correlated with the number of intrahepatic T-bet+ Th1 cells (r=0.685; P<0.001; Supplementary Figure 1).

Intrahepatic Th1 cells were linked to the progress of liver fibrosis in BA. (a) Successive liver sections were used for Masson's trichrome staining to assess the stage of liver fibrosis. Data shown are representative of 32 early-stage BA patients with liver fibrosis grades I to IV. (b) Immumohistochemical staining was performed to detect T-bet+ cells, which specifically identifies Th1 cells.Black arrows indicate T-bet+Th1 cells. (c) Immunofluorescence staining for alpha-SMA+HSCs (red) and T-bet+Th1 cells (green) identified HSCs surrounded by Th1 cells. White arrows mark the positions of T-bet+Th1 cells
Th1 cells accelerate the proliferation and secretion of HSCs through the IFN-γ/STAT1 pathway and this effect is attenuated by Tregs
Then, to evaluate the function of Th1 cells on HSCs, we used a co-culture system. Under normal physiological conditions, the activities of matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs) are precisely regulated. A loss of activity control may result in diseases such as fibrosis. MMP2 is one of the most important members of the MMP family because it preferentially cleaves denatured collagens (gelatin), laminin and collagens. TIMP1 can both promote cell proliferation and inhibit the proteolytic activity of MMPs.23,24 Upregulation of profibrogenic genes MMP2, TIMP1 and collagen I was used as an indicator of the activation state of HSCs.16 The proliferation of HSCs and expression of profibrogenic genes in HSC supernatants were tested in our study.
Both Th1 supernatants and Th1 cells accelerated the proliferation of HSCs (Figures 4a and b) and increased the levels of MMP2, TIMP1 and collagen I in HSC supernatants (Figures 5a–d). Increased HSCs proliferation and collagen I secretion directly contribute to deposition of extracellular matrix proteins and scarring.25 As fibrosis is also associated with altered patterns of matrix degradation, the regulation of MMP2 and TIMP1 by Th1 cells might therefore provide another mechanism for fibrosis. Our result showed that although there was a substantial increase in both MMP2 and TIMP1 levels in Th1-stimulated groups, the ratio of MMP2 and TIMP1 expressions decreased in Th1-stimulated groups compared with control group (Supplementary Figure 2). Next, to explore the potential pathways specifically mediating HSCs proliferation and activation, Th1 cells were pretreated with neutralizing antibodies against candidate cytokines. As shown in Supplementary Figure 3, blocking of IFN-γ cytokine inhibited Th1-induced effects on HSCs, whereas anti-IL-2 and anti-TNF-α had negligible effects. Given that STAT1 is a downstream effector of IFN-γ pathway, we applied short interfering RNAs (siRNA), specifically targeting STAT1 (siSTAT1) or IFN-γR (siIFN-rR) expression, to investigate their functions in HSCs. The changes of STAT1 and phosphorylation STAT1 (p-STAT1) in this pathway were also analyzed. The results revealed that both Th1 supernatants and Th1 cells upregulated STAT1 activity in HSCs (Figure 5a, bottom panel and Figure 5c, right panel). The depletion of either STAT1 or IFN-γR significantly inhibited the Th1-induced proliferation of HSCs (Figure 4a and Figure 4b, left panel) and downregulated both basal and Th1-induced secretion of profibrogenic markers (Figure 5a, top panel and Figure 5c, left panel), a result which demonstrated that IFN-γ/STAT1 activation was involved in the proliferative and secretion ability of HSCs.

