
Abstract
Selective modulation of cell death is important for rational chemotherapy. By depleting Hsp90-client oncoproteins, geldanamycin (GA) and 17-allylamino-17-demethoxy-GA (17-AAG) (heat-shock protein-90-active drugs) render certain oncoprotein-addictive cancer cells sensitive to chemotherapy. Here we investigated effects of GA and 17-AAG in apoptosis-prone cells such as HL60 and U937. In these cells, doxorubicin (DOX) caused rapid apoptsis, whereas GA-induced heat-shock protein-70 (Hsp70) (a potent inhibitor of apoptosis) and G1 arrest without significant apoptosis. GA blocked caspase activation and apoptosis and delayed cell death caused by DOX. Inhibitors of translation and transcription and siRNA Hsp70 abrogated cytoprotective effects of GA. Also GA failed to protect HL60 cells from cytotoxicity of actinomycin D and flavopiridol (FL), inhibitors of transcription. We next compared cytoprotection by GA-induced Hsp70, caspase inhibitors (Z-VAD-fmk) and cell-cycle arrest. Whereas cell-cycle arrest protected HL60 cells from paclitaxel (PTX) but not from FL and DOX, Z-VAD-fmk prevented FL-induced apoptosis but was less effective against DOX and PTX. Thus, by inducing Hsp70, GA protected apoptosis-prone cells in unique and cell-type selective manner. Since GA does not protect apoptosis-reluctant cancer cells, we envision a therapeutic strategy to decrease side effects of chemotherapy without affecting its therapeutic efficacy.
Similar content being viewed by others
Introduction
Chemotherapy causes apoptosis in certain cancer and normal cells, leading to both therapeutic and side effects. Broadly, all cells could be subdivided into apoptosis-prone and reluctant. As a canonical example, HL60 cells are apoptosis prone. Thus, HL60 cells undergo apoptosis in response to chemotherapeutic agents with diverse mechanisms of action: doxorubicin (DOX) and etoposide (topoisomease-II inhibitors), paclitaxel (PTX; microtubule active agent), flavopiridol (FL; an inhibitor of transcription) and proteasomal inhibitors.1, 2, 3, 4, 5, 6 In contrast, apoptosis-reluctant cells, including common cancer cell lines, undergo either slow (nonapoptotic) types of cell death or cycle arrest.7 For example, K562 leukemia cells are apoptosis reluctant, because they express the Bcr-Abl antiapoptotic kinase, which in turn induces heat-shock protein-70 (Hsp70).8 Similarly, transfection of Hsp70 renders HL60 cells apoptosis reluctant.8, 9
Hsp70 blocks several steps of the apoptotic cascade: upstream from mitochondria, release of cytochrome C and apoptosis-inducing factor (AIF), nuclear import of AIF, activation of procaspases-9 and -3, and even downstream of active caspase-3.8, 10, 11, 12, 13, 14, 15 Hsp70 renders cells resistant to chemotherapy.8 Similarly, heat shock protects colon cancer cells from TRAIL-induced apoptosis.16 By inducing Hsps, heat shock can prevent apoptosis in cardiomyocytes.17, 18 Also, hyperthermia protected mice from tumor necrosis factor19 and gamma radiation.20 Yet, heat shock cannot be safely used in patients. Obviously, expression of ectopic Hsp70 is not suitable for clinical applications too. What is needed is pharmacological induction of Hsp70.
Geldanamycin (GA) and its analog 17-allylamino-17-demethoxy-GA (17-AAG) binds to heat-shock protein 90 (Hsp90).21 Hsp90 is a major repressor of the heat-shock transcription factor 1 (HSF1).22 By binding Hsp90, GA via HSF1 induces Hsp70, resembling heat shock response.22, 23 On the other hand, GA and 17-AAG are cytotoxic and can induce apoptosis in cancer cells. 17-AAG is undergoing clinical trials as an anticancer drug. Furthermore, numerous reports (including ours) have demonstrated that GA potentiated the cytotoxicity of some anticancer drugs.5, 24, 25, 26, 27, 28 Yet, in such studies, GA was used to deplete antiapoptotic kinases in apoptosis-reluctant cancer cells. The goal of this study is to investigate Hsp70-mediated effects in apoptosis-prone HL60 cells and to determine conditions for cell-type selective cytoprotection.
Results
GA induces Hsp70 in parallel with cytoprotection
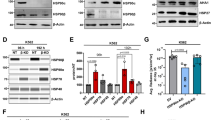
GA-induced Hsp70 in a dose-dependent manner (Figure 1a). In contrast, Hsp90 was unchanged. DOX neither induced Hsp70 nor prevented GA-induced Hsp70 (Figure 1a). Importantly that induction of Hsp70 correlated with the ability of GA to protect HL60 cells from DOX (Figure 1b). GA minimally inhibited cell survival, as measured by MTT assay (Figure 1b, open circles). In the absence of GA (X-axis: GA=0), 500 ng/ml DOX completely inhibited cell survival. GA abrogated DOX-induced cytotoxicity in a dose-dependent manner, coincident with Hsp70 induction (Figure 1).

