
Abstract
Study design: In vitro study on the effects of mycophenolate mofetil (MMF) on isolated human monocytes and endothelial cells.
Objectives: Haematogenous macrophages play an essential role in the development of secondary damage following spinal cord injury (SCI), and there is evidence that the use of immunosuppressants such as MMF can reduce monocyte invasion and neuronal damage.
Setting: University Hospital for Orthopaedic Surgery, Frankfurt am Main, Germany.
Methods: The effects of MMF on the adhesion of human monocytes to human umbilical vein endothelial cells (HUVEC), monocyte binding to immobilised E-selectin, and monocyte expression of intercellular adhesion molecule (ICAM)-1, sialyl Lewis X (sLeX) and major histocompatibility complex (MHC)-II were studied. The binding of monocytes to E-selectin was examined by using purified and immobilised E-selectin fusion protein. Adhesion molecule expression was investigated by flow cytometry.
Results: The binding of monocytes to HUVEC was significantly reduced by 30.1% after treatment of monocytes with MMF (10 μg/ml), whereas the pretreatment of HUVEC with MMF did not result in significant changes in monocyte adhesion. MMF forcefully inhibited monocyte binding to immobilised E-selectin by 55.7%. Furthermore, MMF significantly inhibited the upregulation of ICAM-1- and MHC-II-expression on monocytes stimulated with either lipopolysaccharide or interferon-γ, whereas the expression of sLeX was not impaired. Toxic effects were excluded by propidium-iodide staining and measurement of fluorescein-diacetate metabolism.
Conclusion: MMF can downregulate important monocytic adhesion molecules and inhibits monocyte adhesion to endothelial cells, thus indicating that treatment with MMF could be beneficial after SCI.
Sponsorship: This study was supported by the DFG (Ha 2721/1-3), the Paul und Ursula Klein-Stiftung and the Stiftung Friedrichsheim.
Similar content being viewed by others

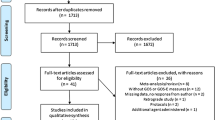
Introduction
The pathophysiology of spinal cord injury (SCI) is divided into two separate phases: during the phase of primary damage, compression of the spinal cord results in local tissue destruction with an immediate loss of neurons and axons; however, the extent of the initial lesion is dramatically enlarged in a process called secondary damage. Different pathophysiological mechanisms are thought to be responsible for this secondary damage: most importantly, resident microglial cells and haematogenous macrophages are recruited and activated, resulting in the release of neurotoxic substances such as glutamate, free radicals and nitric oxide (NO). Further neurotoxic effects are mediated by neuropetides and monoamines.1,2,3,4 Consequently, neurons and axons that were initially spared are irreversibly damaged in the hours and days following the traumatic insult.
The migration of monocytes from the blood stream into areas of tissue damage represents a major source of potentially neurotoxic agents: the proinflammatory cytokines tumour necrosis factor (TNF)-α, interleukin (IL)-1, IL-6; furthermore, NO and proteases are secreted by these haematogenous macrophages.5,6 Cytokines and chemotactic stimuli enhance the recruitment of further inflammatory cells to the site of neuronal damage. Recent studies have shown that activated monocytes release the excitotoxic substance quinolinic acid,7,8 and there is also evidence that infiltrating monocytes are involved in the production of chondroitin sulphate proteoglycans that are known to inhibit axonal outgrowth.9,10 Taken together, monocyte recruitment and activation contribute to secondary damage of neurons and a delayed loss of axons.
The recruitment of monocytes through the damaged blood–brain barrier is known to be mediated by adhesion molecules. Initially, leucocytes establish a transient adhesion via the selectin family of adhesion molecules, also called ‘tethering’, mediated by E- and P-selectin. Firm adhesion is established by enhanced ligand affinity of the leucocyte integrins such as leucocyte function-associated antigen (LFA)-1 and very late antigen (VLA)-4. Consequently, these integrins bind to their corresponding ligands intercellular cell adhesion molecule (ICAM)-1 and vascular cell adhesion molecule (VCAM)-1, both belonging to the immunoglobulin superfamily. Finally, transendothelial migration of adherent leucocytes into the damaged tissue occurs.11,12,13,14
Interference with different steps of the adhesion cascade seems to result in reduced secondary damage and improved functional outcome in animal models of SCI: Hamada et al showed that ICAM-1 mRNA expression correlates with the severity of the injury in a rat model of SCI; furthermore, animals treated with monoclonal antibodies directed against ICAM-1 showed a significantly better recovery and reduced oedema at the lesion site.15,16 Farooque et al17 used double knockout mice lacking both ICAM-1 and P-selectin and showed improved locomotor function after SCI in double knockout mice when compared to lesioned wild-type animals.