Th1 cells accelerate the proliferation of HSCs through the IFN-γ/STAT1 pathway and this impact is attenuated by Tregs. (a) HSCs were stimulated with supernatants or cells of Th1 or Treg from BA for 24 h, with or without pretreatment of IFN-γR siRNA or STAT1 siRNA for 12 h. NC, normal control; Th1+Treg supernatants, supernatants from Th1 cells prestimulated with Tregs; Th1+Treg cells, Th1 cells prestimulated with Tregs. (b) Quantification of cell proliferative assay at 24 and 48 h. Values are means±S.D. of 12 independent experiments assayed in duplicate. (*P<0.05; **P<0.01). (c) Left panel: HSCs were transfected with its specific siRNAs (siSTAT1 or siIFN-γR) following treatment with various concentrations of rIFN-γ for 24 h. Right panel: quantification of cell proliferative assay at 24 h

Expression of profibrogenic markers TIMP1, MMP2 and collagen I in HSC supernatants. (a and b) HSCs were treated as described in Figure 4a. Top panel: the level of profibrogenic markers TIMP1, MMP2 and collagen I in HSC supernatants was assessed by western blot analyses. Bottom panel: HSC lysates were assessed for STAT1, p-STAT1 and β-tubulin expression. (c and d) Quantification of western blot results shown in (a orb). Values represent means±S.D. of 12 independent experiments (*P<0.05; **P<0.01). (e) Detection of profibrogenic markers TIMP1, MMP2 and collagen I in HSC supernatants after rIFN-γ treatment for 24 h. (f) Quantification of western blot results shown in (e) (*P<0.05; **P<0.01)
As Treg is an important regulator cell, we wonder whether Tregs regulate the Th1-mediated effects on HSCs by inhibiting IFN-γ production. We prestimulated Th1 cells with Tregs over 24 h, and added mixed supernatants or cells to HSCs. The results showed that Tregs blocked the Th1-stimulated proliferation (Figures 4a and b, right panel) and secretion of profibrogenic markers of HSCs (Figure 5b, top panel and Figure 5d, left panel), by inhibiting Th1-induced upregulation of STAT1 activity in HSCs (Figure 5b, bottom panel and Figure 5d, right panel). Then, HSCs were stimulated with various concentrations of rIFN-γ. As shown in Figure 4c and Figures 5e and f, rIFN-γ promoted the proliferation and secretion of profibrogenic markers of HSCs in a dose-dependent manner. Furthermore, siSTAT1 or siIFN-γR treatment inhibited the rIFN-γ-mediated pro-proliferation and secretion effects on the HSCs.
Taken together, these data suggest that Th1 cells from BA patients accelerate the proliferation and secretion of profibrogenic markers of HSCs through the IFN-γ/STAT1 pathway. Of note, Tregs blocked the Th1-stimulated effects on HSCs. The regulation of HSCs by the interactions of Tregs and Th1 cells might therefore provide a mechanism for progressive liver fibrosis.
Intrahepatic IFN-γ/p-STAT1 levels increased over the progression of liver fibrosis in BA
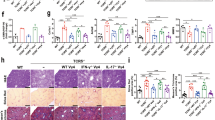
The above in vitro study demonstrated that Th1 cells acted on HSCs through the IFN-γ/STAT1 pathway. We next investigated the IFN-γ/STAT1 pathway in BA livers. Liver tissues were staged by Masson staining as described above. We further divided patients into two groups: mild liver fibrosis (fibrosis grades I and II) and severe liver fibrosis (fibrosis grades III and IV). We tested the expressions of collagen I, TIMP1, MMP2 and components of IFN-γ/STAT1 pathway (IFN-γ and p-STAT1) in the liver specimens of 32 early-stage BA patients based on the immunohistochemical assay. Figure 6a showed that both IFN-γ andp-STAT1 protein expressions were detectable in mild and severe groups of livers. Furthermore, the expressions of IFN-γ protein were elevated in severe liver fibrosis compared with those in mild liver fibrosis. Immunohistochemical nuclear staining for p-STAT1, revealed that intrahepaticp-STAT1+ cell populations were significantly increased in severe liver fibrosis compared with those in mild liver fibrosis, and p-STAT1+ cells were located in close proximity to areas of fibrosis. As for profibrogenic markers staining, Collagen I, TIMP1 and MMP2 protein levels were higher in severe liver fibrosis than in mild liver fibrosis. In addition, the expression of TIMP1 in severe liver fibrosis increased by almost threefold compared with expressions in mild liver fibrosis (P<0.01), whereas the expression of MMP2 was only slightly increased in severe liver fibrosis, and this change did not reach a statistically significant level (Figure 6b). Thus, consistent with in vitro study, the intrahepatic IFN-γ/STAT1 levels increased in relation to the degree of progression of liver fibrosis in BA, and the ratio of TIMP1 and MMP2 expressions increased in severe liver fibrosis than in mild liver fibrosis.