Effects of GA on Hsp70 and cytotoxicity of DOX. (a) HL60 cells were treated with indicated concentrations of GA and 300 ng/ml DOX. After 16 h, Hsp70 and Hsp90 were measured by immunoblot. (b) HL60 cells were treated (in triplicate) with indicated concentrations of GA: either GA alone (empty circles) or GA plus 500 ng/ml DOX (closed circles). At GA=0, HL60 were treated with DOX alone or not treated (control). After 36 h, MTT assay was performed as described in Materials and Methods. Results are shown as percent of control (M±m)
GA prevents caspase-9 activation and apoptosis
In HL60 cells, DOX activated the intrinsic pathway of apoptosis that is manifested by cleavage of caspase-9 (Figure 2a). This was followed by PARP cleavage (Figure 2a) and the appearance of apoptotic morphology (Figure 2b) and cell death, as evidenced by cessation of cell metabolism (Figure 2b). GA did not cause cleavage of caspase-9 and PARP in HL60 cells (Figure 2a). GA was preferentially cytostatic at this time point (Figure 3 and data not shown). As expected, GA-induced Hsp70 and blocked caspase-9 activation and PARP cleavage caused by DOX (Figure 2a) and prevented nuclear fragmentation (Figure 2b). This in turn resulted in cytoprotection of HL60 cells (Figure 2c). Thus, GA protected HL60 cells by blocking the intrinsic apoptotic pathway activated by DOX.

Effects of DOX and GA on apoptosis: (a). HL60 cells were treated with 500 ng/ml DOX, 90 nM GA and their combination as indicated. After 16 h, immunoblot for Hsp70, Hsp90 and caspase-9 was performed. After 20 h, immunoblot for PARP was performed. (b) After 20 h, DAPI staining for nuclear fragmentation was performed as described in Materials and Methods. (c) After 36 h, MTT assay was performed as described in Materials and Methods

Effects of DOX and GA on TUNEL assay and cell-cycle distributions. HL60 cells were treated with 300 nM GA, 50 ng/ml DOX (50) or 500 ng/ml (500) either alone or in combinations. After 20 h, TUNEL assay and DNA contents were measured as described in Materials and Methods
GA prevented apoptosis but not cell-cycle arrest
GA arrested HL60 cells in G1 phase of the cell cycle (Figure 3). Low concentrations of DOX (50 ng/ml) cause G2 arrest in HL60 cells without appearance of TUNEL-positive cells. Cytotoxic concentrations of DOX (500 ng/ml) caused apoptosis, as evidenced by TUNEL-positive cells, associated with accumulation of cells with S-phase DNA content (Figure 3). Importantly, GA completely blocked apoptosis caused by 500 ng/ml DOX (Figure 3: GA+DOX 500).
GA and 17-AAG have similar effects on Hsp70 and cell death
Next we investigated whether other Hsp90-active agents induce Hsp70 and protect HL60 cells from DOX-induced cytotoxicity. 17-(allylamino)-17-demethoxygeldanamycin (17-AAG) is a less toxic analog of GA and is currently undergoing clinical trials. Like GA, 17-AAG induced Hsp70 in a dose-dependent manner (Figure 4a). Both agents abrogated cytotoxic effects of DOX, consistent with a dose-dependent induction of Hsp70. 17-AAG had two to three-fold lower potency compared with GA. Noteworthy, due to an excessive cytotoxicity, high concentrations (10 000 nM) of GA and 17-AAG could not sufficiently protect cells from DOX. Optimal protective concentrations were 500–2000 and 200–1000 nM for 17-AAG and GA, respectively (Figure 4b and data not shown).

Comparison of GA and 17-AAG. (a) HL60 cells were treated with indicated concentrations of GA and 17-AAG. After 16 h, cells were lysed and immonoblot for Hsp70 and actin was performed as described in Materials and Methods. (b) HL60 cells were treated with 500 ng/ml DOX plus the indicated concentrations of GA and 17-AAG. (Note: at concentration zero – there is DOX alone). After 36 h, MTT assay was performed as described in Materials and Methods. Results are shown as percent of control (untreated HL60 cells)
Cytoprotection by GA requires transcription and translation
In the presence of CHX, an inhibitor of translation, GA did not protect cells from DOX (Figure 5). Similarly, GA did not protect cells from ActD and FL (Figure 5). Both ActD and FL are potent inhibitors of transcription and prevented GA-induced Hsp70 (data not shown). GA protected cells from cytotoxicity of topo-I and topo-II inhibitors such as camptothecin (CAM), etoposide and metaxantrone (Figure 5 and data not shown). However, by damaging and intercalating DNA, very high concentrations of DOX and other DNA-damaging drugs can inhibit transcription. In agreement, GA did not protect HL60 cells from the cytotoxicity caused by DOX at concentrations above 2 μg/ml (Figure 6).

