
Abstract
The acquisition of neural fate by embryonic ectodermal cells is a fundamental step in the formation of the vertebrate nervous system. Neural induction seems to involve signalling by fibroblast growth factors (FGFs) and attenuation of the activity of bone morphogenetic protein (BMP)1,2,3,4. But FGFs, either alone or in combination with BMP antagonists, are not sufficient to induce neural fate in prospective epidermal ectoderm of amniote embryos1,3,4. These findings suggest that additional signals are involved in the specification of neural fate. Here we show that the state of Wnt signalling is a critical determinant of neural and epidermal fates in the chick embryo. Continual Wnt signalling blocks the response of epiblast cells to FGF signals, permitting the expression and signalling of BMP to direct an epidermal fate. Conversely, a lack of exposure of epiblast cells to Wnt signals permits FGFs to induce a neural fate.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

References
Harland, R. Neural induction. Curr. Opin. Genet. Dev. 10, 357–362 (2000).
Wilson, P. A. & Hemmati-Brivanlou, A. Induction of epidermis and inhibition of neural fate by Bmp-4. Nature 376, 331–333 (1995).
Wilson, S. I., Graziano, E., Harland, R., Jessell, T. M. & Edlund, T. An early requirement for FGF signalling in the acquisition of neural cell fate in the chick embryo. Curr. Biol. 10, 421–429 (2000).
Streit, A., Berliner, A. J., Papanayotou, C., Sirulnik, A. & Stern, C. D. Initiation of neural induction by FGF signalling before gastrulation. Nature 406, 74–78 (2000).
Noordermeer, J., Klingensmith, J., Perrimon, N. & Nusse, R. dishevelled and armadillo act in the wingless signalling pathway in Drosophila. Nature 367, 80–83 (1994).
Kelly, G. M., Erezyilmaz, D. F. & Moon, R. T. Induction of a secondary embryonic axis in zebrafish occurs following the overexpression of β-catenin. Mech. Dev. 53, 261–273 (1995).
Guger, K. A. & Gumbiner, B. M. β-Catenin has Wnt-like activity and mimics the Nieuwkoop signaling center in Xenopus dorsal–ventral patterning. Dev. Biol. 172, 115–125 (1995).
Roeser, T., Stein, S. & Kessel, M. Nuclear β-catenin and the development of bilateral symmetry in normal and LiCl-exposed chick embryos. Development 126, 2955–2965 (1999).
Eyal-Giladi, H. & Kochav, S. From cleavage to primitive streak formation: a complementary normal table and a new look at the first stages of the development of the chick. I. General morphology. Dev. Biol. 49, 321–337 (1976).
Hsieh, J. C., Rattner, A., Smallwood, P. M. & Nathans, J. Biochemical characterization of Wnt-frizzled interactions using a soluble, biologically active vertebrate Wnt protein. Proc. Natl Acad. Sci. USA 96, 3546–3551 (1999).
Shimizu, H. et al. Transformation by Wnt family proteins correlates with regulation of beta-catenin. Cell Growth Differ. 8, 1349–1358 (1997).
Rex, M. et al. Dynamic expression of chicken Sox2 and Sox3 genes in ectoderm induced to form neural tissue. Dev. Dyn. 209, 323–332 (1997).
Mallamaci, A., Di Blas, E., Briata, P., Boncinelli, E. & Corte, G. OTX2 homeoprotein in the developing central nervous system and migratory cells of the olfactory area. Mech. Dev. 58, 165–178 (1996).
Streit, A. & Stern, C. D. Establishment and maintenance of the border of the neural plate in the chick: involvement of FGF and BMP activity. Mech. Dev. 82, 51–66 (1999).
Sheng, G. & Stern, C. D. Gata2 and Gata3: novel markers for early embryonic polarity and for non-neural ectoderm in the chick embryo. Mech. Dev. 87, 213–216 (1999).
Pera, E., Stein, S. & Kessel, M. Ectodermal patterning in the avian embryo: epidermis versus neural plate. Development 126, 63–73 (1999).
