
Key Points
-
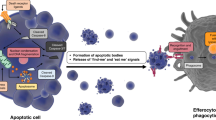
Classic apoptosis is characterized by a stereotyped series of morphological changes, including chromatin condensation and rapid uptake of the corpses by neighbouring cells. Since many forms of programmed cell death (PCDs), including autophagy, paraptosis, dark cell death and 'necrosis-like PCD' result in a non-apoptotic morphology, the biochemical pathways of PCD might be far more diverse than first predicted. Other proteases are proposed to take over the role of caspases for cleaving for example nuclear lamins and cytoskeletal elements.
-
There are many distinct forms of non-apoptotic PCD. In 'apoptosis-like PCD', chromatin may condense, but not to the characteristic geometric shape associated with classic apoptosis; in 'necrosis-like PCD' chromatin may not condense at all. All of them may be mediated by classic death receptors and result in the display of 'eat-me' signals.
-
The classic role of caspases in promoting PCD is now being refined as more cases are found in which caspases are shown to control the morphology of cell death rather than the actual decision to die. In other examples of PCD they are not required at all. This has led to the proposal that other evolutionary ancient and conserved pathways exist.
-
Mitochondria can trigger cell death by activation of different pathways, which may be caspase-dependent or independent. Although these pathways may be triggered simultaneously, the final outcome and form of cell death will depend on the individual cell type, the initial triggers, and the metabolic situation.
-
The control of apoptosis is often misregulated in tumours and, although many cancer therapies induce classic apoptosis, potential drugs that engage other PCD routes are emerging.
-
In the adult nervous system, it is crucial that the induction of PCD is tightly controlled to prevent unnecessary damage. For this reason, there seem to be additional layers of control for caspase activation. For instance, cells might first require a 'competence to die' signal before apoptosis can be subsequently activated, and often caspase-independent death pathways are preferentially activated.
Abstract
A single family of proteases, the caspases, has long been considered the pivotal executioner of all programmed cell death. However, recent findings of evolutionarily conserved, caspase-independent controlled death mechanisms have opened new perspectives on the biology of cell demise, with particular implications for neurobiology, cancer research and immunological processes.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


References
Kerr, J. F. R., Wyllie, A. H. & Currie, A. R. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 26, 239–257 (1972).
Hengartner, M. O. The biochemistry of apoptosis. Nature 407, 770–776 (2000).
Strasser, A., O'Connor, L. & Dixit, V. M. Apoptosis signaling. Annu. Rev. Biochem. 69, 217–245 (2000).
Wright, S. C. et al. Activation of CPP32-like proteases is not sufficient to trigger apoptosis: inhibition of apoptosis by agents that suppress activation of AP24, but not CPP32-like activity. J. Exp. Med. 186, 1107–1117 (1997).
Lacana, E., Ganjei, J. K., Vito, P. & D'Adamio, L. Dissociation of apoptosis and activation of IL-1β-converting enzyme/Ced-3 proteases by ALG-2 and the truncated Alzheimer's gene ALG-3. J. Immunol. 158, 5129–5135 (1997).
Jäättelä, M., Wissing, D., Kokholm, K., Kallunki, T. & Egeblad, M. Hsp70 exerts its anti-apoptotic function downstream of caspase-3-like proteases. EMBO J. 17, 6124–6134 (1998).
De Maria, R. et al. Negative regulation of erythropoiesis by caspase-mediated cleavage of GATA-1. Nature 401, 489–493 (1999).
Harvey, K. J., Lukovic, D. & Ucker, D. S. Caspase-dependent Cdk activity is a requisite effector of apoptotic death events. J. Cell Biol. 148, 59–72 (2000).
Foghsgaard, L. et al. Cathepsin B acts as a dominant execution protease in tumor cell apoptosis induced by tumor necrosis factor. J. Cell Biol. 153, 999–1009 (2001).The first identification of a protease (cathepsin B) that mediates death-receptor-induced caspase-independent apoptosis-like PCD. The requirement of cathepsin B for phosphatidylserine exposure, blebbing and final execution in TNF-induced caspase-dependent death is also shown. Provides a table of specificity profiles for commonly used protease inhibitors.
Roberts, L. R., Adjei, P. N. & Gores, G. J. Cathepsins as effector proteases in hepatocyte apoptosis. Cell. Biochem. Biophys. 30, 71–88 (1999).