Consequently, a possible therapeutic strategy following SCI should consist of pharmacological immunosuppression and antiadhesive treatment to prevent monocyte recruitment. This goal could be achieved with the immunosuppressive drug mycophenolate mofetil (MMF), an inhibitor of the inosine monophosphate dehydrogenase that blocks de novo synthesis of guanine nucleotides. The depletion of guanosine triphosphate (GTP) and deoxyguanosine triphosphate (dGTP) pools inhibits the transfer of fucose and mannose to glycoproteins, some of which are adhesion molecules.18,19,20 Thus, by modulating the gycosylation of adhesion molecules, MMF could reduce the recruitment of monocytes and thereby reduce the amount of secondary damage after SCI.
It was the aim of our study to investigate whether MMF can interfere with different steps of the adhesion cascade of monocytes to endothelial cells. We therefore studied the adhesion of monocytes to human umbilical endothelial vein endothelial cells (HUVEC), and the effects of MMF treatment on the adhesion of monocytes to purified E-selectin as an important ligand for monocytic adhesion receptors. Furthermore, we assessed the monocytic expression of sialyl Lewis X (sLeX), ICAM-1 and major histocompatibility complex (MHC)-II.
Materials and methods
Isolation and cell cultures of human monocytes
Monocytes were isolated from venous blood of healthy human donors. The blood was diluted 1:1 with phosphate-buffered saline (PBS; Ca2+/Mg2+-free, Gibco) and then centrifuged in Leuco-Sep tubes (Greiner, Frickenhausen, Germany) including Ficoll separating solution (Seromed Biochrom, Berlin, Germany; density 1.077 g/ml) at 600 g for 30 min. The resulting layer of peripheral blood mononuclear cells was then removed and washed twice with PBS. After centrifugation, cells were resuspended in RPMI medium (Gibco) containing 50% fetal calf serum (FCS) (Gibco, Karlsruhe, Germany), 2% 1 M HEPES buffer (Seromed, Berlin, Germany), 100 U/ml penicillin and 0.1 mg/ml streptomycin (Sigma-Aldrich, Irvine, UK), placed in six-well plates (Falcon) and incubated for 2 h (37°C, 5% CO2, 100% humidified atmosphere). After this preplating for 2 h, the supernatant was removed and plates were washed with RPMI medium (37°C) until nonadherent cells were removed from the plates. Adherent monocytes were incubated in RPMI medium containing 10% FCS, 2% 1 M HEPES buffer, 100 U/ml penicillin and 0.1 mg/ml streptomycin. The purity of monocyte cultures was assessed by flow cytometry using double labelling with anti-CD14-PE and anti-CD45-FITC and was found to be >90%.
Isolation of endothelial cells
Endothelial cells were derived from human umbilical veins (HUVEC). Briefly, a cannula was inserted into the umbilical vein, fixed with a clamp and the vein perfused with Ca2+/Mg2+-free PBS (Gibco). The umbilical vein was then loaded with 2–5 ml α-chymotrase (Strathmann AG, Hamburg, Germany) and flushed after 10 min of incubation with 50 ml Medium 199 (Biozol, Munich, Germany). The cell suspension was centrifuged at 300 g for 10 min, resuspended and cells were seeded into culture flasks (Falcon Primaria). HUVEC were cultured under standard conditions (see above) in Medium 199 containing 10% FCS (Gibco, Karlsruhe, Germany), 10% pooled human serum (Blood Bank of the Red Cross, Frankfurt, Germany), 20 μg/ml endothelial cell growth factor (Boehringer, Mannheim, Germany), 10 IU/ml heparin (Roche, Basel, Switzerland), 100 μg/ml gentamycin (Gibco) and 2% 1 M HEPES buffer (Seromed, Berlin, Germany). The cells were grown until reaching confluency and only the first two passages of HUVEC were utilised for experiments.