New, D. A. A new technique for the cultivation of the chick embryo in vitro. J. Embryol. Exp. Morphol. 3, 320–331 (1955).
Mohammadi, M. et al. Structures of the tyrosine kinase domain of fibroblast growth factor receptor in complex with inhibitors. Science 276, 955–960 (1997).
Dufort, D., Schwartz, L., Harpal, K. & Rossant, J. The transcription factor HNF3beta is required in visceral endoderm for normal primitive streak morphogenesis. Development 125, 3015–3025 (1998).
Wood, H. B. & Episkopou, V. Comparative expression of the mouse Sox1, Sox2 and Sox3 genes from pre-gastrulation to early somite stages. Mech. Dev. 86, 197–201 (1999).
Kishi, M. et al. Requirement of Sox2-mediated signaling for differentiation of early Xenopus neuroectoderm. Development 127, 791–800 (2000).
Pevny, L. H., Sockanathan, S., Placzek, M. & Lovell-Badge, R. A role for SOX1 in neural determination. Development 125, 1967–1978 (1998).
Zorn, A. M. et al. Regulation of Wnt signaling by Sox proteins: XSox17 α/β and XSox3 physically interact with β-catenin. Mol. Cell 4, 487–498 (1999).
Shibamoto, S. et al. Cytoskeletal reorganization by soluble Wnt-3a protein signalling. Genes Cells 3, 659–670 (1998).
Koenig, B. B. et al. Characterization and cloning of a receptor for BMP-2 and BMP-4 from NIH 3T3 cells. Mol. Cell. Biol. 14, 5961–5974 (1994).
ten Dijke, P. et al. Characterization of type I receptors for transforming growth factor-beta and activin. Science 264, 101–104 (1994).
Schaeren-Wiemers, N. & Gerfin-Moser, A. A single protocol to detect transcripts of various types and expression levels in neural tissue and cultured cells: in situ hybridization using digoxigenin-labelled cRNA probes. Histochemistry 100, 431–440 (1993).
Hamburger, V. & Hamilton, H. L. A series of normal stages in the development of the chick embryo. J. Morph. 88, 49–92 (1951).
Read, E. M. et al. Evidence for non-axial A/P patterning in the nonneural ectoderm of Xenopus and zebrafish pregastrula embryos. Int. J. Dev. Biol. 42, 763–774 (1998).
Acknowledgements
We thank members of the Edlund lab for helpful discussions. We are very grateful to J. Nathans for providing the Wnt8 cell line and mFrz8CRD–IgG plasmid; R. Lovell-Badge and E. Boncinelli for antibodies; C. Healy and M. Kessel for chick Sox3 and DLX-5, respectively; J. C. Hsieh for technical advice; and H. Alstermark for technical assistance. T.E. is supported by the Swedish Medical Research Council and by the Foundation for Strategic Research. T.M.J. is supported by grants from the NIH. T.M.J. and R.N. are Investigators of the Howard Hughes Medical Institute.
Author information
Authors and Affiliations
Corresponding author
Supplementary information
Main Figure Legends: Detailed Quantitation.
Main Figure 1. RT-PCR analysis of marker gene expression in medial and lateral epiblast explants.
RT-PCR analysis of RNA isolated from stage X-XIII (EK X-XIII) explants.
(a) Wnt3A and Wnt8C are not expressed in stage XII-XIII (M) explants but are expressed in stage X (L) explants before culture. Fgf receptor2 (FgfR2) and the Wnt receptor Frizzled8 are expressed in both stage X- XIII (M) and (L) explants before culture.
(b) Fgf3 is expressed in stage XII-XIII (M) explants after 40h of culture and is down-regulated in explants cultured in the presence of BMP4.