Effects of GA on the cytotoxicity of inhibitors of transcription and topoisomerase inhibitors. HL60 cells were incubated with indicated anticancer drugs (open bars) plus 300 nM GA (closed bars). DOX: 500 ng/ml doxorubicin; DOX+CHX (cycloheximide 10 μg/ml); ActD: 2 μg/ml actinomycin D; CAM: 500 nM camptothecin; FL: 500 nM flavopiridol. MTT assay was performed after 36 h

Cytoprotection from different concentrations of DOX. HL60 cells were incubated without (open circles) or with (closed circles) 300 nM GA and with the increasing concentrations of DOX. After 36 h, MTT assay was performed as described in Materials and Methods
siRNA Hsp70 abrogated cytoprotection
Thus, cytoprotection by GA requires transcription and inhibition of global transcription prevents cytoprotective effects. Next, we investigated whether specific abrogation of Hsp70 precludes cytoprotection. As shown in Figure 7, siRNA Hsp70 prevented the induction of Hsp70. These cells become sensitive to cytotoxicity of GA (Figure 7b). Furthermore, this abrogated cytoprotective effects of GA.

Effects of siRNA-Hsp70 on cytoprotective activity of GA. HL60 cells were transfected with vector (HL60/v) and with siRNA-Hsp70 (HL60/siRNA), as described in references 8 and 43. (a) Nontransfected HL60 cells (control), cells transfected with vector or siRNA-Hsp70 were treated with 500 nM GA (+) or left untreated (−). After 16 h, cells were lysed and immunoblot for Hsp70 was performed. (b) HL60/v and HL60/siHsp70 were treated with 500 nM GA, 500 ng/ml DOX, GA+DOX or left untreated (control). After 36 h, MTT assay was performed
Hsp70 protects cells from DOX, mimicking GA
Next, we investigated whether expression of Hsp70 substitutes for GA-mediated cytoprotection from DOX. We used both K562 cells with high endogenous Hsp70 and HL60/Hsp70 cells, transfected with Hsp70 (Figure 8, inset). K562 cells have high basal levels of Hsp70,8 which cannot be further induced by GA. Unlike HL60, K562 and HL60/Hsp70 cells were resistant to DOX (Figure 8). GA protected HL60 cells from DOX but did not affect HL60/Hsp70 cells, which were already protected by Hsp70 (Figure 8). Therefore, GA-mediated cytoprotection could not be observed in cells that are resistant to DOX (apoptosis-reluctant cells). Furthermore, GA slightly sensitized K562 cells to DOX. This can be explained by depletion of the antiapoptotic Bcr-Abl caused by GA, as shown previously.5, 8 In HL60/Hsp70 cells, resistance to GA was due to inhibited apoptosis, as shown in Figure 9. Whereas DOX caused nuclear fragmentation in parental HL60 cells, either Hsp70 or GA completely blocked it (Figure 9).

Effects of endogenous and ectopic Hsp70 on cell resistance to DOX and cytoprotective activity of GA. HL60 cells, HL60 cells were transfected with vector (HL60/Hsp70) and K562 cells were treated with 500 nM GA, 500 ng/ml DOX, GA+DOX or left untreated (control). After 36 h, MTT assay was performed. Inset: Basal levels of Hsp70 (−) and levels of Hsp70 following treatment with 500 nM GA for 16 h

Effects of Hsp70 overexpression on apoptosis. HL60 cells transfected with Hsp70 (HL60/Hsp70) and HL60 cells were treated with 500 ng/ml DOX, 1000 nM 17-AAG and their combination as indicated. After 20 h, DAPI staining for nuclear fragmentation was performed as described in Materials and Methods
Finally, like HL60, U937 leukemia cells are apoptosis prone and express low basal levels of Hsp70 (Figure 10a). In U937 cells, DOX causes rapid cell death with disintegration of apoptotic cells (Figure 10b). 17-AAG induced Hsp70 in U937 cells and prevented disintegration and death of DOX-treated U937 cells (Figure 10).

Cytoprotective effects of 17-AAG in U937 cells. U937 cells were treated with 500 ng/ml DOX, 1000 nM 17-AAG and their combination as indicated. (a) After 16 h, Hsp70 and Hsp90 were determined by immunoblot. (b) After 16 h, microphotograph of live culture was taken
Comparison of three types of cytoprotection
Besides Hsp70, GA also induces cell-cycle arrest in HL60 cells. Cell-cycle arrest can protect from the cytotoxicity of cycle-dependent chemotherapy. For example, low concentrations of DOX (50 ng/ml), by arresting HL60 cells in G2, prevented apoptosis caused by PTX and docetaxel.29 Here we compared cytoprotection caused by GA and low cytostatic concentrations of DOX (low DOX). Unlike GA, low DOX did not prevent cell death caused by high concentrations of DOX (Figure 11b). This indicates that apoptotic effects of DOX are not cell-cycle dependent. As expected, low DOX prevented cell death caused by PTX (Figure 11b). Low DOX did not prevent cell death caused by FL, suggesting that its cytotoxicity is not cell-cycle dependent. Whereas GA could not abrogate cytotoxicity caused by FL (FL inhibits transcription, preventing Hsp70 induction), pharmacological inhibitors of caspases such as Z-VAD-fmk were very effective (Figure 11c). Interestingly, Z-VAD-fmk poorly abrogated the cytotoxicity of DOX and PTX (Figure 11c). Finally, PMA (phorbol ester) induces macrophage-like differentiation of HL60 cells. These differentiated HL60 cells are resting and apoptosis reluctant.30 Not surprisingly, pre-treatment with PMA prevented the cytotoxicity of both PTX and DOX (Figure 11). Pre-treatment with PMA was less effective against FL, which, by inhibiting transcription, can prevent differentiation.

