
Abstract
Temperature and humidity are among the most important environmental factors affecting insect adaptive strategies and evolution. Here, we report multiple adaptive differences between Drosophila melanogaster isofemale lines derived from the opposite slopes of Lower Nahal Oren canyon at Mount Carmel, Israel. The slopes are separated by 100 m at the bottom and 400 m at the top, and contrast sharply in physical and biotic factors. The multivariate fitness complex analysed in D. melanogaster included oviposition temperature preferences, viability and longevity changes, caused by short-term and lifetime temperature treatments, and resistance to drought stress at different temperatures. Some of these measures were obtained for the sibling species, D. simulans, and gave results that paralleled those of D. melanogaster. We conclude that strong microclimatic natural selection overrides migration in Drosophila at this microsite.
Similar content being viewed by others


Introduction
The opposite slopes of Lower Nahal Oren canyon at Mount Carmel, Israel, designated by us ‘Evolution Canyon’, separated by 100 and 400 m at the bottom and top, respectively, contrast sharply both physically and biotically (Nevo, 1995). The main underlying causal factor for this microsite dissimilarity is the difference in solar radiation. The south-facing slope (SFS) obtains up to 260–300 per cent more solar radiation than the north-facing slope (NFS). Consequently, the SFS is much warmer and drier than the NFS, causing dramatic interslope species diversity and genetic diversity divergence (Nevo, 1995). Temperature and humidity are among the most important environmental factors affecting insect adaptive strategies and evolution. Therefore, Drosophila adaptations to temperature and drought stress have been a popular target system for evolutionary studies for a long time (Ayala, 1965; Coyne et al., 1983; Hoffmann & Parsons, 1991; Huey et al., 1991; James & Partridge, 1995; Neat et al., 1995). Here, we show that Drosophila populations from ‘Evolution Canyon’ display sharply divergent adaptive complexes over a very short geographical distance. It is worth mentioning that stressful environmental conditions cause higher rates of mutation (including insertional mutagenesis) and recombination (Wills, 1984; Zhuchenko & Korol, 1985; Cullis, 1990; Parsons, 1992). Stress-induced changes in the amount and spectrum of genetic variation may serve as an important source of raw material for natural selection to produce novel adaptations (McDonald, 1983; Zhuchenko & Korol, 1983; Carson, 1990; Hoffmann & Parsons, 1991; Korol et al., 1994).
Correlated responses in D. melanogaster have been reported previously in experiments with artificial selection for temperature and/or desiccation resistance (Hoffmann & Parsons, 1989; Rose et al., 1992). Such accumulated differences allow for a subsequent analysis of the genetic basis of responses to artificial selection. On the other hand, biometrical genetic methods applied to analyse adaptive traits in natural populations inhabiting temperate environments usually show relatively low genetic differentiation (Gebhardt & Stearns, 1992; Jenkins & Hoffmann, 1994; Yamamoto, 1994). Here, we report multiple highly significant adaptive differences in D. melanogaster at a microscale, i.e. between genotypes from the close opposite slopes, partly confirmed for its sibling species D. simulans. The revealed strong interslope differentiation could serve as ideal material for unravelling the genetic basis of environmental tolerance in natural populations.
Materials and methods
Materials
Drosophila melanogaster and D. simulans sampling was conducted during 1994 at seven stations across the canyon, three on each slope (nos 1–3 on the SFS and nos 5–7 on the NFS; stations are located at different elevations separated on each slope by only 30 m from each other, at 60, 90 and 120 m above sea level) and one station (no. 4) at the valley bottom. A series of isofemale lines was established from samples collected during the spring–summer period of 1994 and maintained at 25°C.
Preferred oviposition temperature
Forty-two lines of D. melanogaster (six lines from each of the seven stations) and 12 lines of D. simulans from stations 2 and 6 (six lines from each of the two stations) were tested. The test was conducted in a tube (25 cm×17 cm×100 cm) with a temperature gradient from 14°C to 32°C. Vials with fresh medium were placed at five points along the gradient, at 14°C, 19°C, 23°C, 29°C and 32°C, three vials at each point. About 800 4-day-old flies of each line (males and females) were put into the tube for 40 h, and then the vials were transferred to normal conditions (25°C). The proportion of the progeny samples collected from different points along the temperature gradient served as an indicator of the female laying behaviour (temperature preference). The percentage of eclosed imagoes, compared with the pupae number, is a measure of the effect of temperature during early development on the survival rate. The foregoing procedure was repeated six times for each line.