Immunohistochemical staining of liver sections from BA patients. (a) Representative images of IFN-γ, p-STAT1, MMP2, TIMP1 and collagen I staining in liver sections of BA patients. (b) Quantification of the results showed in a (*P<0.05; **P<0.01)
Inhibitory capacity of Tregs from BA correlates with their CTLA-4 expression
Given that Tregs interfere with the regulation of fibrogenesis by Th1 cells, as we previously demonstrated, we wish to identify the interactions between Tregs and Th1 cells. First, we examined whether Tregs, isolated from BA patients induced the same immunosuppressive changes in Th1 cells as Tregs from controls. Supplementary Figure 4 demonstrated that Tregs from BA patients inhibit cytokine secretion by Th cells to a greater extent than Tregs from controls. Then, to identify the mechanism for Treg-mediated suppression of Th1 cells, Supplementary Figure 5 demonstrated that transfer of Tregs suppressed Th1 cells in PBMC cultures; pretreatment of Tregs with neutralizing anti-CTLA-4 antibody reversed this effect on Th1 cells, while the inhibitory effects of Tregs on Th1 cells were only slightly reversed upon addition of anti-IL-8, anti-TGF-β1 and anti-IL-10. Furthermore, the CTLA-4 expression of Tregs from BA patients correlated with their ability to inhibit cytokines secretion by Th cells (Figure 7a and Supplementary Figure 6).

Treg subsets in BA patients at different stages. (a) Pearson correlation was performed between the CTLA-4 expression of Tregs from 10 early-stage BA patients and their ability to inhibit cytokine secretion by Th1, Th2 and Th17 cells. (b) Flow cytometry of PBMCs gated on CD4+ T cells. Based on the expression of FoxP3 and CD45RA, human FoxP3+CD4+ T cells were separated into three different subpopulations. Treg group I: CD45RA+FoxP3lo resting Treg cells (rTregs); Treg group II: CD45RA-FoxP3hi activated Treg cells (aTregs); Treg group III: cytokine-secreting CD45RA-FoxP3lo non-Treg cells. (c) Quantification of results from (b). Columns represent means±S.D. (*P<0.05; **P<0.01). (d) Expression of CTLA-4 by each Treg subset from BA patient. (e) Left panel: detection of Treg subsets in PBMCs; right panel: quantification of results from left panel. Columns represent means±S.D. (*P<0.05)
As for Treg subsets analysis, the results revealed that a deficiency of rTregs was detected in both early-stage and late-stage BA groups compared with controls (Figures 7b and c, Treg group I). In contrast, there was an obvious increase in the proportion of aTregs in all BA groups (Figures 7b and c, Treg group II). Then, the CTLA-4 expression was assessed for each subset, showing that aTregs highly expressed CTLA-4, whereas rTregs only weakly expressed this molecule (Figure 7d). Because CTLA-4 expressions of Tregs correlated with their ability to inhibit cytokines secretion, aTregs with the highest expression of CTLA-4 may be the main effectors for Th cells suppression.
Finally, to determine whether elevated Th1 cells in BA patients could affect the function and differentiation of Tregs, we assessed the proportions of Treg subsets after the PBMCs were stimulated with Th1 supernatants. As illustrated in Figure 7e, incubation with Th1 supernatants for 48 h upregulated the proportion of aTregs, without affecting the proportions of other Treg subsets. Next, to explore the possible pathways mediating Treg differentiation, Th1 cells were pretreated with neutralizing anti-IFN-γ, anti-IL-2 and anti-TNF-α antibodies. Blockage of IFN-γ cytokine apparently inhibited Th1-induced effects on aTregs (Figure 7e), whereas the effects of other neutralizing antibodies were negligible (data not shown). Thus, Th1 cells upregulated the proportion of aTreg cells by secreting IFN-γ cytokine. The role of the IFN-γ signaling pathway in Treg differentiation and function will be investigated in a later study.
Discussion
A decreased frequency of Tregs in peripheral blood has been reported in BA patients.5,20 However, the dynamic of Tregs and Th cells in BA is unclear. In this study, we demonstrated that BA patients manifest persistent deficiency of Tregs and increased Th1, Th2 and Th17 frequencies in the peripheral blood. Furthermore, in contrast to previous studies,20 in which liver tissue was not available for flow cytometry analysis, we used both fresh liver tissue and porta hepatis lymph nodes from BA patients for study of Tregs and Th cells. Subset analyses demonstrated an opposite changing pattern of Tregs and Th cells from BA PBMCs to BA lymph nodes and BA livers. The reason for a relatively lower frequency of Th cells in BA lymph nodes may be that increased Tregs suppressed the aberrant Th-cell function. On the contrary, the lowest proportion of Tregs in BA livers and consequent decreased inhibition for Th cells may account for the highest frequency of Th cells in BA livers (Figures 2a and b).