Comparison of cytoprotective activities of GA, low DOX, Z-VAD-fmk and PMA. First, HL60 cells were (a) treated with 300 nM GA, (b) pretreated for 12 h with 50 ng/ml DOX (low DOX), (c) treated with 20 μM Z-VAD-fmk or (d) pretreated for 24 h with 50 nM PMA (closed bars). Then, cells were treated with cytotoxic agents: either with 500 ng/ml DOX (DOX), or 500 nM flavopiridol (FL) or 60 nM paclitaxel (PTX). After 36 h, MTT assay was performed as described in Materials and Methods
Discussion
Here we showed that GA blocked the activation of the intrinsic apoptotic pathway and prolonged survival of HL60 cells treated with DOX and some other inhibitors of topoisomerases II and I (daunomycin, etoposide, camptothecin). GA did not protect HL60 cells from inhibitors of transcription ActD and FL. Like ActD, super-high concentrations of DOX (above 2 μM) inhibit transcription; and GA did not protect cells from super-high concentrations of DOX. Yet, such high concentrations of DOX are above therapeutic levels. Noteworthy, that GA did not protect HL60 cells from cisplatin and radiation (data not shown), which damage not only DNA but also proteins (and perhaps require intact Hsp90 functions for cytoprotection). In summary, GA protected against apoptosis-induced DNA-damaging topoisomerase inhibitors. The cytoprotective effect of GA correlated with Hsp70 induction. Both CXH and siRNA Hsp70 abrogated the cytoprotection. Conversely, transfection with Hsp70 rendered HL60 cells resistant to chemotherapy. Since DOX did not induce apoptosis in Hsp70-transfected cells, GA did not affect cytotoxicity of DOX in such cells. Taking together these data indicate that induction of Hsp70 is a main mechanism of cytoprotection by GA and 17-AAG.
In addition, cytoprotection can be achieved by cell-cycle arrest and synthetic caspase inhibitors. First, by causing G2 arrest, pre-treatment with low cytostatic (50 ng/ml) concentrations of DOX effectively protected cells from PTX but not FL and DOX. This further indicates that, unlike PTX, which kills cells strictly in mitosis, the cytotoxicity of both FL and DOX is not cell-phase dependent, at least in apoptosis-prone HL60 cells. Second, caspase inhibitors, such as Z-VAD-fmk, directly inhibit caspases and do not require transcription. Therefore, Z-VAD-fmk inhibited FL-induced apoptosis, whereas GA was absolutely ineffective. In contrast, GA demonstrated superior protection against DOX. In fact, Hsp70 can inhibit late stages of apoptosis, block events downstream from caspase-3.11 Also, Hsp70 inhibits caspase-independent events and lysosomal-mediated cell death31 and may be superior as an inhibitor of chemotherapy-induced cytotoxicity. Cytoprotection is clinically useful, only if it is selective, meaning that cancer cells are not protected. For example, cell-cycle-based cytoprotection exploits the differences in cell-cycle regulation between normal and cancer cells.32 Similarly, caspase inhibitors can protect selectively parental cells but not Pgp-expressing multidrug-resistant cells, which pump Z-VAD-fmk and Z-DEVD-fmk out.33 Cytoprotection by GA is selective: GA does not protect apoptosis-reluctant cancer and leukemia cells5, 27 including Hsp70-expressing cells (Figures 8 and 9). Furthermore, effects of GA are both cell-type and drug-sequence dependent.28 Effects of Hsp70 are also cell-type selective. For example, elevation of heat-shock protein did not attenuate radiation and etoposide-induced apoptosis in lung cancer cell lines.34 Furthermore, GA can induce apoptosis in oncogene-dependent cancer cells. Numerous antiapoptotic kinases such as Raf-1, EGF-R, ErbB, Akt, Bcr-Abl, depend on Hsp90 for their stability and activity. By promoting degradation of these kinases, GA causes cytostatic and cytotoxic effects.5, 26, 35, 36 Also, by depleting these oncoproteins, GA sensitizes cancer cells to chemotherapy.25, 37 For example, GA sensitizes Bcr-Abl-transfected HL60 cells to standard chemotherapeutic agents, such as DOX (a topoisomerase-II inhibitor).5 Our data can explain previously inexplicable observations in the literature. For example, it has been shown (as expected) that the kinase inhibitor herbimycin A sensitized Bcr-Abl-expressing cells to cytotoxicity of topoisomerase II inhibitor (etoposide). Unexpectedly, hebrimycin A protected HL60 and U937 cells from the cytotoxicity of etoposide.2 This can be explained taking into account that hebrimycin A is an Hsp90-active agent which, like GA, induces Hsp70.38
17-AAG and other Hsp90-active agents are currently undergoing clinical trials to treat common cancers.21, 39 It is expected that these agents will be especially effective in drug combinations. In selected types of cancer (oncogene-addictive cancers with Bcr-Abl, EGF-R), a combination of 17-AAG and DOX are synergistic. On the other hand, 17-AAG and DOX may be selectively antagonistic in certain apoptosis-prone normal cells. Most normal cells such as fibroblasts and smooth muscle cells are resistant to chemotherapy. It's toxicity to apoptosis-prone hematopoietic, mucosal, cardiomyocites, colonic epithelial, hair follicules that causes side effects of DOX. There is no adequate model for such apoptosis-prone cells in vitro. Therefore, HL60 and U937 cell lines (albeit leukemic) might represent cell types that are protectable by GA. There are three prerequisites for GA-mediated cytoprotection. First, in HL60 cells, basal Hsp70 levels are low and GA induces Hsp70. Second, in HL60 cells, DOX causes caspase-dependent apoptosis that is Hsp70 inhibitable. Third, GA by itself does not cause significant apoptosis in HL60 cells.