Reaction to heat treatment
Flies (4–5 days old) were placed for 16 h at 25°C in standard vials containing fresh food (20 females and 20 males per vial). Then, after the parental flies were taken out, the vials were subjected to 36°C for 8 h (treatment 1) or to 30°C for the whole subsequent development (treatment 2). The resulting progeny size is an indicator of the viability as affected by the applied treatments (variable V in Table 1). Each ‘treatment×line’ combination was conducted in three independent blocks. The same scheme was used to evaluate how the heat treatment affects the longevity (L=period of survival of 50 per cent of the initial number of flies). This measurement was conducted using 200 mL vials each with 100 flies at the beginning. The flies were periodically transferred onto fresh medium. For each ‘treatment×line’ combination, the results were repeated six times.
Reaction to desiccation treatment
Measurements of resistance to drought stress involved a series of desiccation tests with D. melanogaster isofemale lines from the SFS (station 2) and NFS (station 6), conducted separately on males and females. The resistance to desiccation was estimated by comparing the time to death of 50 per cent of the flies that were subjected to starvation at 0 per cent and 100 per cent humidity; the comparisons were conducted at high (32°C) and normal (25°C) temperatures. Tests for each ‘line×treatment’ combination were repeated six times. Changes in fly weight resulting from desiccation/starvation were also measured in this study. The specific measurements included the weight of fresh flies (4 days old) before starvation and desiccation, Wini; weight of flies that died because of starvation, Wstarv; and weight of dry flies, Wdry. These measurements were repeated five times.
Statistics
Standard statistical tools, multiple linear regression, stepwise linear regression and ANOVA, were used for data treatment. Because of the spatial structure of the collection of flies used to establish the isofemale lines (see Materials and methods), the major ‘explanatory’ variables for the analysis of the observed variation in the adaptive traits were ‘slope’, ‘altitude’ and the interaction ‘slope×altitude’. In addition, the effects of ‘line’ and ‘sex’ were also considered. In the stepwise regression, the F-values for ‘including’ and ‘excluding’ the explanatory variables at each step were 4.0 and 3.9, respectively. In addition to the significance of the regression model, the proportion of between-line variability explained by the above explanatory variables was calculated (Draper & Smith, 1981).
Results
Preferred oviposition temperature
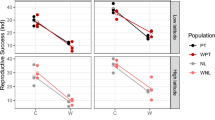
The obtained distribution of the progeny along the temperature gradient in the experimental tube allowed us to calculate the proportions of offspring from vials positioned in the tube at the temperature extremes (28°C+32°C) (Fig. 1a) and (14°C+19°C) (Fig. 1b). Likewise, the mean preferred temperature (Fig. 1c) was calculated for each line using as weights the progeny size obtained at the five temperature points along the gradient in the tube. Using multiple linear regression analysis, we found that, for these three traits, the major explanatory variables were ‘slope’ and ‘slope×altitude’. The proportion (R2) of between-line variability, explained jointly by the variables ‘slope’ and ‘slope×altitude’, was 90.6 per cent, 85.9 per cent and 92.3 per cent for the traits in (Fig. 1a, b and c) respectively. The significance of the regression was P<0.001 in all three cases, with the variance ratio of the variation explained by the regression to the residual variation (F2,39) being, correspondingly, F=188.2, 118.7 and 234.6.

Preferred oviposition temperature in Drosophila melanogaster and D. simulans from the opposite slopes of ‘Evolution Canyon’, Lower Nahal Oren, Mount Carmel: (a) the proportion of offspring from vials positioned at high temperatures; (b) the proportion of offspring from vials positioned at low temperatures; (c) mean preferred temperature. The mean values of individual D. melanogaster lines are marked by dashes, whereas those of D. simulans are marked by triangles. Note the remarkable parallelism between the two species.