The impact of Th1 responses on fibrogenesis is still controversial.16,17 Studies have shown that repeated peritoneal inflammation induces Th1 cells to compromise tissue repair by shifting acute inflammation into a more chronic pro-fibrotic state.25 In contrast, other models have highlighted conflicting roles for IFN-γ, described as an antifibrotic regulator that controls collagen synthesis and deposition, in the development of fibrosis.26,27 In our study, key findings are that, in the peripheral blood and intrahepatic environments, Th1 cell numbers positively correlated with the kinetics of liver fibrosis (Figure 1c and Figure 3a), and Th1 cells were enriched in close proximity to alpha-SMA+ HSC and areas of fibrosis in the liver (Figure 2). The positioning of Th1 cell near areas of fibrosis would appear advantageous for a role of Th1 cells in fibrogenesis by acting on HSCs. To identify the impact of Th1 cells on liver fibrosis, Th1 cells or Th1 supernatants from BA patients were co-cultured with HSCs, leading to a novel observation that Th1 cells from BA patients promoted the proliferation and secretion of profibrogenic markers of HSCs through the IFN-γ/STAT1 pathway. Furthermore, Tregs blocked the Th1-induced proliferation and secretion of profibrogenic markers of HSCs by inhibiting Th1-induced upregulation of STAT1 activity (Figures 4 and 5). Consistent with the results of in vitro study, immunostaining revealed that intrahepatic IFN-γ/STAT1 levels increased in relation to the severity of liver fibrosis in BA, and decreased MMP2/TIMP1 ratio in severe liver fibrosis may contribute to increased matrix deposition and fibrosis. These data demonstrated that Th1 cells may accelerate the proliferation and secretion of HSCs by targeting IFN-γ/STAT1 signaling in BA livers. Our studies identified Th1 cells as an intrahepatic source of IFN-γ in BA mediate the fibrogenic response through interactions with HSCs. Despite the previously reported profibrogenic effect of Tregs,16 depletion of Tregs and consequent decreased inhibition in BA livers is likely to contribute to the persistence of activated Th1 cells, resulting in enhanced pro-fibrotic activity.
Given that Tregs in BA livers interfered with the regulation of fibrogenesis by Th1 cells, we then explored the mechanisms in inhibition of Th1 cells by Tregs. Our results demonstrated that Tregs can profoundly inhibit Th1 cells function in PBMCs by a CTLA-4-dependent mechanism. This could be explained by CTLA-4 transmission of inhibitory signals to CD80/CD86-positive antigen presenting cells resulting in induction of indoleamine 2,3-dioxygenase (IDO) activity.28,29 Indoleamine 2,3-dioxygenase catabolizes the amino acid tryptophan, leading to localized tryptophan depletion and the consequent inhibition of T cells.30 Because CTLA-4 expressions of Tregs correlated with their ability to inhibit cytokines secretion, as for Treg subsets, aTregs with the highest expression of CTLA-4 may be the main effectors for Th-cell suppression. Previous studies have identified the effects of Tregs on Th1 cells, while the effects of Th1 cells on Tregs have not been reported. In our study, we demonstrated that Th1 cells upregulated the proportion of aTreg cells by secreting IFN-γ cytokine. Elevated Th1 cells in BA patients may account for the high prevalence of aTregs, which may be generated in an attempt to compensate for the decreased proportion of rTregs.
In summary, deficiencies of Tregs result in aberrant Th1, Th2 and Th17 responses in the peripheral blood and livers of BA patients. The levels of both peripheral and intrahepatic Th1 cells positively correlate with the stage of liver fibrosis. Th1 cells accelerate the proliferation and secretion of profibrogenic markers of HSCs through the IFN-γ/STAT1 pathway in BA. Of note, the Th1-induced effects on HSC were attenuated by Tregs. The regulation of HSCs by the interactions between Tregs and Th1 cells might be part of the mechanism underlying progressive liver fibrosis and may be a suitable target for therapy.