Adhesion to endothelial cells
HUVEC were seeded into 12-well plates (Falcon), cultured until reaching confluency and incubated for 48 h with MMF in different concentrations (1 or 10 μg/ml). Monocytes were also incubated with MMF for 48 h with the same MMF concentrations. In control experiments, HUVEC and monocytes were incubated for 48 h without MMF treatment. After incubation, HUVEC were washed twice with Ca2+/Mg2+-free PBS and 1 ml Medium 199 was added to the cells. Monocytes were removed from six-well plates after 48 h incubation and suspended in RPMI medium at 1 × 106 cells/ml. From this cell suspension, 1 ml was added to HUVEC, resulting in a concentration of 0.5 × 106 monocytes/well. The following paradigms were carried out: (1) monocytes pretreated with MMF cultured with untreated HUVEC, (2) untreated monocytes cultured with HUVEC pretreated with MMF, (3) monocytes and HUVEC pretreated with MMF, (4) HUVEC and monocytes not pretreated with MMF cultured together (control). After incubation for 1 h, the cocultures were carefully washed with warm medium until nonadherent monocytes were removed, and cocultures were fixed with 4% paraformaldehyde (Merck, Darmstadt, Germany). Adherent monocytes were stained as explained below (see Immunocytochemistry). Stained cells were counted in five different fields using bright field microscopy (× 20). Mean adhesion was calculated as follows:

Adhesion spot assay
Isolated adhesion proteins were constructed as previously described (Figure 1).21,22,23 Round culture dishes (Becton Dickinson) were incubated with a spot of goat-anti-IgG (Sigma, 10 mg/ml in 50 mM Tris, pH 9.5, 90 min, 20°C). Dishes were then carefully washed with saline solution (150 mM NaCl) and subsequently incubated overnight with a blocking solution (PBS with Ca2+/Mg2+ and 1% bovine serum albumin, (BSA, Merck). The blocking solution was then removed, the culture dishes incubated with 5 μg/ml E-selectin IgG fusion protein (30 min, 20°C) and washed with saline solution (150 mM NaCl, 2 mM CaCl2). Monocytes were either not treated, stimulated with phorbole-myristate-acetate (PMA), or incubated with MMF (0.1, 1, 10 or 100 μg/ml) for 48 h. The cells were then removed from the six-well plates. After centrifugation, cells were resuspended in binding buffer (50 mM HEPES pH 7.4, 100 mM NaCl, 1 mg/ml BSA, 2 mM MgCl2, 1 mM CaCl2, 3 mM MnCl2, 0.02% NaN3 and 0.2 mM phenylmethylsulphonylfluoride) at a density of 1 × 106 cells/ml, 500 μl of this suspension was added to the prepared culture dishes (0.5 × 106 cells/dish) and incubated for 30 min (37°C). Nonadherent monocytes were carefully washed off with binding buffer and adherent cells counted in five different fields using bright field microscopy (× 20). Percentage of adhesion was calculated as follows:


The fusion protein human E-selectin/Fc was produced by Cos-7 cells. These cells were transfected with the DNA plasmid encoding human E-selectin and the produced fusion protein was isolated from the supernatant. This fusion protein was immobilised in Petri dishes pretreated with anti-FC-antibodies and isolated human monocytes were subsequently added to the dishes
Flow cytometry
Monocytes were stimulated with LPS (Sigma-Aldrich; 10 ng/ml), human recombinant interferon(IFN)-γ (PeproTech, London, UK; 500 U/ml) or human recombinant II-1α (R&D Systems, Wiesbaden, Germany; 100 U/ml) and simultaneously treated with MMF (0.1, 1 or 10 μg/ml) for 48 h. Monocytes were then removed from the wells, pelleted by centrifugation at 300 g for 5 min and stained with the following antibodies for 30 min (4°C): anti-CD54 (anti-ICAM-1)-PE (clone 15.2; Dianova)+anti-CD14-FITC (clone UCHM-1; Dianova); anti-HLA-DR (anti-MHC-II)-PE (clone DDII; Dianova)+anti-CD14-FITC (clone UCHM-1; Dianova) or anti-CD15s (anti-sLeX)-PE (clone 2H5; Pharmingen)+anti-CD14-FITC (clone UCHM-1; Dianova). The monocytes were washed with a buffer containing 0.5% BSA and their mean fluorescence intensity (MFI) was measured using a FACscan (Becton Dickinson, Heidelberg, Germany; 5 × 103 cells/scan). Only CD14+ monocytes were gated and the MFI of all gated cells was detected. MFI was calculated as follows:

Immunocytochemistry