(c) Fgf3 is expressed in stage XII-XIII (M) but not stage X (L) explants after culture. Expression of Fgf3 is downregulated in stage XII-XIII (M) explants cultured in the presence of Wnt3A and upregulated in stage X (L) explants cultured in the presence of mFrz8CRD-IgG for 40h. Expression of Bmp4 is downregulated in stage XII-XIII (M) explants after 40 hr of culture. Repression of Bmp4 expression is blocked in stage XII-XIII (M) explants cultured in the presence of Wnt3A for 40h. Bmp4 is expressed in stage X (L) explants after 40 hr of culture but is down-regulated in stage X (L) explants cultured in the presence of mFrz8CRD-IgG for 40h.
(d) Mesoderm is not induced in stage X (L) (EK X) explants exposed to mFrz8CRD-IgG. The mesodermal markers Brachyury, Chordin and Raldh2 are expressed in stage 4 (HH 4) whole embryo but not in stage X (L) explants cultured in the presence of mFrz8CRD-IgG for 40 hours.
(e) Endoderm is not induced in stage X (L) (EK X) explants exposed to mFrz8CRD-IgG. The endodermal marker HNF3β is detected in stage 4 mesendoderm but is not present in stage X (L) explants cultured in the presence of mFrz8CRD-IgG for 40 hours.
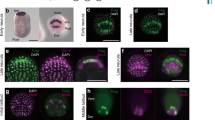
Main Figure 2. Regulation of neural and epidermal fate in medial epiblast explants
(a) Schematic representation of a stage XII (EK XII) embryo. Boxed area indicates the region of medial (M; red) explants used for in vitro studies.
(b-f) Stage XII (M) explants cultured for 40h in the presence of control conditioned medium (n = 6) generate Sox2+ cells (b), Sox3+ cells (c) and Otx2+ cells (d) (90 % ± 10 % cells/section; n = 20 sections), but do not generate Msx1/2+ cells (e) or GATA2+ cells (f).
(g-k) Stage XII (M) explants (n = 6) cultured in the presence of Wnt3A (1x) do not generate Sox2+ cells (g), Sox3+ cells (h) and Otx2+ cells (i) but generate Msx1/2+ cells (90 % ± 10 % cells/section; n = 9 sections; [ j] ) and GATA2+ cells (30 % ± 10 % cells/section; n = 9 sections [ k] ).
(l-p) Stage XII (M) explants (n = 6) cultured in the presence of Wnt3A (1x) and FGF2 (30 nM) do not generate Sox2+ cells (l), Sox3+ cells (m) and Otx 2+ cells (n) but generate Msx1/2+ cells (95 % ± 10 % cells/section; n = 5 sections; [ o] ). Under these conditions no GATA2+ cells (p) cells are generated which may reflect previous findings that FGF can pattern the epidermal ectoderm, resulting in down-regulation of GATA2.
(q-u) Stage XII (M) explants cultured in the presence of both Wnt3A (1 x) conditioned medium and Noggin (50 nM) generate Sox2+ cells (q), Sox3+ cells (r) and Otx2+ cells (s) (80 % ± 20 % cells/section; n = 10 sections) but few Msx1/2+ cells (10 % ± 5 % cells/section ; n = 10 sections; [ t] ) and no GATA2+ cells (u).
(v-z) The FGF receptor tyrosine kinase inhibitor SU5402 inhibits the acquisition of neural character.
Stage XII (M) explants (n = 6) when cultured in the presence of SU5402 (5µ M) do not generate Sox3+ cells (v), Sox2+ cells (w) or Otx2+ cells (x) but generate Msx1/2+ cells (70% ± 10% cells/section [ y] ) and GATA2+ cells (30% ± 10% cells/section [ z] ).
(a’-e’) Stage XII (M) explants cultured in the presence of both SU5402 (5 µ M) and Noggin (50 nM) do not generate Sox3+ cells (a’), Sox2+ cells (b’) and Otx2+ cells (c’) but generate Msx1/2+ cells (60 % ± 15 % cells/section ; n = 7 sections; [ d’] ) and GATA2+ cells (e’).
(f’-j’) Stage XII (M) explants (n = 6) cultured in the presence of Wnt3A (3x) do not generate Sox2+ cells (f’), Sox3+ cells (g’) and Otx 2+ cells (h’) but generate Msx1/2+ cells (90 % ± 10 % cells/section; n = 8 sections; [ i’] ) and GATA2+ cells (60 % ± 10 % cells/section; n = 6 sections [ j’] ).