Materials and methods
Human subjects
Between 2013 and 2016, 146 patients with various stages of BA were enrolled in this study. PBMCs were collected from 128 early-stage BA patients (ages 1 month to 4 months) at the time of Kasai portoenterostomy in Xin Hua Hospital, 18 late-stage BA patients (ages 6 months to 10 months) admitted to Ren Ji hospital for liver transplantation (LT) and 20 age-matched control patients (ages 3 months to 11 months). Eighteen late-stage patients were followed for up to 2 weeks after LT. In addition, both porta hepatis lymph nodes and liver wedge biopsy samples were obtained at the time of the surgery from 24 early-stage BA patients. PBMCs (from 56 early-stage BA, 18 late-stage BA and 13 control patients) and immune cells (from 24 early-stage BA livers and lymph nodes) were analyzed for Tregs and Th cells by flow cytometry. Tregs or Th1 cells were isolated from 72 early-stage BA patients or seven control patients by magnetic cell sorting. This study was approved by the Ethics Committee of the Xin Hua Hospital and the Ren Ji Hospital, Affiliated to Shanghai Jiao Tong University School of Medicine, and written informed consent was obtained from the parents. The clinical characteristics of the patients are presented in Supplementary Tables.
Magnetic cell sorting
Tregs were isolated from PBMCs using the Human CD4+CD25+CD127dim Regulatory T Cell Isolation Kit II (Miltenyi Biotec, Auburn, CA, USA) according to the manufacturer’s instructions. We also depleted Tregs from PBMCs using anti-CD25 microbeads (Miltenyi Biotec). Th1 cells were isolated using the EasySep Human Th1 Cell Isolation Kit (Stem Cell, Vancouver, BC, Canada). The purity of isolated cell populations was >85% as determined by flow cytometric analysis.
Cell culture
Human primary hepatic stellate cells (HSCs) were obtained from ScienCell (San Diego, CA, USA) and were cultured for two to four passages in ‘Stellate Cell Medium’ (SteCM, ScienCell) supplemented with 2% fetal bovine serum, 5 ml stellate cell growth supplement, 10 U/ml penicillin and 10 mg/ml streptomycin (all ingredients obtained from ScienCell) at 37 °C with 5% CO2, and cryopreserved until further use.
Two days before the experiments, HSCs were thawed and 1.5 × 104 HSC/well were seeded in 48-well plates. HSCs were transfected with siRNA duplexes by the use of Lipofectamine RNAiMAX (Carlsbad, CA, USA) according to the manufacturer’s instructions. The cells were transfected with IFN-γR siRNA or STAT1 siRNA for 12 h, then stimulated with Th1 cells or Tregs from BA patients, or various concentrations of recombinant IFN-γ (rIFN-γ) (Prospecbio, Ness Ziona, Israel) for 24 or 48 h. The number of viable cells was determined with a microplate reader using a cell-counting kit-8 (Dojindo, Japan). Isolated Th1 cells were prestimulated with Tregs (1 : 1 ratio) or left unstimulated in RPMI 1640 medium (100 μl) over 24 h. Then, Th1 cells – prestimulated with Tregs or left unstimulated – were incubated with HSCs at a ratio of 1:1, or supernatants (50 μl) from Th1 cells – prestimulated with Tregs or left unstimulated – were added to HSC culture. For blocking experiment, Th1 cells were pretreated with neutralizing anti-IFN-γ, anti-IL-2 or anti-TNF-α antibody (5 μg/ml each; Abcam, San Francisco, CA, USA) for 12 h. Tregs were pretreated with neutralizing anti-IL-8 and anti-TGF-β1 or anti-IL-10 antibody (5 μg/ml each; Abcam) for 12 h.
Statistical analysis
All data were expressed as mean±S.D. Data were evaluated byT-test for two different groups or one-way ANOVA for multiple comparisons. Pearson’s correlation coefficients were calculated for correlation analysis. P-values of P<0.05 were considered statistically significant. Calculations were done with the SPSS package (SPSS 15.0, Chicago, IL, USA) and the GraphPad Prism software package 4.0 (GraphPad Prism, San Diego, CA, USA).