The laboratory results of D. melanogaster oviposition preferences clearly show two ecological effects: (i) a significantly higher mean temperature was preferred by females derived from the SFS compared with those from the NFS; and (ii) a general increase in the mean level of preferred laying temperatures with the station altitude (Fig. 1). The latter trend is much stronger for the SFS than for the NFS, reflecting the ‘slope×altitude’ interaction (compare the changes from the bottom station 4 upwards to station 1 on the SFS, and from station 4 upwards to station 7 on the NFS). Significantly higher between-line variation was found on the SFS compared with the NFS: for the mean preferred temperature, the variance ratio was F17,17=8.46 (P<0.001). This effect appeared to be caused by the between-station variation associated with altitude. Notably, the highest preferred temperature was by station 1 flies, the hottest and driest station. Data on other Drosophila species indicate a rather high heritability of the trait ‘temperature preference’ (Yamamoto, 1994), although a direct analysis of the genetic basis should involve mapping of the corresponding behavioural quantitative trait loci (QTL).
A parallel, although on a smaller scale, result was displayed by D. simulans (Fig. 1): significantly higher temperatures were preferred by the SFS females (24.2±0.15°C) compared with the NFS ones (22.9±0.18°C) (Student's t10=5.4, P<0.001). It is interesting to compare the two species with respect to temperature preferences. Based on the data from contrasting stations 2 and 6, higher temperatures are preferred by D. melanogaster only for the lines from station 2. This is explicable by the fact that this station is on the SFS, and D. melanogaster is adapted to higher temperatures than D. simulans (Jenkins & Hoffmann, 1994).
Viability and longevity changes caused by high temperature
The reactions to two treatments have been compared between D. melanogaster lines from stations 2 (SFS) and 6 (NFS): (i) heat shock stress (36°C) for 8 h at the embryo stage, with subsequent development at normal temperature (25°C); (ii) high temperature (30°C) applied during the whole lifetime starting from the embryo stage. In both tests, changes in viability and longevity relative to the control level (all stages at 25°C) were used for interslope comparisons. The standard variance ratio test for nested ANOVA was conducted for the initial traits VT and LT (with T=25°, 30° and 36°C) and for the induced changes of the traits relative to the control levels (V25 and L25) (Table 1). The induced changes were measured by differences, e.g. V25–V30 is the difference between viability measures at 25°C and 30°C. The lower the difference, the higher the stability with regard to temperature treatment. The degrees of freedom for the F-test were 1 and 10 for ‘slope’ and ‘line’, respectively, and 24 for the residual variation. The results indicate that the SFS subpopulation manifests lower fitness than that of the NFS when the entire development proceeds at normal temperature (25°C). This is true for both mean progeny size (V25) and mean longevity (L25).
Heat treatments caused a significant reduction in mean longevity of lines from both slopes. However, SFS lines appeared to be much less sensitive than NFS lines. Likewise, SFS lines showed no reduction in mean viability, in contrast to NFS lines. Especially pronounced is the interslope divergence with respect to their reactions to short-term heat shock treatment of the embryos. In this case, the joint distributions of the variables V25–V36 and L25–L36 for the subpopulations do not overlap even if scores for individual repeats are considered (Fig. 2). Noteworthy is the higher between-subpopulation (localities) variance compared with the between-lines variance (see Table 1 and Fig. 2). This has been reported by other authors at the macrogeographical level (Coyne et al., 1983; Hoffmann & Parsons, 1991).

Reaction of Drosophila melanogaster isofemale lines from the opposite slopes of ‘Evolution Canyon’, Lower Nahal Oren, to short-term heat shock treatment of the embryos. The joint distribution of the induced changes in viability and longevity is represented in the form of a scatter diagram of bivariate scores for individual repeats. The difference between the SFS and NFS subpopulations is so large that even these individual scores manifest distinct, virtually nonoverlapping clusters. The upper and lower clusters represent the NFS (rhombi) and SFS (circles) subpopulations, respectively.
The lower influence of heat treatment on the longevity of the SFS lines than on the NFS lines (see Table 1 and Fig. 2) fits the conclusions reached in artificial selection experiments with D. melanogaster, namely that selection for stress resistance may increase longevity (Rose et al., 1992; Parsons, 1993).