Lavoie, J. N., Nguyen, M., Marcellus, R. C., Branton, P. E. & Shore, G. C. E4orf4, a novel adenovirus death factor that induces p53-independent apoptosis by a pathway that is not inhibited by zVAD-fmk. J. Cell Biol. 140, 637–645 (1998).
Berndt, C., Mopps, B., Angermuller, S., Gierschik, P. & Krammer, P. H. CXCR4 and CD4 mediate a rapid CD95-independent cell death in CD4+ T cells. Proc. Natl Acad. Sci. USA 95, 12556–12561 (1998).
Borner, C. & Monney, L. Apoptosis without caspases: an inefficient molecular guillotine. Cell Death Differ. 6, 497–507 (1999).
Kitanaka, C. & Kuchino, Y. Caspase-independent programmed cell death with necrotic morphology. Cell Death Differ. 6, 508–515 (1999).
Mathiasen, I. S., Lademann, U. & Jäättelä, M. Apoptosis induced by vitamin D compounds in breast cancer cells is inhibited by Bcl-2 but does not involve known caspases or p53. Cancer Res. 59, 4848–4856 (1999).
Nylandsted, J. et al. Selective depletion of heat shock protein 70 (Hsp70) activates a tumor-specific death program that is independent of caspases and bypasses Bcl-2. Proc. Natl Acad. Sci. USA 97, 7871–7876 (2000).The first demonstration of an endogenous protein (Hsp70) that is specifically required for the survival of tumorigenic cells. The transformation-associated death programme inhibited by Hsp70 is shown to be caspase independent.
Elliott, K., Ge, K., Du, W. & Prendergast, G. C. The c-Myc-interacting adaptor protein Bin1 activates a caspase-independent cell death program. Oncogene 19, 4669–4684 (2000).
Joza, N. et al. Essential role of the mitochondrial apoptosis-inducing factor in programmed cell death. Nature 410, 549–554 (2001).Gene deletion of Aif in mice. Genetic evidence for the essential role of AIF in developmental apoptosis.
Leist, M., Single, B., Castoldi, A. F., Kuhnle, S. & Nicotera, P. Intracellular adenosine triphosphate (ATP) concentration: a switch in the decision between apoptosis and necrosis. J. Exp. Med. 185, 1481–1486 (1997).Initial demonstration that initially similar cell-death signals are switched between apoptosis and programmed necrosis by metabolic conditions in the cell.
Vercammen, D. et al. Dual signaling of the Fas receptor: initiation of both apoptotic and necrotic cell death pathways. J. Exp. Med. 188, 919–930 (1998).Demonstrates the ability of tumour necrosis factor receptor 1 and Fas to trigger caspase-independent programmed necrosis, mediated by the production of mitochondrial reactive oxygen species.
Leist, M. et al. Inhibition of mitochondrial ATP generation by nitric oxide switches apoptosis to necrosis. Exp. Cell Res. 249, 396–403 (1999).
Chung, S., Gumienny, T. L., Hengartner, M. O. & Driscoll, M. A common set of engulfment genes mediates removal of both apoptotic and necrotic cell corpses in C. elegans. Nature Cell Biol. 2, 931–937 (2000).
Hirt, U. A., Gantner, F. & Leist, M. Phagocytosis of nonapoptotic cells dying by caspase-independent mechanisms. J. Immunol. 164, 6520–6529 (2000).Shows that cells dying by different caspase-independent mechanisms can be recognized by phagocytes in various ways and can be efficiently ingested.
Aravind, L., Dixit, V. M. & Koonin, E. V. Apoptotic molecular machinery: vastly increased complexity in vertebrates revealed by genome comparisons. Science 291, 1279–1284 (2001).
Frohlich, K. U. & Madeo, F. Apoptosis in yeast — a monocellular organism exhibits altruistic behaviour. FEBS Lett. 473, 6–9 (2000).
Ameisen, J. C. The origin of programmed cell death. Science 272, 1278–1279 (1996).
Wyllie, A. H. & Golstein, P. More than one way to go. Proc. Natl Acad. Sci. USA 98, 11–13 (2001).
Ellis, H. M. & Horvitz, H. R. Genetic control of programmed cell death in nematode C. elegans. Cell 44, 817–829 (1986).