Following fixation and washing with PBS, cells were treated (10 min) with methanol including 1.5% H2O2 (30%, Merck, Darmstadt, Germany). Cells were then washed again with PBS and pretreated with normal horse serum diluted 1:20 in PBS (0.3% Triton, Merck, Darmstadt, Germany) to minimise unspecific binding. Cells were then incubated overnight (20°C) with a primary antibody directed against Mac-1 (mouse-anti-human CD11b IgG1, clone ICRF 44, Serotec, Oxford, UK). The antibody was diluted 1:25 in PBS (0.3% Triton, 1% BSA). The cells were washed again with PBS and subsequently incubated for 60 min with the secondary antibody, anti-mouse IgG conjugated with biotin (Sigma-Aldrich, Steinheim, Germany, diluted 1:100 with PBS/Triton X-100). After washing with PBS, the cells were incubated for 60 min with ExtrAvidin (Sigma-Aldrich, Steinheim, Germany, diluted 1:100 in PBS/Triton X-100). Following washing with PBS and 0.05 M Tris-buffer, the chromogene 3,3′-diaminobenzidine tetrahydrochloride (DAB; Sigma-Aldrich, Steinheim, Germany, 5 μg/ml dissolved in 0.05 M Tris) was added for 2–5 min. The cells were rinsed again with 0.05 M Tris and embedded in fluid Kaysers glycerol gelatine (Merck, Darmstadt, Germany). Wells were then evaluated as described above.
MMF cytotoxicity assays
The cytotoxicity of MMF was evaluated by two different assays using fluorescein-diacetate (FDA) or propidium-iodide (PI). FDA is incorporated by intact cells and metabolised by intracellular esterases. Thus, quantitative fluorescence analysis of metabolised FDA correlates with the intracellular esterase activity in undamaged cells and thereby indicates the viability of the investigated monocytes. Monocytes were incubated with MMF (1, 10 or 100 μg/ml) and simultaneously stimulated with LPS (10 ng/ml). Untreated cells and cells destroyed by methanol treatment (5 min) were used as positive and negative controls. FDA was diluted in RPMI medium (1 μg/ml), added to the cells for 1 h (37°C) and the cells were subsequently pelleted (300 g, 5 min) and resuspended in PBS. Their MFI was measured using the a FACscan (Becton Dickinson, Heidelberg, Germany, FITC-filter, 1 × 104 cells/scan). Viability was calculated as follows:

PI penetrates cell membranes of lethally or sublethally damaged cells and intercalates with the DNA. Quantitative fluorescence analysis of PI correlates with the number of damaged cells. To assess cytotoxity, monocytes were treated with MMF (1, 10, or 100 μg/ml) alone, or simultaneously stimulated with LPS (10 ng/ml). After 48 h, cells were removed from the wells, centrifuged (300 × g, 5 min) and placed on ice. Cells were then resuspended with 500 μl RPMI medium containing PI (2.5 μg/ml) and assessment of MFI (1 × 104 cells/scan) was carried out immediately. The percentage of monocytes not labelled by PI was determined, indicating the proportion of viable, undamaged cells. The viability was evaluated as follows: viability (%)=percentage of gated monocytes with MFI<101 in FL1-H.
HPLC assay
The concentrations of mycophenolic acid (MPA) in the cell culture supernatant were determined after exposure of HUVEC to MMF (1, 10 or 100 μg/ml) for 3 h by reversed-phase HPLC.
Statistics
All experiments were carried out between three and 11 times. Numbers are given as mean±SEM. Statistical significance was investigated using the Wilcoxon-matched-pairs test. Results with a P-value of less than 0.05 were considered statistically significant.
Results
MMF inhibits monocyte adhesion to HUVEC
Incubation of monocytes with MMF for 48 h resulted in a significant reduction of their adhesion to HUVEC (Figure 2). The incubation with MMF (1 μg/ml) reduced monocyte adhesion by 26.2% (1 μg/ml MMF versus control: P<0.01) when compared to control experiments with untreated monocytes adhering to untreated HUVEC. Treatment of monocytes with 10 μg/ml MMF resulted in a 30.1% reduction of monocyte adhesion to HUVEC (10 μg/ml MMF versus control: P<0.01). In contrast, the preincubation of HUVEC with MMF had no significant effect on the adhesion of untreated monocytes. The adhesion of monocytes to HUVEC pretreated with either 1 or 10 μg/ml MMF was not significantly altered when compared to controls. Further experiments were carried out to assess whether combined treatment of monocytes and HUVEC with different concentrations of MMF induced antiadhesive effects, revealing that there was no additive synergism (data not shown).