Our results (for both D. melanogaster and D. simulans) clearly indicate that adaptation to the sharply contrasting temperature conditions of the opposite slopes has also resulted in genetic divergence for habitat preferences. The direction of the revealed interslope difference in the oviposition preferred temperatures is exactly that expected in accordance with the average station temperature and habitat selection and habitat choice models of polymorphism maintenance in spatially heterogeneous environments (Hedrick, 1990).
Resistance to desiccation
Treatment of these data used linear regression on dichotomous variables allowing simultaneous evaluation of the direction and the significance of the effects. Clearly, the NFS lines manifest a significant superiority in survival over the SFS lines, when subjected to starvation at conditions in which only one factor of the two, either temperature or humidity, is limiting (lines 1 and 4 in Table 2 This is shown by highly significant ‘slope’ components of the regression model. The interslope difference between the flies reduces at normal conditions (line 3) and vanishes when both stresses are applied (line 2). The most important information concerning the stability of the compared subpopulations comes from the analysis of differences between the survival times at: (i) H=99 per cent and H=0 per cent, when T=25°C or T=32°C (lines 5 and 6); and (ii) T=25°C and T=32°C when H=99 per cent or H=0 per cent (lines 7 and 8). The lower the difference, the higher the stability. Thus, in spite of higher values for survival time manifested by NFS lines under one of the two stresses, heat or drought (lines 1 and 4), the SFS flies manifest a much higher stability when the reaction to desiccation (H=99 per cent−H=0 per cent) is measured under high temperature (line 5) or the reaction to heat treatment (T=25°C−T=32°C) is measured under drought stress (line 8).
Fly weight
We also analysed the changes in fly weight. The results parallel the survival data (see Table 2). In particular, NFS lines are significantly heavier than SFS lines in body weight (line 9), a trait related to general fitness. This interslope weight difference disappears when weight is compared in flies that died because of starvation or when dry weight is compared (lines 10 and 11). Consequently, NFS flies scored higher for the instability parameters than SFS flies (lines 12 and 13). It is noteworthy that the interslope trend of line 9 corroborates the known macrogeographical clines in Drosophila size (Coyne et al., 1983; Neat et al., 1995), i.e. representing the Bergman rule on a microscale. Similar interslope size divergence supporting the Bergman rule was found in landsnails from the opposite slopes of ‘Evolution Canyon’ (Broza & Nevo, 1996) and in the scarab beetle Pygopleurus (Amphicoma) libanonensis (Nevo, 1997).
Discussion
Using desiccation and/or temperature treatments, many authors have found correlations between Drosophila reactions in the laboratory and macrogeographical climatic clinal patterns (Coyne et al., 1983; Hoffmann & Parsons, 1991). Our data concern Drosophila microscale differentiation caused by contrasting temperature and humidity conditions. Strong inter- and intraslope phenotypical and genotypical effects caused by sharp microclimatic divergence have also been found in other organisms at ‘Evolution Canyon’ (Nevo, 1995). The Drosophila adaptive syndrome arising from natural selection at the local scale includes a behavioural trait (oviposition temperature preference), survival and longevity at normal conditions and at temperature, drought and starvation stresses. It should be noted that, before being tested, our lines were raised over 1.5 years in standard laboratory conditions. The corresponding processes of adaptation to laboratory conditions could affect the results obtained only in one direction: reducing the initial genetic differences between flies collected from the opposite slopes. The fact that, in spite of intraslope between-line variation, a 1.5-year maintenance of the material under the same conditions has not abolished the differences in reaction to heat and drought treatments allows us to conclude that the observed dissimilarities reflect the specificity of genetic adaptations to the drastic interslope microclimatic contrasts.
An important future implication of the multivariate adaptive syndrome revealed is that it may be possible to conduct an in-depth genetic analysis using molecular markers. Correlated responses in D. melanogaster have been reported previously in experiments with artificial selection for temperature and/or desiccation resistance (Hoffmann & Parsons, 1989; Rose et al., 1992), which allow for a subsequent analysis of the underlying genetic basis. In contrast, biometrical genetic methods applied to analyse adaptive traits in natural populations inhabiting temperate environments usually show relatively low genetic differentiation (Gebhardt & Stearns, 1992; Yamamoto, 1994). Our material combines the advantages of exhibiting strong differentiation, important for a powerful genetic analysis such as QTL mapping, and being of wild origin. It could provide ideal material for a new QTL mapping strategy dealing with correlated trait complexes (Korol et al., 1994, 1995).