Los, M., Wesselborg, S. & Schulze-Osthoff, K. The role of caspases in development, immunity, and apoptotic signal transduction: lessons from knockout mice. Immunity 10, 629–639 (1999).
Nicotera, P., Leist, M. & Manzo, L. Neuronal cell death: a demise with different shapes. Trends Pharmacol. Sci. 20, 46–51 (1999).
Woodle, E. S. et al. Anti-human class I MHC antibodies induce apoptosis by a pathway that is distinct from the Fas antigen-mediated pathway. J. Immunol. 158, 2156–2164 (1997).
McCarthy, N. J., Whyte, M. K. B., Gilbert, C. S. & Evan, G. I. Inhibition of Ced-3/ICE-related proteases does not prevent cell death induced by oncogenes, DNA damage, or the Bcl-2 homologue Bak. J. Cell Biol. 136, 215–227 (1997).Shows that caspase activation functions as a general switch between necrosis and apoptosis, but is not required for death itself.
Mateo, V. et al. CD47 ligation induces caspase-independent cell death in chronic lymphocytic leukemia. Nature Med. 5, 1277–1284 (1999).
Holler, N. et al. Fas triggers an alternative, caspase-8-independent cell death pathway using the kinase RIP as effector molecule. Nature Immunol. 1, 489–495 (2000).The first description of the requirement of RIP kinase activity for the death-receptor-induced necrotic pathway. Also indicates that programmed necrosis might be the dominant death mode induced by Fas in activated primary lymphocytes.
Schulze-Osthoff, K. et al. Cytotoxic activity of tumor necrosis factor is mediated by early damage of mitochondrial functions. Evidence for the involvement of mitochondrial radical generation. J. Biol. Chem. 267, 5317–5323 (1992).
Ha, H. C. & Snyder, S. H. Poly(ADP-ribose) polymerase is a mediator of necrotic cell death by ATP depletion. Proc. Natl Acad. Sci. USA 96, 13978–13982 (1999).
Bursch, W. et al. Active cell death induced by the anti-estrogens tamoxifen and ICI 164 384 in human mammary carcinoma cells (MCF-7) in culture: the role of autophagy. Carcinogenesis 17, 1595–1607 (1996).
Chi, S. et al. Oncogenic Ras triggers cell suicide through the activation of a caspase-independent cell death program in human cancer cells. Oncogene 18, 2281–2290 (1999).
Roach, H. I. & Clarke, N. M. Physiological cell death of chondrocytes in vivo is not confined to apoptosis. New observations on the mammalian growth plate. J. Bone Joint Surg. Br. 82, 601–613 (2000).
Turmaine, M. et al. Nonapoptotic neurodegeneration in a transgenic mouse model of Huntington's disease. Proc. Natl Acad. Sci. USA 97, 8093–8097 (2000).
Deas, O. et al. Caspase-independent cell death induced by anti-CD2 or staurosporine in activated human peripheral T lymphocytes. J. Immunol. 161, 3375–3383 (1998).
Luschen, S., Ussat, S., Scherer, G., Kabelitz, D. & Adam-Klages, S. Sensitization to death receptor cytotoxicity by inhibition of FADD/caspase signaling: requirement of cell cycle progression. J. Biol. Chem. 275, 24670–24678 (2000).
Volbracht, C., Leist, M., Kolb, S. A. & Nicotera, P. Apoptosis in caspase-inhibited neurons. Mol. Med. 7, 36–48 (2001).
Xue, L., Fletcher, G. C. & Tolkovsky, A. M. Autophagy is activated by apoptotic signalling in sympathetic neurons: an alternative mechanism of death execution. Mol. Cell Neurosci. 14, 180–198 (1999).
Xiang, J., Chao, D. T. & Korsmeyer, S. J. Bax-induced cell death may not require interleukin 1β-converting enzyme-like proteases. Proc. Natl Acad. Sci. USA 93, 14559–14563 (1996).The first demonstration of caspase-independent programmed cell death induced by Bax-like molecules.
Khwaja, A. & Tatton, L. Resistance to the cytotoxic effects of tumor necrosis factor-α can be overcome by inhibition of a FADD/caspase-dependent signaling pathway. J. Biol. Chem. 274, 36817–36823 (1999).