Influence of MMF on the adhesion of monocytes to HUVEC. Monocytes or HUVEC were pretreated with different concentrations of MMF for 48 h and adhesion of monocytes to HUVEC was analysed. Pretreatment of monocytes significantly decreased their binding to HUVEC; however, adhesion of monocytes was not significantly influenced when HUVEC were pretreated with MMF (**P<0.01)
MMF inhibits monocyte adhesion to purified E-selectin
The adhesion spot experiments showed that untreated monocytes adhered strongly to E-selectin, whereas the incubation of monocytes with MMF significantly reduced their adhesion to immobilised E-selectin (Figure 3a, b). Treatment of monocytes with 0.1 μg/ml MMF for 48 h resulted in a moderate and statistically not significant reduction by 8.2%, whereas higher concentrations of MMF significantly reduced the monocyte adhesion to immobilised E-selectin: Incubation of monocytes with MMF reduced the binding by 27.8% (1 μg/ml MMF versus control: P<0.05), by 55.7% (10 μg/ml MMF versus control: P<0.001) or by 72.2% (100 μg/ml MMF versus control: P<0.001), demonstrating a dose-dependent inhibition of monocyte adhesion following treatment with MMF.

(a) Influence of MMF on monocyte adhesion to purified E-selectin. The spot adhesion assay revealed that pretreatment of monocytes with MMF resulted in reduced adhesion to purified E-selectin. The reduction of monocyte adhesion after pretreatment with different concentrations of MMF appeared to be dose dependent (*P<0.05, ***P<0.001). (b) Adherent monocytes in the spot adhesion assay (bright field microscopy, × 20). The high number of monocytes showing specific adhesion to purified E-selectin (*) can easily be distinguished from the lower number of monocytes that bind unspecifically to the areas surrounding the spot (**)
MMF reduces monocyte ligand expression
To study the effects of MMF on the monocytic expression of different adhesion molecules, monocytes, were stimulated with IFN-γ (500 U/ml) or LPS (10 ng/ml) for 48 h. Stimulation of monocytes with LPS resulted in a significant 2.73-fold increase in ICAM-1 immunoreactivity (LPS versus control: P<0.001), whereas IFN-γ induced a significant 2.14-fold increase in MHC-II immunoreactivity (IFN-γ versus control: P<0.001); (Figures 4 and 5). Simultaneous treatment with LPS and MMF (10 μg/ml) inhibited ICAM-1 upregulation by 18.9% (LPS vs [LPS+MMF 10 μg/ml]: P<0.001), whereas treatment of monocytes with 10 μg/ml MMF alone induced no significant changes in ICAM-1 immunoreactivity (Figure 4). The influences of higher MMF concentrations on the expression of ICAM-1 were not investigated. MHC-II expression was also influenced by MMF treatment, as MMF (1 or 10 μg/ml) significantly reduced MHC-II immunoreactivity on monocytes by 18.3% (IFN-γ versus (IFN-γ+MMF 1 μg/ml): P<0.001) and 15.7% (IFN-γ versus (IFN-γ+MMF 10 μg/ml): P<0.01), respectively (Figure 5). Surface immunoreactivity of sLeX on monocytes was not enhanced after stimulation with LPS. Futhermore, FACscan analysis revealed that sLeX immunoreactivity after simultaneous incubation with LPS and MMF (0.1, 1 or 10 μg/ml) was not significantly different from controls (Figure 6).

Expression of ICAM-1 on monocytes after treatment with MMF (10 μg/ml, 48 h) evaluated by FACscan analysis. The LPS-induced expression of ICAM-1 was significantly inhibited by treatment of monocytes with MMF (10 μg/ml). Treatment of unstimulated monocytes with MMF had no significant effect (***P<0.001)

Expression of MHC-II on monocytes after treatment with MMF evaluated by FACscan analysis. IFN-γ induced an increase in MHC-II expression that was reduced by simultaneous incubation with MMF (48 h; 1 or 10 μg/ml). Lower concentrations of MMF induced no significant effects (**P<0.01, ***P<0.001)

FACscan analysis of sLeX expression on monocytes after 48 h treatment with LPS revealed that sLeX was not induced by stimulation with LPS. The simultaneous treatment with MMF in different concentrations did not impair surface density of sLex
MPA-HPLC
Reverse-phase HPLC was performed to investigate whether HUVEC can metabolise MMF into MPA. The data revealed that the supernatant of HUVEC incubated with 100 μg/ml MMF for 3 h contained 23.1 μg/ml MPA. The incubation of HUVEC with lower concentrations of MMF (1 or 10 μg/ml) also resulted in the metabolisation of MMF into MPA (0.2 and 2.74 μg/ml, respectively). Thus, the ratio of MMF metabolised into MPA was similar for all analysed MMF concentrations (Figure 7).