The established significant population genetic divergence between the slopes with respect to adaptive traits is expected but not trivial. Indeed, the distance between the slopes is only 100 m and 400 m at the bottom and top, respectively; hence, a high migration rate cannot be excluded (Baker et al., 1989; Turelli & Hoffmann, 1995). This is even more likely for the intraslope migration. Nevertheless, our data indicate clear divergence between stations, especially strong between the slopes. Remarkably, significant interslope differentiation is manifested by both D. melanogaster and D. simulans. Therefore, to create such an adaptive pattern, selection must be very strong, overriding the migration effect. Significant microscale differentiation (at a few hundreds of metres) was also found in a D. persimilis population at the Sierra Nevada Mountains, California (Taylor & Powell, 1977). In that study, the subpopulations compared were scored for the frequencies of inversion and allozymic variants. The observed differentiation was explained in terms of the habitat choice mechanism.
Theoretical modelling shows that selection against migrants in systems with spatially heterogeneous selection can bring about a genetically determined reduction in the migration rate (Wiener & Feldman, 1993). Clearly, the resulting habitat preferences will help in storing a large amount of genetic polymorphism in the population without a heavy genetic load (Taylor, 1975; Parsons, 1991). In view of that, our parallel data on oviposition temperature preferences in two sibling species of Drosophila from the opposite slopes deserve special attention. These results establish a promising natural system, which allows an in-depth analysis of behavioural adaptation under heterogeneous stressful conditions (Parsons, 1991, 1993).
References
Ayala, F. (1965). Relative fitness of populations of Drosophila serrata and Drosophila birchii. Genetics, 51: 527–544.
Barker, J. S. F., East, P. D. and Christiansen, F. B. (1989). Estimation of migration from perturbation experiments in natural populations. Biol J Linn Soc, 37: 311–334.
Broza, M. and Nevo, E. (1996). Selective land snail predation by the spiny mouse, Acomys cahirinus in Nahal Oren, Mt. Carmel, Israel. Isr J Zool, 40: 143–176.
Carson, H. L. (1990). Increased genetic variance after a population bottleneck. Trends Ecol Evol, 5: 228–230.
Coyne, J. A., Bundgaard, J. and Prout, T. (1983). Geographic variation of tolerance to environmental stress in Drosophila pseudoobscura. Am Nat, 122: 474–488.
Cullis, C. A. (1990). DNA rearrangements in response to environmental stress. Adv Genet, 28: 73–97.
Draper, N. R. and Smith, H. (1981). Applied Regression Analysis, 2nd edn. John Wiley, New York.
Gebhardt, M. D. and Stearns, S. C. (1992). Phenotypic plasticity for life-history traits in Drosophila melanogaster III. Effect of the environment on genetic parameters. Genet Res, 60: 87–101.
Hedrick, P. W. (1990). Genotype-specific habitat selection: a new model and its application. Heredity, 65: 145–149.
Hoffmann, A. A. and Parsons, P. A. (1989). Selection for increased desiccation resistance in Drosophila melanogaster: additive genetic control and correlated responses for other stresses. Genetics, 122: 837–845.
Hoffmann, A. A. and Parsons, P. A. (1991). Evolutionary Genetics and Environmental Stress, Oxford Science Publications, Oxford.
Huey, R. B., Partridge, L. and Fowler, K. (1991). Thermal sensitivity of Drosophila melanogaster responds rapidly to laboratory natural selection. Evolution, 45: 751–756.
James, A. C. and Partridge, L. (1995). Thermal evolution of rate of larval development in Drosophila melanogaster in laboratory and field populations. J Evol Biol, 8: 315–330.
Jenkins, N. L. and Hoffmann, A. A. (1994). Genetic and maternal variation for heat resistance in Drosophila from the field. Genetics, 137: 783–789.
Korol, A. B., Preygel, I. A. and Preygel, S. I. (1994). Recombination Variability and Evolution, Chapman & Hall, London.