(k’-o’) Stage XII (M) explants (n = 6) cultured in the presence of Wnt3A (3x) and Noggin (50 nM) do not generate Sox2+ cells (k’), Sox3+ cells (l’) and Otx 2+ cells (m’) but generate Msx1/2+ cells (90 % ± 10 % cells/section; n = 5 sections; [ n’] ) and GATA2+ cells (40 % ± 20 % cells/section; n = 5 sections [ o’] ).
Figure 3. Regulation of neural and epidermal fate in lateral epiblast explants.
(a) Schematic representation of a stage X (EK X) embryo. Boxed area indicates region of lateral (L; green) explants used for in vitro studies.
(b-f) Stage X (L) explants explants cultured for 40h in the presence of control conditioned medium (n = 6) do not generate Sox2+ cells (b), Sox3+ cells (c) or Otx 2+ cells (d) but generate Msx1/2+ cells (90 % ± 10 % cells/section; n = 15 sections; [ e] ) and GATA2+ cells (70 % ± 10 % cells/section; n = 15 sections; [ f] ).
(g-k) Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium (n = 6) generate Sox2+ cells (g), Sox3+ cells (h) and Otx2+ cells (i) (90 % ± 20 % cells/section; n = 15 sections), but do not generate Msx1/2+ cells (j) or GATA2+ cells (k). N.B. Explants isolated from the area opaca of stage X embryos grown in the presence of mFrz8CRD-IgG conditioned medium (n = 6) generate Sox2+ cells, Sox3+ cells and Otx2+ cells, but do not generate Msx1/2+ cells or GATA2+ cells.
(l-p) Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium and SU5402 (2 µ M) do not generate Sox2+ cells (l), Sox3+ cells (m) or Otx2+ cells (n) but generate Msx1/2+ cells (80 % ± 20 % cells/section; n = 6 sections; [ o] ) and GATA2+ cells (40 % ± 20 % cells/section; n = 6 sections; [ p] ).
(q-u) Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium and BMP4 do not generate Sox2+ cells (q), Sox3+ cells (r) or Otx2+ cells (s) but generate Msx1/2+ cells (80 % ± 10 % cells/section; n = 7 sections; [ t] ) and GATA2+ cells (65 % ± 20 % cells/section; n = 7 sections; [ u] ).
(v-z) Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium, SU5402 (2 µ M) and Noggin (50nM) generate Sox3+ cells (v), Sox3+ cells (w) or Otx2+ cells (x) (90 % ± 20 % cells/section; n = 6 sections) but do not generate Msx1/2+ (y) and GATA2+ (z).
(a'-e') Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium and SU5402 (5 µ M) do not generate Sox2+ cells (a'), Sox3+ cells (b') or Otx2+ cells (c') but generate Msx1/2+ cells (70 % ± 30 % cells/section; n = 6 sections; [ d'] ) and GATA2+ cells (30 % ± 20 % cells/section; n = 6 sections; [ e'] ).
(f'-j') Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium and SU5402 (5 µ M) and Noggin (50nM) do not generate Sox2+ cells (f'), Sox3+ cells (g') or Otx2+ cells (h') but generate Msx1/2+ cells (i') and GATA2+ cells (j').
Methods
Primers and conditions for RT-PCR: S17, Brachyury, Chordin, Fgf3 and Bmp4 primers and conditions are as described previously. Oligonucleotide primers against the chicken Wnt8C gene (5’tg aaa tac gac caa gcg ca 3’; 5’ta tct cct gtg gcc ttt gt 3’), Wnt3A (5’att ctg tcg gaa cta tgt gg 3’; 5’ctt cag atg cat gag ttc aa 3’), Raldh2 (5’ct aca ggg act caa aag gt 3’; 5’gga agg aag aag gag ata at 3’), FgfR2 (5’ccg ctg tat gtg att gta ga 3’; 5’tag cac acc aaa tga cca ta 3’) and Frizzled8 (5’gat gcc tct gtc tca agt gt 3’; 5’tat ccc aaa aac ctg tag ttc 3’) were used at 55° C. S17 was amplified for 25 cycles and all other samples were subjected to 30-35 cycles of amplification.