Reverse HPLC was performed to measure the metabolisation of MMF into MPA by HUVEC. Results revealed that HUVEC can metabolise MMF into the active drug MPA. The ratio of MMF metabolised into MPA was similar for all analysed MMF concentrations
MMF is not toxic to monocytes
Two different assays were used to investigate whether treatment of monocytes with MMF has toxic effects on monocytes. The amount of fluorescence, as assessed by flow cytometry after incubation of monocytes with FDA, was similar in all investigated cells following incubation with control medium or after treatment with different concentrations of MMF (1, 10 or 100 μg/ml; Figure 8). In contrast, treatment with methanol resulted in an almost complete abrogation of intracellular esterase activity and a subsequent complete loss of fluorescence. Flow cytometry analysis of PI uptake of monocytes revealed that the viability of monocytes was not impaired by MMF treatment: the incubation with LPS (10 ng/ml), MMF (1, 10 or 100 μg/ml) or simultaneous treatment with both LPS and MMF did not induce an increase in the number of stained cells (Figure 9).

The metabolism of incorporated FDA was used to investigate whether treatment with MMF has toxic effects on monocytes. FACscan analysis revealed that FDA metabolism was similar in control medium and after treatment with LPS or LPS+MMF, respectively. In contrast, treatment with methanol resulted in an almost complete abrogation of intracellular esterase activity, resulting in loss of fluorescence

Incorporation of PI and subsequent FACscan analysis was used to investigate whether MMF alone or simultaneous treatment with LPS and MMF induces toxic effects and a subsequent increased uptake of PI. It was observed that the uptake of PI was similar in all monocytes, irrespective of whether cells were treated with MMF or not, thus, monocyte viability was not impaired by MMMF
Discussion
It was our aim to investigate the potential of MMF to alter the recruitment of monocytes into areas of tissue damage. Therefore, the adhesion of monocytes to HUVEC, the adhesion of monocytes to purified E-selectin and the monocytic expression of important adhesion molecules were investigated. Our results show that MMF treatment of monocytes significantly reduced their binding capacity to HUVEC as well as to purified E-selectin. Furthermore, the expression of molecules important for the establishment of cell–cell contacts such as ICAM-1 and MHC-II was reduced after treatment with MMF, whereas the surface density of the E-selectin ligand sLeX was not impaired by MMF.
Various lesion models of the spinal cord have revealed that there is a characteristic time course of infiltrating cells.24,25,26 In general, leucocyte migration into the spinal cord or CNS is severely restricted by the blood–brain barrier and is characterised by a low turn over. By allowing only small numbers of mononuclear cells to migrate across the blood–brain barrier, the highly specialised endothelium represents an efficient obstacle supporting the immune privileged state of the CNS.27,28,29 In contrast to these specific structural and functional characteristics, brain endothelial cells present a repertoire of cell adhesion molecules very similar to that of most other vessels in the human body.30
However, following SCI the invasion of leucocytes increases dramatically and contributes to the development of secondary damage.24,25,26 Monocytes play an essential role in these mechanisms.31,32,33,34,35 Their ability to secrete proinflammatory cytokines and various other potentially neurotoxic substances such as NO, proteases and quinolinic acid is a main reason for delayed neuronal destruction in the hours and days following SCI.3,4,5,6,7,8,36,37 Recent evidence has shown that cyclooxygenase-1 is rapidly expressed in microglia and macrophages after SCI and is thought to mediate early secondary injury.38 Consequently, several experiments have demonstrated that the pharmacological inhibition of monocyte infiltration into the lesioned spinal cord improved the functional outcome after SCI: 32,39 For instance, intravenous injection of liposome-encapsulated clodronate that induces depletion of peripheral monocytes resulted in an improved functional outcome in spinally injured rats.39