Noteworthy, GA is considered for therapy of neurodegenerative diseases due to its Hsp70-dependent antiapoptotic effects in neuronal cells.40 Similarly, we expect that Hsp70-inducing agents may decrease the toxicity of chemotherapy in apoptosis-prone normal (and cancer) cells. Simultaneously, GA may sensitize cancer cells that depend on antiapoptitcic kinase (Bcr-Abl, EGF-R, Akt) to DOX (Figure 12). Finally, DOX can induce nonapoptotic mitotic catastrophe and cell senescence in cancer cells.41 When mitotic catastrophe and cell senescence is the goal of therapy, then prevention of apoptosis in normal cells may decrease side effects without decreasing therapeutic efficacy of chemotherapy.

Dual effects of Hsp90-active agents on cell sensitivity to chemotherapy. By binding to Hsp90, GA causes depletion of antiapoptotic kinases (Raf-1, Bcr-Abl, Akt, EGF-R) and induces Hsp70. Therefore, they may simultaneously protect cells from and sensitize cells to chemotherapy. Sensitization is observed in apoptosis-reluctant cancer cells that are dependent on antiapoptotic hsp90 client kinases (Bcr-Abl, Akt). In contrast, in apoptosis-prone cells with low basal levels of Hsp70, GA antagonizes chemotherapy
Materials and Methods
Cell lines
HL60, U937, K562 human leukemia cell lines were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). HL60/Bcr-Abl and HL60/Hsp70 were described previously.5, 8, 42, 43 Reagents: Taxol was a Bristol-Myers product (Bristol-Myers, Princeton, NJ, USA). FL was obtained from the Development Therapeutics Program (NCI) and was prepared as 10 mM stock solution in DMSO. Cisplatin, camptotecin, FR901228, etoposide, GA and 17-(allylamino)-17-demethoxygeldanamycin (17-AAG) were obtained from the Developmental Therapeutics Program, National Cancer Institute (Bethesda, MD, USA). DOX (Adriamycin), actinomycin D (ActD), cycloheximide (CHX), etoposide and 4,5 dimethyl-2-yl 2,5-diphenyl tetrazolium bromide (MTT) were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Immunoblot analysis
Cells were lysed and soluble proteins harvested in TNES buffer and proteins resolved as previously described.5, 6 Immunoblot for PARP was performed using rabbit polyclonal anti-human PARP (Upstate Biotechnology, Lake Placid, NY, USA), for Bcr using monoclonal anti-human Bcr antibodies N 2 (Oncogene Science, Calbiochem, San Diego, CA, USA), mouse monoclonal antihuman tubulin and actin (Sigma, St. Louse, MO, USA), rabbit polyclonal anti-human Raf-1 (C12, Santa Cruz, CA, USA), monoclonal anti-human Hsp90 and Hsp70 (Upstate Biotechnology, NY, USA), mouse monoclonal anti-human caspase-9 (Pharmingen, San Diego, CA, USA) and caspase-3 (Transduction Lab., Lexington, KY, USA).
MTT assay
A total of 15 000 leukemia cells were plated in 0.1 ml in 96-well flat bottom plates and then exposed to tested agents (final volume 0.2 ml per well). At the indicated times, 20 μl of 5 mg/ml MTT solution in PBS were added to each well for 4 h. After removal of the medium, 170 μl of DMSO were added to each well to dissolve the formazan crystals. The absorbance at 540 nm was determined using a Biokinetics plate reader (Bio-Tek Instruments, Inc, Winooski, VT, USA). Triplicate wells were assayed for each condition and S.D. will be determined.
Flow cytometry
Cells were harvested, washed with PBS and resuspended in 75% ethanol in PBS and kept at 4°C for at least 30 min. Cells were resuspended and incubated for 30 min in propidium iodide staining solution containing 0.05 mg/ml propidium iodide (Sigma), 1 mM EDTA, 0.1% Triton-X-100 and 1 mg/ml RNAse A in PBS. The suspension was then analyzed on a Becton Dickinson FACScan. DNA content frequency histograms were measured using a FACScan flow cytometer (Becton Dickinson Immunocytometry Systems, San Jose, CA, USA). To calculate percentage of cells in respective phases of the cell cycle the DNA content frequency histograms were deconvoluted using the MultiCycle software (Phoenix Flow Systems, San Diego, CA, USA).