Change history
07 October 2019
"An amendment to this paper has been published and can be accessed via a link at the top of the paper."
References
Hartley JL, Davenport M, Kelly DA . Biliary atresia. Lancet 2009; 374: 1704–1713.
Petersen C, Davenport M . Aetiology of biliary atresia: what is actually known? Orphanet J Rare Dis 2013;8: 128.
Sokol RJ, Mack C, Narkewicz MR, Karrer FM . Pathogenesis and outcome of biliary atresia: current concepts. J Pediatr Gastroenterol Nutr 2003; 37: 4–21.
Selmi C, Vergani D, Mieli-Vergani G . Viruses and autoantibodies in biliary atresia. Gastroenterology 2010;139: 1461–1464.
Tucker RM, Feldman AG, Fenner EK, Mack CL . Regulatory T cells inhibit Th1 cell-mediated bile duct injury in murine biliary atresia. J Hepatol 2013; 59: 790–796.
Mack CL, Tucker RM, Sokol RJ, Kotzin BL . Armed CD4+ Th1 effector cells and activated macrophages participate in bile duct injury in murine biliary atresia. Clin Immunol 2005; 115: 200–209.
Mack CL, Tucker RM, Lu BR, Sokol RJ, Fontenot AP, Ueno Y et al. Cellular and humoral autoimmunity directed at bile duct epithelia in murine biliary atresia. Hepatology 2006; 44: 1231–1239.
Klemann C, Schroder A, Dreier A, Mohn N, Dippel S, Winterberg Tet al. Interleukin 17, produced by gammadelta T cells, contributes to hepatic inflammation in a mouse model of biliary atresia and is increased in livers of patients. Gastroenterology 2016; 150: 229–241 e225.
Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor.Immunity 2009; 30: 899–911.
Haafiz AB . Liver fibrosis in biliary atresia. Expert Rev Gastroenterol Hepatol 2010; 4: 335–343.
Kyoden Y, Tamura S, Sugawara Y, Yamashiki N, Matsui Y, Togashi Jet al. Outcome of living donor liver transplantation for post-Kasai biliary atresia in adults. Liver Transpl 2008; 14: 186–192.
Karakayali H, Sevmis S, Ozcelik U, Ozcay F, Moray G, Torgay Aet al. Liver transplantation for biliary atresia. Transplant Proc 2008; 40: 231–233.
Harpavat S, Garcia-Prats JA, Shneider BL . Newborn bilirubin screening for biliary atresia. N Engl J Med 2016; 375: 605–606.
Guidotti LG, Chisari FV . Immunobiology and pathogenesis of viral hepatitis. Annu Rev Pathol 2006; 1: 23–61.
Spengler U, Nattermann J . Immunopathogenesis in hepatitis C virus cirrhosis. Clin Sci (Lond) 2007; 112: 141–155.
Langhans B, Kramer B, Louis M, Nischalke HD, Huneburg R, Staratschek-Jox A et al. Intrahepatic IL-8 producing Foxp3(+)CD4(+) regulatory T cells and fibrogenesis in chronic hepatitis C. J Hepatol 2013; 59: 229–235.
Langhans B, Alwan AW, Kramer B, Glassner A, Lutz P, Strassburg CPet al. Regulatory CD4+ T cells modulate the interaction between NK cells and hepatic stellate cells by acting on either cell type. J Hepatol 2015; 62: 398–404.
Li J, Bessho K, Shivakumar P, Mourya R, Mohanty SK, Dos Santos JLet al. Th2 signals induce epithelial injury in mice and are compatible with the biliary atresia phenotype. J Clin Invest 2011; 121: 4244–4256.
Shivakumar P, Campbell KM, Sabla GE, Miethke A, Tiao G, McNeal MMet al. Obstruction of extrahepatic bile ducts by lymphocytes is regulated by IFN-gamma in experimental biliary atresia.J Clin Invest 2004; 114: 322–329.
Brindley SM, Lanham AM, Karrer FM, Tucker RM, Fontenot AP, Mack CL . Cytomegalovirus-specific T-cell reactivity in biliary atresia at the time of diagnosis is associated with deficits in regulatory T cells. Hepatology 2012; 55: 1130–1138.