Supplementary Figures and Legends
Supplementary Figure 1. Neural and epidermal differentiation in New culture.

To test whether Wnts also block neural and induce epidermal character in medial epiblast cells in intact chick embryos maintained in New culture , aggregates of control mouse fibroblast L cells, or L cells expressing Wnt3A, were implanted into the medial region of the epiblast of stage X-XI embryos. Embryos were maintained in New culture for ~24h (until HH stage 3+-4) , and the expression of the neural marker Sox3 and the epidermal marker Dlx5 was monitored by whole mount in situ hybridisation.
(a) Schematic showing the site (yellow) where either mouse L-cells or L-cells expressing mouse Wnt3A were grafted in to the medial region of stage X - XI (EK X- XI) embryos.
(b) Sox3 is expressed in prospective neural plate cells in embryos implanted with control cells (arrow). Dotted line indicates the outer border of the neural plate.
(c) Sox3 expression in prospective neural plate cells is suppressed in embryos implanted with Wnt3A expressing cells (arrow). Dotted line indicates the outer border of the neural plate.
(d) Dlx5 is expressed in prospective epidermal but not in prospective neural cells in embryos implanted with control cells (arrow). Dotted line indicates the outer border of the prospective neural plate.
(e) Expression of Dlx5 is induced in prospective neural plate cells in embryos implanted with Wnt3A expressing cells (arrow). Dotted line indicates the outer border of the neural plate.
To examine whether mFrz8CRD-IgG also induces neural and blocks epidermal character in prospective epidermal cells in chick embryos maintained in New culture, aggregates of HEK293 cells transfected with a control plasmid or with a mFrz8CRD-IgG expression construct were implanted into the lateral region of the epiblast of stage X-XI embryos and maintained in New culture for ~24h until HH stage 3+-4, and expression of the neural marker Sox3 and the epidermal marker Dlx5 was monitored.
(f) Schematic showing the sites (yellow) where either HEK293 cells or HEK293 cells expressing Frz8CRD-IgG were grafted in to lateral regions of stage X - XI (EK X- XI) embryos.
(g) Sox3 is expressed in prospective neural but not in prospective epidermal cells in embryos implanted with control cells (arrow). Dotted line indicates the outer border of the neural plate.
(h) Expression of Sox3 is induced in prospective epidermis in embryos implanted with mFrz8CRD-IgG expressing cells (arrow). Dotted line indicates the outer border of the neural plate.
(i) Dlx5 is expressed in prospective epidermis in embryos implanted with control cells (arrow). Dotted line indicates the outer border of the neural plate.mFrz8CRD-IgG
(j) Expression of Dlx5 is suppressed in prospective epidermis in embryos implanted with mFrz8CRD-IgG expressing cells (arrow). Dotted line indicates the outer border of the neural plate.
Supplementary Figure 2. FGF and BMP antagonists are unable to induce neural fate in lateral epiblast cells unless Wnt signalling is attenuated.

(a) Schematic representation of a stage X (EK X) embryo. Boxed area indicates region of lateral (L; green) explants used for in vitro studies.
(b-f) Stage X (L) explants explants cultured for 40h in the presence of control conditioned medium (n = 6) do not generate Sox2+ cells (b), Sox3+ cells (c) or Otx 2+ cells (d) but generate Msx1/2+ cells (90 % ± 10 % cells/section; n = 15 sections; [ e] ) and GATA2+ cells (70 % ± 10 % cells/section; n = 15 sections; [ f] ).