In situ DNA strand break labeling (TUNEL assay)
Cells were rinsed with PBS, fixed in 1% methanol-free formaldehyde for 15 min at room temperature and stored in 70% ethanol at −20°C for at least 1 h. The cells were then rinsed twice with PBS for 5 min. DNA strand break labeling was performed using the APO-BRDU kit provided by Phoenix Flow Systems (San Diego, CA, USA). After washing with PBS, cells were stained with propidium iodine (5 μg/ml PI) dissolved in PBS containing RNase A, for 20 min. Cellular fluorescence was measured using a FACScan flow cytometer (Becton-Dickinson).
Analysis of cell viability
Cells were incubated with Trypan blue and then blue (dead) cells and transparent (live) cells were counted in a hemocytometer.
Nuclear fragmentation assay
Cells were fixed with 90% ethanol with 10% glacial acid and stained with DAPI as described previously.5 Nuclei were visualized under UV microscopy.
Abbreviations
- AIF:
-
apoptosis-inducing factor
- 17-AAG:
-
17-allylamino-17-demethoxy-GA
- DOX:
-
doxorubicin
- FL:
-
flavopiridol
- GA:
-
geldanamycin
- Hsp70:
-
heat-shock protein-70
- Hsp90:
-
heat-shock protein-90
- HSF1:
-
heat-shock transcription factor1
- PTX:
-
paclitaxel
- PMA:
-
phorbol ester
References
Dubrez L, Goldwasser F, Genne P, Pommier Y and Solary E (1995) The role of cell cycle regulation and apoptosis triggering in determining the sensitivity of leukemic cells to topoisomerase I and II inhibitors. Leukemia 9: 1013–1024
Riordan FA, Bravery CA, Mengubas K, Ray N, Borthwick NJ, Akbar AN, Hart SM, Hoffbrand AV, Mehta AB and Wickremasinghe RG (1998) Herbimycin A accelerates the induction of apoptosis following etoposide treatment or gamma-irradiation of bcr/abl-positive leukaemia cells. Oncogene 16: 1533–1542
Gangemi RM, Santamaria B, Bargellesi A, Cosulich E and Fabbi M (2000) Late apoptotic effects of taxanes on K562 erythroleukemia cells: apoptosis is delayed upstream of caspase-3 activation. Int. J. Cancer 85: 527–533
An WG, Hwang SG, Trepel JB and Blagosklonny MV (2000) Protease inhibitor-induced apoptosis: accumulation wt p53, p21WAF1/CIP1, and induction of apoptosis are independent markers of proteasome inhibition. Leukemia 14: 1276–1283
Blagosklonny MV, Fojo T, Bhalla KN, Kim JS, Trepel JB, Figg WD, Rivera Y and Neckers LM (2001) The Hsp90 inhibitor geldanamycin selectively sensitizes Bcr-Abl-expressing leukemia cells to cytotoxic chemotherapy. Leukemia 15: 1537–1543
Demidenko ZN and Blagosklonny MV (2004) Flavopiridol induces p53 via initial inhibition of Mdm2 and p21 and, independently of p53, sensitizes apoptosis-reluctant cells to tumor necrosis factor. Cancer Res. 64: 3653–3660
Blagosklonny MV (2001) Paradox of Bcl-2 (and p53): why may apoptosis-regulating proteins be irrelevant to cell death ? Bioessays 23: 947–953
Guo F, Sigua C, Bali P, George P, Fiskus W, Scuto A, Annavarapu S, Mouttaki A, Sondarva G, Wei S, Wu J, Djeu J and Bhalla K (2005) Mechanistic role of heat shock protein 70 in Bcr-Abl-mediated resistance to apoptosis in human acute leukemia cells. Blood 105: 1246–1255
Kwak HJ, Jun CD, Pae HO, Yoo JC, Park YC, Choi BM, Na YG, Park RK, Chung HT, Chung HY, Park WY and Seo JS (1998) The role of inducible 70-kDa heat shock protein in cell cycle control, differentiation, and apoptotic cell death of the human myeloid leukemic HL-60 cells. Cell Immunol. 187: 1–12
Mosser DD, Caron AW, Bourget L, Denis-Larose C and Massie B (1997) Role of the human heat shock protein hsp70 in protection against stress-induced apoptosis. Mol. Cell Biol. 17: 5317–5327
Jaattela M, Wissing D, Kokholm K, Kallunki T and Egeblad M (1998) Hsp70 exerts its anti-apoptotic function downstream of caspase-3-like proteases. EMBO J. 17: 6124–6134
Beere HM, Wolf BB, Cain K, Mosser DD, Mahboubi A, Kuwana T, Tailor P, Morimoto RI, Cohen GM and Green DR (2000) Heat-shock protein 70 inhibits apoptosis by preventing recruitment of procaspase-9 to the Apaf-1 apoptosome. Nat. Cell Biol. 2: 469–475
Mosser DD, Caron AW, Bourget L, Meriin AB, Sherman MY, Morimoto RI and Massie B (2000) The chaperone function of hsp70 is required for protection against stress-induced apoptosis. Mol. Cell Biol. 20: 7146–7159