Chautan, M., Chazal, G., Cecconi, F., Gruss, P. & Golstein, P. Interdigital cell death can occur through a necrotic and caspase-independent pathway. Curr. Biol. 9, 967–970 (1999).
Matsumura, H. et al. Necrotic death pathway in Fas receptor signaling. J. Cell Biol. 151, 1247–1256 (2000).
Amarante-Mendes, G. P. et al. Anti-apoptotic oncogenes prevent caspase-dependent and independent commitment for cell death. Cell Death Differ. 5, 298–306 (1998).
Chua, B. T., Guo, K. & Li, P. Direct cleavage by the calcium-activated protease calpain can lead to inactivation of caspases. J. Biol. Chem. 275, 5131–5135 (2000).
Lankiewicz, S. et al. Activation of calpain I converts excitotoxic neuron death into a caspase-independent cell death. J. Biol. Chem. 275, 17064–17071 (2000).
Jäättelä, M. Escaping cell death: survival proteins in cancer. Exp. Cell Res. 248, 30–43 (1999).
Smith, K. G., Strasser, A. & Vaux, D. L. CrmA expression in T lymphocytes of transgenic mice inhibits CD95 (Fas/APO-1)-transduced apoptosis, but does not cause lymphadenopathy or autoimmune disease. EMBO J. 15, 5167–5176 (1996).
Doerfler, P., Forbush, K. A. & Perlmutter, R. M. Caspase enzyme activity is not essential for apoptosis during thymocyte development. J. Immunol. 164, 4071–4079 (2000).
Kunstle, G. et al. Concanavalin A hepatotoxicity in mice: tumor necrosis factor-mediated organ failure independent of caspase-3-like protease activation. Hepatology 30, 1241–1251 (1999).
Savill, J. & Fadok, V. Corpse clearance defines the meaning of cell death. Nature 407, 784–788 (2000).
Wolf, B. B. et al. Calpain functions in a caspase-independent manner to promote apoptosis-like events during platelet activation. Blood 94, 1683–1692 (1999).
Mattson, M. P. Apoptosis in neurodegenerative disorders. Nature Rev. Mol. Cell Biol. 1, 120–129 (2000).
Leverrier, Y. & Ridley, A. J. Apoptosis: caspases orchestrate the ROCK 'n' bleb. Nature Cell Biol. 3, E91–E93 (2001).
Deiss, L. P., Galinka, H., Berissi, H., Cohen, O. & Kimchi, A. Catepsin D protease mediates programmed cell death induced by interferon-γ, Fas/APO-1 and TNF-α. EMBO J. 15, 3861–3870 (1996).
Guicciardi, M. E. et al. Cathepsin B contributes to TNF-α-mediated hepatocyte apoptosis by promoting mitochondrial release of cytochrome c. J. Clin. Invest. 106, 1127–1137 (2000).
Waterhouse, N. J. et al. Calpain activation is upstream of caspases in radiation-induced apoptosis. Cell Death Differ. 5, 1051–1061 (1998).
Vanags, D. M., Porn-Ares, M. I., Coppola, S., Burgess, D. H. & Orrenius, S. Protease involvement in fodrin cleavage and phosphatidylserine exposure in apoptosis. J. Biol. Chem. 271, 31075–31085 (1996).
Johnson, D. E. Noncaspase proteases in apoptosis. Leukemia 14, 1695–1703 (2000).A review that profoundly discusses different proteases and the technologies used to identify their contribution to cell death.
Leist, M. & Jäättelä, M. Triggering of apoptosis by cathepsins. Cell Death Differ. 8, 324–326 (2001).
Squier, M. K., Miller, A. C., Malkinson, A. M. & Cohen, J. J. Calpain activation in apoptosis. J. Cell Physiol. 159, 229–237 (1994).
Yamashima, T. Implication of cysteine proteases calpain, cathepsin and caspase in ischemic neuronal death of primates. Prog. Neurobiol. 62, 273–295 (2000).
Wang, K. K. Calpain and caspase: can you tell the difference? Trends Neurosci. 23, 20–26 (2000).
Roberg, K. Relocalization of cathepsin D and cytochrome c early in apoptosis revealed by immunoelectron microscopy. Lab. Invest. 81, 149–158 (2001).