(g-k) Stage X (L) explants cultured for 40h in the presence of Noggin (50 nM) (n = 6) do not generate Sox2+ cells (g), Sox3+ cells (h) or Otx 2+ cells (i) but generate Msx1/2+ cells (80 % ± 20 % cells/section; n = 6 sections; [ j] ) and GATA2+ cells cells (50 % ± 20 % cells/section; n = 6 sections; [ k] ).
(l-p) Stage X (L) explants cultured for 40h in the presence of FGF (30 nM) (n = 6) do not generate Sox2+ cells (l), Sox3+ cells (m) or Otx 2+ cells (n) but generate Msx1/2+ cells (60 % ± 30 % cells/section; n = 6 sections; [ o] ). Under these conditions no GATA2+ cells (p) cells are generated which may reflect previous findings that FGF can pattern the epidermal ectoderm, resulting in down-regulation of GATA2.
(q-u) Stage X (L) explants cultured for 40h in the presence of FGF (30 nM) and Noggin (50 nM) (n = 6) do not generate Sox2+ cells (q), Sox3+ cells (r) or Otx 2+ cells (s) but generate Msx1/2+ cells (50 % ± 30 % cells/section; n = 6 sections; [ t] ). Under these conditions no GATA2+ cells (u) cells are generated which may reflect previous findings that FGF can pattern the epidermal ectoderm, resulting in down-regulation of GATA2.
(v-z) Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium (0.3 x) (n = 6) do not generate Sox2+ cells (v), Sox3+ cells (w) or Otx 2+ cells (x) but generate Msx1/2+ cells (50 % ± 20 % cells/section; n = 6 sections; [ y] ) and GATA2+ cells (20 % ± 20 % cells/section; n = 6 sections; [ z] ).
(a'-e') Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium (0.3 x) and FGF (30 nM) generate Sox2+ cells (a'), Sox3+ cells (b') and Otx2+ cells (c') (90 % ± 20 % cells/section; n = 6 sections) but do not generate Msx1/2+ (d') or GATA2+ (e').
(f'-j') Stage X (L) explants cultured for 40h in the presence of mFrz8CRD-IgG conditioned medium (0.3 x) and Noggin (50 nM) generate Sox2+ cells (f'), Sox3+ cells (g') and Otx2+ cells (h') (70 % ± 20 % cells/section; n = 6 sections) but do not generate Msx1/2+ (i') or GATA2+ (j').
Scale bar shown in j'=100 µ m and applies to a-j'.
References
-
1.
New, D.A. A New Technique for the Cultivation of the Chick Embryo in vitro. J Embryol Exp Morphol 3, 320-331 (1955).
-
2.
Hamburger, V. & Hamilton, H.L. A series of normal stages in the development of the chick embryo. J Morph 88, 49-92 (1951).
-
3.
Wilson, S.I., Graziano, E., Harland, R., Jessell, T.M. & Edlund, T. An early requirement for FGF signalling in the acquisition of neural cell fate in the chick embryo. Curr Biol 10, 421-429 (2000).
-
4.
Read, E.M. et al. Evidence for non-axial A/P patterning in the nonneural ectoderm of Xenopus and zebrafish pregastrula embryos. Int J Dev Biol 42, 763-774 (1998).
Rights and permissions
About this article
Cite this article
Wilson, S., Rydström, A., Trimborn, T. et al. The status of Wnt signalling regulates neural and epidermal fates in the chick embryo. Nature 411, 325–330 (2001). https://doi.org/10.1038/35077115
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/35077115
This article is cited by
-
Head and neck squamous cell carcinoma: a potential therapeutic target for the Wnt signaling pathway
European Journal of Plastic Surgery (2022)
-
Single cell transcriptomics of human epidermis identifies basal stem cell transition states
Nature Communications (2020)
-
Hindbrain induction and patterning during early vertebrate development
Cellular and Molecular Life Sciences (2019)
-
Cytoplasmic localization of GRHL3 upon epidermal differentiation triggers cell shape change for epithelial morphogenesis
Nature Communications (2018)
-
Time-Course Gene Expression Profiling Reveals a Novel Role of Non-Canonical WNT Signaling During Neural Induction
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}
{kind=link}