
Abstract
Billions of songbirds migrate between continents twice each year, but the energy costs of this feat have never been measured for free-flying individuals. Here we follow New World Catharus thrushes during their nocturnal migratory flights1,2, recapturing individuals after journeys of up to 600 km and determining their energy expenditure by using doubly labelled water3,4. Although flight itself is costly, our results confirm the counterintuitive prediction that songbirds expend double the amount of energy during stopovers that they spend on flight over their entire migration5,6.
Similar content being viewed by others

Main
Quantifying the energy costs of migration should improve our understanding of songbird ecology7, but these costs have so far been estimated only from models8 and wind-tunnel studies6,8,9. To measure migration costs for free-flying Swainson's (Catharus ustulatus) and hermit (C. guttatus) thrushes, we injected 38 birds (each weighing about 30 g; ref. 10) with 0.1 ml doubly labelled water (containing 2H and 18O isotopes) in the morning and followed 12 throughout one nocturnal spring-migration flight for up to 600 km or 7.7 h (Fig. 1a)1,2; we followed six of these birds until they landed at a stopover habitat. Injected birds that did not make a migratory flight that night were treated as controls (n = 26). The next morning, we recaptured birds at 9:10 (± 1 h 40 min) by mist-netting them and took a blood sample to quantify their energy expenditure4.

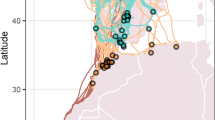
We pooled data for final analysis because all traits were statistically indistinguishable between species10. a, Tracks for 12 individuals starting from Urbana, Illinois: full lines, recaptured birds; dashed lines, lost birds. Ends of lines indicate stopover sites. b, Daily energy expenditure (DEE) of migrating (red symbols) and stopover (green symbols) birds; for migrating birds, this is described as a linear regression: DEE (kJ) = 70 (± 9) + 12.5 (± 2.1) × flight duration in hours; r2 = 0.89, P = 0.004 (standard deviations are given in parentheses) (ref. 11). Dashed line, average flight duration below which energy expenditure is indistinguishable from stopover controls (2.5 h). c, Variability in energy expenditure of birds during stopover is explained by differences in average daily ambient temperature (DEE (kJ) = 109.6 (± 6.5) − 1.52 (± 0.37) × ambient temperature; r2 = 0.4, n = 26, P < 0.001). We calibrated measurements using doubly labelled water with five birds held in a respirometer for 24 h (accuracy of ± 4% according to equation 7.17 of ref. 4).
Our measurements indicate that both sustained natural flight and stopover are costly in cool weather (Fig. 1b, c). Migratory flight used 12.5 kJ h−1, determined as flying cost above resting metabolism, or 15.5 kJ h−1 (4.3 watts) total energy while flying, which agrees with predicted values5,6,8; the cost of transport is 1.1 (calculated as a flight power of 4.3 watts divided by a flight speed of 13 m s−1 and a body weight of 0.3 N (equivalent to a body mass of 30 g)), which results in a mechanical cost of transport of about 0.25 (1.1 × 0.23, where 0.23 is the energy-conversion efficiency).
These results for free-flying birds validate a generation of theoretical models5,8 and wind-tunnel studies9,11. Surprisingly, stopover thrushes that did not fly on cold nights incurred increased thermoregulation costs comparable to those of a 2.5-h flight (Fig. 1b), illustrating the importance of climate during spring migration (Fig. 1c). Three of six migrating birds chose to stop over within 50 m of houses in urban or suburban areas.
We found that individual birds had roughly the same body weight and fat content in the mornings before and after their migratory flights (6% body-weight loss, no change in fat content), indicating that they probably regulate the duration of a migratory flight according to their body weight and fat stores6.
We can now estimate the energy expenditure of spring migration in a songbird11. Catharus thrushes can travel from Panama to Canada in 42 days10. Only 18 of 42 days include nocturnal flights, which are 265 km on average (4.6 h in flight; Fig. 1b). The average cost for migration days is 130 kJ, of which 71.3 kJ is spent on flight alone; the remaining 24 days are stopovers where an average of 88 kJ d−1 is expended, assuming an ambient temperature similar to that in central Illinois. For a 4,800-km migration, about 4,450 kJ is spent — an overall cost of some 0.93 kJ km−1.
Our measurements indicate that actual flight represents only 29% of total energy expenditure during the entire migration, confirming the predicted 1:2 ratio of energy costs during flight versus stopover6. These calculations do not take into account adverse weather conditions, which could strongly affect the energy balance, or the wide variation in stopover costs for different habitats7. Days that include migratory flight (130 kJ d−1) are significantly less expensive than days spent feeding large chicks, for example12.
We suggest that the timing of migration in relation to climate, particularly the ambient temperature, is important for birds that use twice as much energy during stopover as during migratory flight. Springtime reverse migration13 can now be better understood as an energy-saving strategy to minimize thermoregulation costs.
References
Cochran, W. in Animal Orientation and Navigation (eds Galler, S. R., Schmidt-Koenig, K., Jacobs, G. J. & Belleville, R. E.) 39–59 (NASA, Washington DC, 1972).
Cochran, W. Anim. Behav. 35, 927–929 (1987).
Nagy, K. A. Am. J. Physiol. 238, 454–473 (1980).
Speakman, J. R. Doubly Labeled Water: Theory and Practice (Chapman and Hall, London, 1997).
Hedenström, A. & Alerstam, T. J. Theor. Biol. 189, 227–234 (1997).
Alerstam, T. & Hedenström, A. J. Avian Biol. 29, 343–369 (1998).
Faaborg, J., Brittingham, M., Donovan, T. & Blake, J. in Ecology and Management of Neotropical Migratory Birds: A Synthesis and Review of Critical Issues (eds Martin, T. E. & Finch, D. M.) 357–380 (Oxford Univ. Press, New York, 1995).
Pennycuick. C. J. J. Theor. Biol. 191, 47–61 (1998).
Klaassen, M. J. Exp. Biol. 199, 57–64 (1996).
Graber, R. R., Graber, J. W. & Kirk, E. L. Biol. Notes No. 75 (Illinois Natural History Survey, Champaign, Illinois, 1971).
Masman, D. & Klaasen, M. Auk 104, 603–616 (1987).
Bennett, P. & Harvey, P. H. J. Zool. 213, 327–363 (1987).
Richardson, W. J. Oikos 30, 224–272 (1978).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Wikelski, M., Tarlow, E., Raim, A. et al. Costs of migration in free-flying songbirds. Nature 423, 704 (2003). https://doi.org/10.1038/423704a
Issue Date:
DOI: https://doi.org/10.1038/423704a
This article is cited by
-
Expression patterns of heat-shock genes during stopover and the trade-off between refueling and stress response in a passerine migrant
Journal of Comparative Physiology B (2024)
-
Pre-migratory flights in migrant songbirds: the ecological and evolutionary importance of understudied exploratory movements
Movement Ecology (2023)
-
Artificial light at night is a top predictor of bird migration stopover density
Nature Communications (2023)
-
Intelligence at any price? A criterion for defining AI
AI & SOCIETY (2023)
-
The Yellow-browed Warbler (Phylloscopus inornatus) as a model to understand vagrancy and its potential for the evolution of new migration routes
Movement Ecology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