Schotte, P., Declercq, W., Van Huffel, S., Vandenabeele, P. & Beyaert, R. Non-specific effects of methyl ketone peptide inhibitors of caspases. FEBS Lett. 442, 117–121 (1999).
Laster, S. M., Wood, J. G. & Gooding, L. R. Tumor necrosis factor can induce both apoptic and necrotic forms of cell lysis. J. Immunol. 141, 2629–2634 (1988).
Kawahara, A., Ohsawa, Y., Matsumura, H., Uchiyama, Y. & Nagata, S. Caspase-independent cell killing by Fas-associated protein with death domain. J. Cell Biol. 143, 1353–1360 (1998).
Yang, X., Koshravi-Far, R., Chang, H. Y. & Baltimore, D. Daxx, a novel Fas-binding protein that activates JNK and apoptosis. Cell 89, 1067–1076 (1997).
Chang, H. Y., Nishitoh, H., Yang, X., Ichijo, H. & Baltimore, D. Activation of apoptosis signal-regulating kinase 1 (ASK1) by the adapter protein Daxx. Science 181, 1860–1863 (1998).
Tobiume, K. et al. ASK1 is required for sustained activations of JNK/p38 MAP kinases and apoptosis. EMBO Rep. 2, 222–228 (2001).
Tournier, C. et al. Requirement of JNK for stress-induced activation of the cytochrome c-mediated death pathway. Science 288, 870–874 (2000).
Charette, S. J., Lavoie, J. N., Lambert, H. & Landry, J. Inhibition of Daxx-mediated apoptosis by heat shock protein 27. Mol. Cell. Biol. 20, 7602–7612 (2000).
Susin, S. A. et al. Molecular characterization of mitochondrial apoptosis-inducing factor. Nature 397, 441–446 (1999).Cloning and cell biological characterization of the mitochondrial caspase-independent programmed cell death mediator AIF.
Suter, M. et al. Age-related macular degeneration. The lipofusion component N-retinyl-N-retinylidene ethanolamine detaches proapoptotic proteins from mitochondria and induces apoptosis in mammalian retinal pigment epithelial cells. J. Biol. Chem. 275, 39625–39630 (2000).
Daugas, E. et al. Mitochondrio-nuclear translocation of AIF in apoptosis and necrosis. FASEB J. 14, 729–739 (2000).
Miller, T. M. et al. Bax deletion further orders the cell death pathway in cerebellar granule cells and suggests a caspase-independent pathway to cell death. J. Cell Biol. 139, 205–217 (1997).
Hirsch, T. et al. The apoptosis–necrosis paradox. Apoptogenic proteases activated after mitochondrial permeability transition determine the mode of cell death. Oncogene 15, 1573–1581 (1997).
Finn, J. T. et al. Evidence that Wallerian degeneration and localized axon degeneration induced by local neurotrophin deprivation do not involve caspases. J. Neurosci. 20, 1333–1341 (2000).
Schierle, G. S. et al. Differential effects of Bcl-2 overexpression on fibre outgrowth and survival of embryonic dopaminergic neurons in intracerebral transplants. Eur. J. Neurosci. 11, 3073–3081 (1999).
Tamatani, M. et al. ORP150 protects against hypoxia/ischemia-induced neuronal death. Nature Med. 7, 317–323 (2001).
Schmitt, C. A. & Lowe, S. W. Apoptosis and therapy. J. Pathol. 187, 127–137 (1999).
Kerr, J. F. R., Winterford, C. M. & Harmon, B. V. Apoptosis. Its significance in cancer and cancer therapy. Cancer 73, 2013–2026 (1994).
Lavoie, J. N., Champagne, C., Gingras, M. C. & Robert, A. Adenovirus E4 open reading frame 4-induced apoptosis involves dysregulation of Src family kinases. J. Cell Biol. 150, 1037–1056 (2000).
Nylandsted, J., Brand, K. & Jäättelä, M. Heat shock protein 70 is required for the survival of cancer cells. Ann. NY Acad. Sci. 926, 122–125 (2000).
Duffy, M. J. Proteases as prognostic markers in cancer. Clin. Cancer Res. 2, 613–618 (1996).
Alexander, C. M., Howard, E. W., Bissell, M. J. & Werb, Z. Rescue of mammary epithelial cell apoptosis and entactin degradation by a tissue inhibitor of metalloproteinases-1 transgene. J. Cell Biol. 135, 1669–1677 (1996).