Ravagnan L, Gurbuxani S, Susin SA, Maisse C, Daugas E, Zamzami N, Mak T, Jaattela M, Penninger JM, Garrido C and Kroemer G (2001) Heat-shock protein 70 antagonizes apoptosis-inducing factor. Nat. Cell Biol. 3: 839–843
Gurbuxani S, Schmitt E, Cande C, Parcellier A, Hammann A, Daugas E, Kouranti I, Spahr C, Pance A, Kroemer G and Garrido C (2003) Heat shock protein 70 binding inhibits the nuclear import of apoptosis-inducing factor. Oncogene 22: 6669–6678
Ozoren N and El-Deiry W (2002) Heat shock protects HCT116 and H460 cells from TRAIL-induced apoptosis. Exp. Cell Res. 281: 175–181
Ito H, Shimojo T, Fujisaki H, Tamamori M, Ishiyama S, Adachi S, Abe S, Marumo F and Hiroe M (1999) Thermal preconditioning protects rat cardiac muscle cells from doxorubicin-induced apoptosis. Life Sci. 64: 755–761
Zou Y, Zhu W, Sakamoto M, Qin Y, Akazawa H, Toko H, Mizukami M, Takeda N, Minamino T, Takano H, Nagai T, Nakai A and Komuro I (2003) Heat shock transcription factor 1 protects cardiomyocytes from ischemia/reperfusion injury. Circulation 108: 3024–3030
Van Molle W, Wielockx B, Mahieu T, Takada M, Taniguchi T, Sekikawa K and Libert C (2002) HSP70 protects against TNF-induced lethal inflammatory shock. Immunity 16: 685–695
Shen RN, Hornback NB, Shidnia H, Wu B, Lu L and Broxmeyer HE (1991) Whole body hyperthermia: a potent radioprotector in vivo. Int. J. Radiat. Oncol. Biol. Phys. 20: 525–530
Workman P (2003) Overview: translating Hsp90 biology into Hsp90 drugs. Curr. Cancer Drug. Targets 3: 297–300
Zou J, Guo Y, Guettouche T, Smith DF and Voellmy R (1998) Repression of heat shock transcription factor HSF1 activation by HSP90 (HSP90 complex) that forms a stress-sensitive complex with HSF1. Cell 94: 471–480
Bagatell R, Paine-Murrieta GD, Taylor CW, Pulcini EJ, Akinaga S, Benjamin IJ and Whitesell L (2000) Induction of a heat shock factor 1-dependent stress response alters the cytotoxic activity of hsp90-binding agents. Clin. Cancer Res. 6: 3312–3328
Enmon R, Yang WH, Ballangrud AM, Solit DB, Heller G, Rosen N, Scher HI and Sgouros G (2003) Combination treatment with 17-N-allylamino-17-demethoxy geldanamycin and acute irradiation produces supra-additive growth suppression in human prostate carcinoma spheroids. Cancer Res. 63: 8393–8399
Jones DT, Addison E, North JM, Lowdell MW, Hoffbrand AV, Mehta AB, Ganeshaguru K, Folarin NI and Wickremasinghe RG (2004) Geldanamycin and herbimycin A induce apoptotic killing of B chronic lymphocytic leukemia cells and augment the cells' sensitivity to cytotoxic drugs. Blood 103: 1855–1861
George P, Bali P, Annavarapu S, Scuto A, Fiskus W, Guo F, Sigua C, Sondarva G, Moscinski L, Atadja P and Bhalla K (2005) Combination of the histone deacetylase inhibitor LBH589 and the hsp90 inhibitor 17-AAG is highly active against human CML-BC cells and AML cells with activating mutation of FLT-3. Blood 105: 1768–1776
Bisht KS, Bradbury CM, Mattson D, Kaushal A, Sowers A, Markovina S, Ortiz KL, Sieck LK, Isaacs JS, Brechbiel MW, Mitchell JB, Neckers LM and Gius D (2003) Geldanamycin and 17-allylamino-17-demethoxygeldanamycin potentiate the in vitro and in vivo radiation response of cervical tumor cells via the heat shock protein 90-mediated intracellular signaling and cytotoxicity. Cancer Res. 63: 8984–8995
Vasilevskaya IA, Rakitina TV and O'Dwyer PJ (2003) Geldanamycin and its 17-allylamino-17-demethoxy analogue antagonize the action of Cisplatin in human colon adenocarcinoma cells: differential caspase activation as a basis for interaction. Cancer Res. 63: 3241–3246
Demidenko ZN, Halicka D, Kunicki J, McCubrey JA, Darzynkiewicz Z and Blagosklonny MV (2005) Selective killing of adriamycin-resistant (G2 checkpoint-deficient and MRP1-expressing) cancer cells by docetaxel. Cancer Res. 65: 4401–4407
Solary E, Bertrand R, Kohn KW and Pommier Y (1993) Differential induction of apoptosis in undifferentiated and differentiated HL-60 cells by DNA topoisomerase I and II inhibitors. Blood 81: 1359–1368
Gyrd-Hansen M, Nylandsted J and Jäättelä M (2004) Heat Shock Protein 70 promotes cancer cell viability by safeguarding lysosomal integrity. Cell Cycle 3: 1484–1485