After SCI and in animal models such as experimental allergic encephalomyelitis (EAE), the adhesion molecules ICAM-1 and LFA-1 are expressed in the CNS and their expression coincides with the infiltration of inflammatory cells.40,41 Consequently, the treatment with anti-ICAM-1 monoclonal antibodies resulted in a reduction in the neurological deficit after SCI.15,16 The relevance of the selectins for recruitment of inflammatory cells towards the CNS is a matter of dispute. It has been shown that E-selectin and P-selectin are not involved in the development of EAE in a mouse model.42 In contrast, experiments on spinally injured knockout mice lacking ICAM-1 and P-selectin indicated that the functional outcome was better when compared to wild-type control animals.17 A murine model of focal cerebral ischaemia revealed that mice expressing the P-selectin gene showed significantly greater neutrophil accumulation in the ischaemic cortex compared to P-selectin-null mice.43 In other models of cerebral ischaemia and reperfusion injury, it has been shown that the expression of E-selectin and P-selectin was upregulated on brain microvessels located within the ischaemic lesion, indicating that the selectins facilitate the adhesion of inflammatory cells and subsequently increase neuronal injury.44,45
We hypothesised that the immunomodulatory drug MMF can influence the process of monocyte adhesion to endothelial cells, possibly by altering the glycosylation of adhesion molecules. It has been described that different adhesion molecules require glycosylation for the activation of their adhesive functions,18 consequently, Heemann et al46 showed that MMF treatment reduced leucocyte infiltration into transplanted kidneys.46 We found that preincubation of monocytes with MMF significantly inhibited their ability to adhere to HUVEC, which is in agreement with the findings of Laurent et al.47 Unexpectedly, the preincubation of HUVEC with MMF induced no significant changes in monocyte adhesion. More specifically, we found that the interaction of monocytes with E-selectin was inhibited in a dose-dependent manner in the presence of MMF. Taking into account that the monocytic immunoreactivity for sLeX, the appropiate monocyte ligand for E-selectin, was not altered after treatment with MMF, we suggest that conformational changes resulting in reduced affinity of sLeX for E-selectin could be responsible for the observed antiadhesive effects.
In vivo, MMF is known to be metabolised to MPA, which is the active drug. In our experiments, MMF was used because most in vivo studies have been performed with this prodrug. However, even in vitro MMF is rapidly hydrolysed to MPA, as has been shown in human peripheral blood mononuclear cells.18 Our own data on the metabolisation of MMF into MPA in HUVEC support this notion (Figure 7).
MPA is known for its relatively high affinity to human serum albumin and published data support the hypothesis that the pharmacological activity of MPA is a function of unbound drug concentration.48 Thus, it can be expected that the addition of fetal bovine serum or human serum to the culture media reduces the immunosuppressive capacity of MMF. This could be the reason for the relatively high concentrations of MMF that were needed to achieve immunosuppressive activity in vitro, as the culture medium used for human monocytes and endothelial cells contained 10% fetal bovine serum and 10% human serum.
A major question is whether MMF is able to access the site of SCI in the hours or days after SCI. Studies about the blood–brain-barrier-permeability of MMF have not yet been published. Nevertheless, it is known that a measurable leakage of the blood–brain barrier occurs as early as 1 h after the spinal cord lesion and that the size of the ‘leaky area’ increases in parallel with the size of the lesion site. In general, the size of the ‘leaky area’ is much larger than the lesion size. After 2 weeks, no more blood–brain-barrier-leakage can be detected in and around the lesion.24 It is known that other drugs with comparable physicochemical properties such as steroids can permeate the blood–brain barrier in the aftermath of SCI to fulfil their beneficial effects. Thus, we expect that MMF also permeates the blood–brain barrier after SCI.
It is important to note that the observed effects of MMF were not unspecifically toxic, as the treatment of monocytes with MMF induced no decrease in FDA metabolism or an increase in PI labelling. Our findings are in agreement with the results of other authors, who showed that concentrations up to 100 μM are not toxic to lymphocytes or EC.49,50
In summary, our results indicate that MMF has the capacity to reduce the interaction of monocytes with endothelial cells, thereby inhibiting the recruitment to sites of inflammation. These pharmacological antiadhesive and immunosuppressive effects could have beneficial effects in the treatment of acute SCI.