Mack CL, Tucker RM, Sokol RJ, Karrer FM, Kotzin BL, Whitington PFet al. Biliary atresia is associated with CD4+ Th1 cell-mediated portal tract inflammation. Pediatr Res 2004; 56: 79–87.
Shivakumar P, Sabla G, Mohanty S, McNeal M, Ward R, Stringer Ket al. Effector role of neonatal hepatic CD8+ lymphocytes in epithelial injury and autoimmunity in experimental biliary atresia.Gastroenterology 2007; 133: 268–277.
Visse R . Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure, function, and biochemistry. Circ Res 2003; 92: 827–839.
Xiao Y, Wang J, Chen Y, Zhou K, Wen J, Wang Y et al. Up-regulation of miR-200b in biliary atresia patients accelerates proliferation and migration of hepatic stallate cells by activating PI3K/Akt signaling. Cell Signal 2014; 26: 925–932.
Fielding CA, Jones GW, McLoughlin RM, McLeod L, Hammond VJ, Uceda Jet al. Interleukin-6 signaling drives fibrosis in unresolved inflammation. Immunity 2014; 40: 40–50.
Wynn TA . Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat Rev Immunol 2004; 4: 583–594.
Gurujeyalakshmi G, Giri SN . Molecular mechanisms of antifibrotic effect of interferon gamma in bleomycin-mouse model of lung fibrosis: downregulation of TGF-beta and procollagen I and III gene expression. Exp Lung Res 1995; 21: 791–808.
Mellor AL, Chandler P, Baban B, Hansen AM, Marshall B, Pihkala Jet al. Specific subsets of murine dendritic cells acquire potent T cell regulatory functions following CTLA4-mediated induction of indoleamine 2,3 dioxygenase. Int Immunol 2004; 16: 1391–1401.
Walker LS, Sansom DM . The emerging role of CTLA4 as a cell-extrinsic regulator of T cell responses. Nat Rev Immunol 2011; 11: 852–863.
Fallarino F, Grohmann U, Hwang KW, Orabona C, Vacca C, Bianchi Ret al. Modulation of tryptophan catabolism by regulatory T cells. Nat Immunol 2003; 4: 1206–1212.
Acknowledgements
This study is supported by National Natural Science Foundation of China (81200457), National Natural Science Foundation of China (81571468), National Natural Science Foundation of China (81300516), Shanghai Key Laboratory of Pediatric Gastroenterology and Nutrition (14DZ2272400), Shanghai Municipal Commission of Health and Family Planning (2013ZYJB0017), Science and Technology Commission of Shanghai Municipality (14411950400), Science and Technology Funding in Shanghai Jiao Tong University School of Medicine (12XJ10066), Shanghai Municipal Commission of Health and Family Planning (20164Y0176).
Author contributions
The study was conceived and designed by JWe and WC. The experiments were performed by JWe, YZ, WY and JY. The data were analyzed by JWa, JWu, KZ, YX and YW. Contributions of reagents/materials/analysis tools were made by JC and QX. The manuscript was written by JWe and WC.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by Y Shi
Supplementary Information accompanies this paper on Cell Death and Differentiation website
The Editors have retracted this article [1]. After publication concerns were raised with respect to the Western blot data presented in Figure 5a and Figure 5b. The authors have not been able to provide the original, un-cropped, high-resolution images for examination by the Editors; the Editors therefore no longer have confidence in the integrity of these data. Additionally, Figure 4a and Figure 4c contain duplicate images of the siSTAT1 samples. Eleven of the twelve authors agree with this retraction. Qiang Xia did not respond to correspondence about this retraction.
1. Wen J, Zhou Y, Wang J, Chen J, Yan W, Wu J et al. Interactions between Th1 cells and Tregs affect regulation of hepatic fibrosis in biliary atresia through the IFN-?/STAT1 pathway. Cell Death and Differentiation 2017; 24: 997-1006
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Wen, J., Zhou, Y., Wang, J. et al. RETRACTED ARTICLE:Interactions between Th1 cells and Tregs affect regulation of hepatic fibrosis in biliary atresia through the IFN-γ/STAT1 pathway. Cell Death Differ 24, 997–1006 (2017). https://doi.org/10.1038/cdd.2017.31
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cdd.2017.31
This article is cited by
-
Association of T-helper cell cytokine level with age in patients with biliary atresia: a preliminary study
World Journal of Pediatrics (2018)