Quignon, F. et al. PML induces a novel caspase-independent death process. Nature Genet. 20, 259–265 (1998).The first demonstration of a tumour-suppressor protein (PML) that activates a caspase-independent PCD.
Wang, Z. G. et al. PML is essential for multiple apoptotic pathways. Nature Genet. 20, 266–272 (1998).
Wang, Q., Yang, W., Uytingco, M. S., Christakos, S. & Wieder, R. 1,25-Dihydroxyvitamin D3 and all-trans-retinoic acid sensitize breast cancer cells to chemotherapy-induced cell death. Cancer Res. 60, 2040–2048 (2000).
Mathiasen, I. S., Hansen, C. M., Foghsgaard, L. & Jäättelä, M. Sensitization to TNF-induced apoptosis by 1,25-dihydroxy vitamin D(3) involves up-regulation of the TNF receptor 1 and cathepsin B. Int. J. Cancer 93, 224–231 (2001).
Raff, M. C. Social controls on cell survival and cell death. Nature 356, 397–400 (1992).
Johnson, M. D., Kinoshita, Y., Xiang, H., Ghatan, S. & Morrison, R. S. Contribution of p53-dependent caspase activation to neuronal cell death declines with neuronal maturation. J. Neurosci. 19, 2996–3006 (1999).
Deshmukh, M., Kuida, K. & Johnson, E. M. Jr Caspase inhibition extends the commitment to neuronal death beyond cytochrome c release to the point of mitochondrial depolarization. J. Cell Biol. 150, 131–143 (2000).
Stadelmann, C. et al. Activation of caspase-3 in single neurons and autophagic granules of granulovacuolar degeneration in Alzheimer's disease. Evidence for apoptotic cell death. Am. J. Pathol. 155, 1459–1466 (1999).
Hansson, O. et al. Transgenic mice expressing a Huntington's disease mutation are resistant to quinolinic acid-induced striatal excitotoxicity. Proc. Natl Acad. Sci. USA 96, 8727–8732 (1999).
Yamamoto, A., Lucas, J. J. & Hen, R. Reversal of neuropathology and motor dysfunction in a conditional model of Huntington's disease. Cell 101, 57–66 (2000).
Jellinger, K. A. & Stadelmann, C. H. The enigma of cell death in neurodegenerative disorders. J. Neural Transm. S21–S36 (2000).
Xue, L., Fletcher, G. C. & Tolkovsky, A. M. Mitochondria are selectively eliminated from eukaryotic cells after blockade of caspases during apoptosis. Curr. Biol. 11, 361–365 (2001).
Sperandio, S., de Belle, I. & Bredesen, D. E. An alternative, nonapoptotic form of programmed cell death. Proc. Natl Acad. Sci. USA 97, 14376–14381 (2000).Definition of paraptosis as a mode of neuronal death that is independent of caspase activity. Contains numerous references on alternative death modes.
Fujikawa, D. G. Confusion between neuronal apoptosis and activation of programmed cell death mechanisms in acute necrotic insults. Trends Neurosci. 23, 410–411 (2000).
Roy, M. & Sapolsky, R. Neuronal apoptosis in acute necrotic insults: why is this subject such a mess? Trends Neurosci. 22, 419–422 (1999).
Colbourne, F., Sutherland, G. R. & Auer, R. N. Electron microscopic evidence against apoptosis as the mechanism of neuronal death in global ischemia. J. Neurosci. 19, 4200–4210 (1999).
Pennacchio, L. A. et al. Progressive ataxia, myoclonic epilepsy and cerebellar apoptosis in cystatin B-deficient mice. Nature Genet. 20, 251–258 (1998).
Sagot, Y. et al. Bcl-2 overexpression prevents motoneuron cell body loss but not axonal degeneration in a mouse model of a neurodegenerative disease. J. Neurosci. 15, 7727–7733 (1995).
Wellington, C. L. & Hayden, M. R. Caspases and neurodegeneration: on the cutting edge of new therapeutic approaches. Clin. Genet. 57, 1–10 (2000).
Bergsmedh, A. et al. Horizontal transfer of oncogenes by uptake of apoptotic bodies. Proc. Natl Acad. Sci. USA 98, 6407–6411 (2001).