Korol, A. B., Ronin, Y. I. and Kirzhner, V. M. (1995). Multitrait analysis in interval mapping of QTL. Genetics, 140: 1137–1147.
Mcdonald, J. F. (1983). The molecular basis of adaptation: a critical review of relevant ideas and observations. Ann Rev Ecol Syst, 14: 77–102.
Neat, F., Fowler, K., French, V. and Partridge, L. (1995). Thermal evolution of growth efficiency in Drosophila melanogaster. Proc R Soc B, 260: 73–78.
Nevo, E. (1995). Asian, African and European biota meet at ‘Evolution Canyon’ Israel: local tests of global biodiversity and genetic diversity patterns. Proc R Soc B, 262: 149–155.
Nevo, E. (1997). Evolution in action across phylogeny caused by microclimatic stresses at ‘Evolution Canyon’. Theor Pop Biol, (in press).
Parsons, P. A. (1991). Evolutionary rates: stress and species boundaries. Ann Rev Ecol Syst, 22: 1–18.
Parsons, P. A. (1992). Fluctuating asymmetry: a biological monitor of environmental and genomic stress. Heredity, 68: 361–364.
Parsons, P. A. (1993). Evolutionary adaptation and stress: energy budgets and habitats preferred. Behav Genet, 23: 231–238.
Rose, M. R., Vu, L. N., Park, S. U. and Graves, J. L. jr (1992). Selection on stress resistance increases longevity in Drosophila melanogaster. Exp Gerontol, 27: 241–250.
Taylor, C. E. (1975). Genetic loads in heterogeneous environments. Genetics, 80: 621–635.
Taylor, C. E. and Powell, J. R. (1977). Microgeographic differentiation of chromosomal and enzyme polymorphisms in Drosophila persimilis. Genetics, 85: 681–695.
Turelli, M. and Hoffmann, A. A. (1995). Cytoplasmic incompatibility in Drosophila simulans: dynamics and parameter estimates from natural populations. Genetics, 140: 1319–1338.
Wiener, P. and Feldman, M. W. (1993). The effect of the mating system on the evolution of migration in a spatially heterogeneous population. Evol Ecol, 7: 251–269.
Wills, C. (1984). The possibility of stress-triggered evolution. In: Mani, G. S. (ed.) Evolutionary Dynamics of Genetic Diversity. Lect Notes Biomath, 53: 299–312.
Yamamoto, A. H. (1994). Temperature preference of Drosophila immigrans and D. virilis: intra- and inter-population genetic variation. Jap J Genet, 69: 67–92.
Zhuchenko, A. A. and Korol, A. B. (1983). Ecological aspects of the recombination problem. Theor Appl Genet, 64: 177–185.
Zhuchenko, A. A. and Korol, A. B. (1985). Recombination in Evolution and Breeding. Nauka Moscow (in Russian).
Acknowledgements
The authors acknowledge with thanks the helpful comments and suggestions of two anonymous referees. This research was supported by the Israeli Ministry of Absorption, the Israeli Ministry of Science, grant no. 4147, the Ancell-Teicher Research Foundation for Genetics and Molecular Evolution and the Israel Discount Bank Chair of Evolutionary Biology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nevo, E., Rashkovetsky, E., Pavlicek, T. et al. A complex adaptive syndrome in Drosophila caused by microclimatic contrasts. Heredity 80, 9–16 (1998). https://doi.org/10.1046/j.1365-2540.1998.00274.x
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1046/j.1365-2540.1998.00274.x
Keywords
This article is cited by
-
Dopamine modulation of sensory processing and adaptive behavior in flies
Cell and Tissue Research (2021)
-
Aggression and courtship differences found in Drosophila melanogaster from two different microclimates at Evolution Canyon, Israel
Scientific Reports (2019)
-
Putative adaptive inter-slope divergence of transposon frequency in fruit flies (Drosophila melanogaster) at “Evolution Canyon”, Mount Carmel, Israel
Biology Direct (2015)
-
Desiccation resistance along an aridity gradient in the cactophilic fly Drosophila buzzatii: sex-specific responses to stress
Evolutionary Ecology (2013)
-
Thermal strategies and energetics in two sympatric colubrid snakes with contrasted exposure
Journal of Comparative Physiology B (2010)