Blagosklonny MV and Pardee AB (2001) Exploiting cancer cell cycling for selective protection of normal cells. Cancer Res. 61: 4301–4305
Blagosklonny MV (2001) Treatment with inhibitors of caspases, that are substrates of drug transporters, selectively permits chemotherapy-induced apoptosis in multidrug-resistant cells but protects normal cells. Leukemia 15: 936–994
Ekedahl J, Joseph B, Marchetti P, Fauvel H, Formstecher P, Lewensohn R and Zhivotovsky B (2003) Heat shock protein 72 does not modulate ionizing radiation-induced apoptosis in U1810 non-small cell lung carcinoma cells. Cancer Biol. Ther. 2: 663–669
An WG, Schnur RC, Neckers LM and Blagosklonny MV (1997) Depletion of ErbB2, Raf-1 and mutant p53 proteins by geldanamycin derivatives correlates with antiproliferative activity. Cancer Chemother. Pharmacol. 40: 60–64
Nimmanapalli R, O'Bryan E, Kuhn D, Yamaguchi H, Wang HG and Bhalla KN (2003) Regulation of 17-AAG-induced apoptosis: role of Bcl-2, Bcl-XL, and Bax downstream of 17-AAG-mediated down-regulation of Akt, Raf-1, and Src kinases. Blood 102: 269–275
Yao Q, Nishiuchi R, Li Q, Kumar AR, Hudson WA and Kersey JH (2003) FLT3 expressing leukemias are selectively sensitive to inhibitors of the molecular chaperone heat shock protein 90 through destabilization of signal transduction-associated kinases. Clin. Cancer Res. 9: 4483–4493
Morris SD, Cumming DV, Latchman DS and Yellon DM (1996) Specific induction of the 70-kD heat stress proteins by the tyrosine kinase inhibitor herbimycin-A protects rat neonatal cardiomyocytes. A new pharmacological route to stress protein expression? J. Clin. Invest. 97: 706–712
Banerji U, O'Donnell A, Scurr M, Pacey S, Stapleton S, Asad Y, Simmons L, Maloney A, Raynaud F, Campbell M, Walton M, Lakhani S, Kaye S, Workman P and Judson I (2005) Phase I pharmacokinetic and pharmacodynamic study of 17-allylamino, 17-demethoxygeldanamycin in patients with advanced malignancies. J. Clin. Oncol. 23: 4152–4161
Klettner A (2004) The induction of heat shock proteins as a potential strategy to treat neurodegenerative disorders. Drug News Perspect. 17: 299–306
Roninson IB, Broude EV and Chang BD (2001) If not apoptosis, then what? Treatment-induced senescence and mitotic catastrophe in tumor cells. Drug Resist. Updat. 4: 303–313
Nimmanapalli R, O'Bryan E and Bhalla K (2001) Geldanamycin and its analogue 17-allylamino-17-demethoxygeldanamycin lowers Bcr-Abl levels and induces apoptosis and differentiation of Bcr-Abl-positive human leukemic blasts. Cancer Res. 61: 1799–1804
Guo F, Rocha K, Bali P, Pranpat M, Fiskus W, Boyapalle S, Kumaraswamy S, Balasis M, Lawrence N and Bhalla K (2005) Abrogation of hsp70 induction as a strategy to increase anti-leukemia activity of hsp90 inhibitor 17-allylamino-demethoxy geldanamycin. Cancer Res. in press
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by B Zhivotovsky
Rights and permissions
About this article
Cite this article
Demidenko, Z., Vivo, C., Halicka, H. et al. Pharmacological induction of Hsp70 protects apoptosis-prone cells from doxorubicin: comparison with caspase-inhibitor- and cycle-arrest-mediated cytoprotection. Cell Death Differ 13, 1434–1441 (2006). https://doi.org/10.1038/sj.cdd.4401812
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.cdd.4401812
Keywords
This article is cited by
-
The increased expression of the inducible Hsp70 (HSP70A1A) in serum of patients with heart failure and its protective effect against the cardiotoxic agent doxorubicin
Molecular and Cellular Biochemistry (2019)
-
BCN057 induces intestinal stem cell repair and mitigates radiation-induced intestinal injury
Stem Cell Research & Therapy (2018)
-
Lipid bodies containing oxidatively truncated lipids block antigen cross-presentation by dendritic cells in cancer
Nature Communications (2017)
-
HSP-90 inhibitor ganetespib is synergistic with doxorubicin in small cell lung cancer
Oncogene (2014)
-
Hsp70 regulates the doxorubicin-mediated heart failure in Hsp70-transgenic mice
Cell Stress and Chaperones (2014)