Boise, L. H. & Collins, C. M. Salmonella-induced cell death: apoptosis, necrosis or programmed cell death? Trends Microbiol. 9, 64–67 (2001).
Hentze, H. et al. In vivo and in vitro evidence for extracellular caspase activity released from apoptotic cells. Biochem. Biophys. Res. Commun. 283, 1111–1117 (2001).
Torriglia, A. et al. A caspase-independent cell clearance program. The LEI/L–DNase II pathway. Ann. NY Acad. Sci. 926, 192–203 (2000).
Acknowledgements
We acknowledge our colleagues for stimulating discussions and the Danish Cancer Society, the German Research Council and the Danish Medical Research Council for financial support. We also apologize to those whose work could only be cited indirectly.
Author information
Authors and Affiliations
Supplementary information
Movie 1
Time lapse imaging of blebbing process in caspase-independent death. WEHI-S murine fibrosarcoma cells were recorded by time lapse videomicroscopy with Varel contrast optics; all sequences correspond to about 1 h in real time. Nuclear morphology was recorded in parallel after staining with H-33342 (not shown; see Ref. 9). Exposure to tumour necrosis factor and activation of caspases. Intensive zeiosis (average active blebbing time per cell = 20 min). Formation of a terminal bleb marks cell death (energy failure) associated with chromatin condensation.
Please note that this movie may take a few seconds to download.
Movie 2
Time lapse imaging of blebbing process in caspase-independent death. WEHI-S murine fibrosarcoma cells were recorded by time lapse videomicroscopy with Varel contrast optics; all sequences correspond to about 1 h in real time. Nuclear morphology was recorded in parallel after staining with H-33342 (not shown; see Ref. 9). Exposure to tumour necrosis factor in the presence of the caspase-inhibitor zVADfmk. Intensive zeiosis (average active blebbing time per cell = 20 min). Formation of a terminal bleb marks cell death (energy failure) associated with chromatin condensation.
Please note that this movie may take a few seconds to download.
Movie 3
Time lapse imaging of blebbing process in caspase-independent death. WEHI-S murine fibrosarcoma cells were recorded by time lapse videomicroscopy with Varel contrast optics; all sequences correspond to about 1 h in real time. Nuclear morphology was recorded in parallel after staining with H-33342 (not shown; see Ref. 9). Simple cell lysis (necrosis) triggered by 2-mM chloroquine. No change in nuclear morphology, no zeiosis.
Please note that this a large (3MB) movie which may take a while to download.
Movie 4
Time lapse imaging of blebbing process in caspase-independent death. WEHI-S murine fibrosarcoma cells were recorded by time lapse videomicroscopy with Varel contrast optics; all sequences correspond to about 1 h in real time. Nuclear morphology was recorded in parallel after staining with H-33342 (not shown; see Ref. 9). Oligomycin-induced passive formation of a simple terminal bleb (energy failure) in the absence of zeiosis and cell movement. No change in nuclear morphology.
Please note that this movie may take a few seconds to download.
Related links
Related links
DATABASE LINKS
FURTHER INFORMATION
Glossary
- CASPASES
-
Family of cysteine proteases that can be divided into inflammatory caspases, and pro-apoptotic caspases, which can be further grouped into initiator and effector caspases.
- PHAGOCYTOSIS
-
Uptake of dying cells by macrophages or neighbouring cells. Recognition of 'eat-me' signals by specific receptors on the phagocytosing cell.
- DEATH RECEPTORS
-
A family of cell-surface receptors that can mediate cell death upon ligand-induced trimerization.
- OEDEMA
-
Water accumulation and swelling within a tissue, cell or organelle.
- ZEIOSIS
-
Dynamic plasma membrane blebbing of a dying cell, analogous to the bubbling of fermenting yeast.
- PHOSPHATIDYLSERINE EXPOSURE
-
Translocation of phosphatidylserine, which is confined to the inner leaflet of the plasma membrane in healthy cells, to the outside of the plasma membrane where it is recognized by a specific receptor on macrophages.
- PARAPTOSIS
-
A form of programmed cell death without prominent chromatin condensation and mainly characterized by cytoplasmic vacuolization.
