
Abstract
Steiner and Salvini-Plawen claim that Acaenoplax cannot be a mollusc because it would sit uneasily within the caudofoveates, solenogastres or polyplacophorans; however, the fossil record frequently reveals character combinations that have not survived to Recent times1. Although Acaenoplax displays a mosaic of molluscan characters that does not accord with any single modern group, this does not necessarily eliminate it from the phylum as a whole.
Similar content being viewed by others


Sutton et al. reply
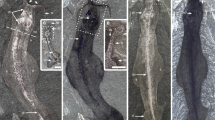
Aragonitic valves that grow by marginal accretion are highly characteristic of molluscs and were undoubtedly present in Acaenoplax. Valves of the closely related genera Arctoplax, Heloplax and Enetoplax occur in coeval deposits on Götland2. These are preserved in a manner that is identical to that of co-occurring molluscan shells of original aragonitic composition (such as the valves of polyplacophorans)2,3 and display clear co-marginal growth lines. The composition of the spines of Acaenoplax is less certain, but they were rigid and sharp (see Fig. 2d in ref. 4) and hence were probably biomineralized, and those that arise from the cuticle are preserved in the same manner as those that project from valves. Our use of the term 'cuticular' was intended to refer to the occurrence of the spicules on the cuticle, rather than to their composition.
The groove on the ventral surface is the one feature of Acaenoplax that appears to be more polychaete-like than molluscan, although this morphology is typically associated with errant rather than sedentary polychaetes. The other proposed polychaete characters are problematic. Although there is clearly a serial structure in Acaenoplax, there are no external signs of segmentation. Indeed, the overlap between subsequent 'lobe chevrons' is incompatible with true segmentation. The lobes and ridges of Acaenoplax are difficult to homologize with parapodial structures, as there are up to eight pairs of lobes per ridge. As spines arise from lobes, ridges and the cuticle (see Fig. 2d in ref. 4), they cannot be homologous with chaetae, which arise only from parapodia5.
Acaenoplax has multiple projections that emerge from a cavity at its larger termination. These features are very hard to homologize with polychaete posterior structures, so if Acaenoplax is a polychaete, then this termination must represent the anterior. Many sedentary polychaetes do have an anterior (prostomial) cavity from which feeding tentacles emerge5. There is, however, no evidence that the projections could extend significantly from the cavity and, with this anterior–posterior polarity, the spine array faces forwards, which is unlikely on functional grounds. By contrast, interpreting this termination as an aplacophoran-like mantle cavity containing gills is compatible with both the observed mobility of the projections and the direction of the spines.
Several Acaenoplax structures (such as lobes, ridges and the postero–ventral plate) lack convincing polychaete or molluscan homologues, and hence are considered to be autapomorphic and phylogenetically uninformative. Nonetheless, this leaves four molluscan characters (polyplacophoran-like aragonitic scleritome, spicules arising from cuticle and probably composed of aragonite, serial rather than segmented structure, and posterior cavity with projections) arrayed against one polychaete-like character (the ventral surface). We consider that the balance of evidence places Acaenoplax firmly within the Mollusca rather than in the Polychaeta.
References
Wills, M. A., Briggs, D. E. G., Fortey, R. A., Wilkinson, M. & Sneath, P. H. A. in Fossils and Phylogeny (ed. Edgecombe, G. E.) 33–105 (Columbia Univ. Press, New York, 1997).
Cherns, L. Palaeontology 41, 939–974 (1998).
Cherns, L. Palaeontology 41, 545–573 (1998).
Sutton, M. D., Briggs, D. E. G., Siveter, D. J. & Siveter, D. J. Nature 410, 461–463 (2001).
Ruppert, E. E. & Barnes, R. D. in Invertebrate Zoology 6th edn, 508–554 (Saunders College, Orlando, Florida, 1994).
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Sutton, M., Briggs, D., Siveter, D. et al. Acaenoplax — polychaete or mollusc?. Nature 414, 602 (2001). https://doi.org/10.1038/414602a
Issue Date:
DOI: https://doi.org/10.1038/414602a
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