- MALE LINKER CELL
-
The linker cell is a male-specific cell at the tip of the developing gonad. It 'guides' growth of the male gonad from the midbody region towards the tail. When the gonad has reached the tail late in larval development, the linker cell is 'murdered' by one of two neighbouring cells.
- DARK CELL DEATH
-
Slow neuronal death observed, for example, during Huntington's disease. Characterized by strong cytoplasmic condensation, chromatin clumping, ruffling of the cell membrane, but no blebbing of the nucleus or plasma membrane.
- PARP
-
Poly(ADP)-ribose polymerase. A nuclear enzyme activated by DNA damage and reducing cellular ATP levels when overactivated.
- STAUROSPORINE
-
Plant-derived cytotoxin known to trigger mitochondria-dependent apoptosis in most cell types. Model apoptosis inducer.
- COLCHICINE
-
A microtubule-depolymerizing poison. Leads to loss of neurites in neurons and to apoptosis in most cell types.
- PAN-CASPASE INHIBITORS
-
Cell-permeable irreversible inhibitors of all caspases examined so far that block or retard caspase-dependent processes. Widely used examples include z-Val-Ala-Asp-fluoromethylkethone (z-VAD-fmk) and Boc-Asp-fluoromethylketone.
- INHIBITOR OF APOPTOSIS PROTEIN
-
(IAP). A class of proteins (IAP, XIAP, NAIP) containing a BIR domain that can act as an intracellular caspase inhibitor.
- DEATH DOMAIN
-
A conserved sequence motif first identified in the intracellular parts of death receptors. Later recognized as the key motif for association of the receptors with cytosolic death-domain-containing proteins (FADD, TRADD, RIP) and the induction of cell death.
- REACTIVE OXYGEN SPECIES
-
(ROS). Collective term comprising intracellularly formed classic oxygen radicals and peroxides.
- APOPTOSOME
-
Multiprotein complex containing cytochrome c, Apaf-1 and procaspase-9. Catalyses ATP-dependent auto-activation of caspases by induced proximity. Key regulatory step of developmental apoptosis.
- AIF
-
Apoptosis-inducing factor, a flavoprotein normally located in the mitochondrial intermembrane space. It is released during apoptosis and is involved in nuclear changes and death induction.
- ONCOPROTEINS
-
Oncoproteins are encoded by oncogenes and their increased activity or expression promotes tumorigenesis either by inducing proliferation or by inhibiting cell death.
- PTEN
-
A tumour-suppressor protein with phosphatase activity specific for inositol phospholipids. Negative regulator of protein kinase B (Akt) pathway in cells.
- PYKNOSIS
-
Poorly defined pathological term for nuclear condensation. Mainly used to describe forms of non-apoptotic chromatin condensation.
- EXCITOTOXICITY
-
Neuronal death triggered by overexcitation of ion channel-gating (ionotropic) receptors (for example, ionotropic glutamate receptors). Release of endogenous excitotoxins triggers excitotoxic processes as common mechanism of neuronal loss during cerebral ischaemia.
- HORIZONTAL TRANSFER
-
Non-germline transmission of genetic material.
Rights and permissions
About this article
Cite this article
Leist, M., Jäättelä, M. Four deaths and a funeral: from caspases to alternative mechanisms. Nat Rev Mol Cell Biol 2, 589–598 (2001). https://doi.org/10.1038/35085008
Issue Date:
DOI: https://doi.org/10.1038/35085008
This article is cited by
-
Nickel oxide nanoparticles synthesized by Rose hip extract exert cytotoxicity against the HT-29 colon cancer cell line through the caspase-3/caspase-9/Bax pathway
Emergent Materials (2023)
-
Effects of a P2X7 receptor antagonist on myenteric neurons in the distal colon of an experimental rat model of ulcerative colitis
Histochemistry and Cell Biology (2022)
-
D,L-Methadone enhances the cytotoxic activity of standard chemotherapeutic agents on pediatric rhabdomyosarcoma
Journal of Cancer Research and Clinical Oncology (2022)
-
Enhanced lysosomal function is critical for paclitaxel resistance in cancer cells: reversed by artesunate
Acta Pharmacologica Sinica (2021)
-
Nanotechnology improves the therapeutic efficacy of gemcitabine against a human hepatocellular carcinoma cell line and minimizes its in vivo side effects
Naunyn-Schmiedeberg's Archives of Pharmacology (2021)