References
Choi DW . Cerebral hypoxia: some new approaches and unanswered questions. J Neurosci 1990; 10: 2493–2501.
Demopoulus HB, Flamm ES, Pietronigro DD, Seligman ML . The free radical pathology and the microcirculation in the major central nervous system disorders. Acta Physiol Scand 1980; 492: 91–119.
Liu D, Ling X, Wen J, Liu J . The role of reactive nitrogen species in secondary spinal cord injury: formation of nitric oxide, peroxynitrite, and nitrated protein. J Neurochem 2000; 75: 2144–2154.
Liu D, Thangnipon W, McAdoo DJ . Exciatory amino acids rise to toxic levels upon impact injury to the rat spinal cord. Brain Res 1991; 547: 344–348.
Cavaillon JM . Cytokines and macrophages. Biomed Pharmacother 1994; 48: 445–453.
van der Laan LJ et al. Macrophage phagocytosis of myelin in vitro determined by flow cytometry: phagocytosis is mediated by CR3 and induces production of tumor necrosis factor-alpha and nitric oxide. J Neuroimmunol 1996; 70: 145–152.
Blight AR, Cohen TI, Saito K, Heyes MP . Quinolinic acid accumulation and functional deficits following experimental spinal cord injury. Brain 1995; 118: 735–752.
Sinz EH et al. Quinolinic acid is increased in CSF and associated with mortality after traumatic brain injury in humans. J Cereb Blood Flow Metab 1998; 18: 610–615.
Davies SJ et al. Regeneration of adult axons in white matter tracts of the central nervous system. Nature 1997; 390: 680–683.
Fitch MT, Silver J . Activated macrophages and the blood brain barrier: inflammation after CNS injury leads to increases in putative inhibitory molecules. Exp Neurol 1997; 14: 587–603.
de Vries HE et al. Lymphocyte adhesion to brain capillary endothelial cells in vitro. J Neuroimmunal 1994; 52: 1–8.
Mabon PJ, Weaver LC, Dekaban GA . Inhibition of monocyte/macrophage migration to a spinal cord injury site by an antibody to the integrin alphaD: a potential new anti-inflammatory treatment. Exp Neurol 2000; 166: 52–64.
Stanimirovic DB, Wong J, Shapiro A, Durkin JP . Increase in surface expression of ICAM-1, VCAM-1 and E-selectin in human cerebromicrovascular endothelial cells subjected to ischemia-like insults. Acta Neurochir Suppl. (Wien) 1997; 70: 12–16.
Carlos TM et al. Expression of endothelial adhesion molecules and recruitment of neutrophils after traumatic brain injury in rats. J Leukoc Biol 1997; 61: 279–285.
Clark WM, Madden KP, Rothlein R, Zivin JA . Reduction of central nervous system ischemic injury by monoclonal antibody to intercellular adhesion molecule. J Neurosurg 1991; 75: 623–627.
Hamada Y et al. Involvement of an intercellular adhesion molecule 1-dependent pathway in the pathogenesis of secondary changes after spinal cord injury in rats. J Neurochem 1996; 66: 1525–1531.
Farooque M, Isaksson J, Olsson Y . Improved recovery after spinal cord trauma in ICAM-1 and P-selectin knockout mice. Neuroreport 1999; 10: 131–134.
Allison AC, Eugui EM . Immunosuppressive and other effects of mycophenolate acid and an ester prodrug mycophenolate mofetil. Immunol Rev 1993; 136: 5–28.
Allison AC, Eugui EM . Mycophenolate mofetil, a rationally designed immunosuppressive drug. Clin Transplant 1993; 7: 96–112.
Allison AC et al. Mycophenolic acid and brequinar, inhibitors of purine and pyrimidine synthesis, block the glycosylation of adhesion molecules. Transpl Proc 1993; 25: 67–70.
Walz G et al. Recognition by ELAM-1 of the sialyl Lewis X determinant on myeloid and tumor cells. Science 1990; 250: 1332–1335.
Wittig BM et al. α (1,3) Fucosyltransferase expression in E-selectin-mediated binding of gastrointestinal tumor cells. Int J Cancer 1996; 67: 80–85.
Zettlmeissel G et al. Expression and characterisation of human CD4: immunoglobulin fusion protein. DNA Cell Biol 1990; 9: 347–353.
Dusart I, Schwab ME . Secondary cell death and inflammatory reaction after dorsal hemisection of the rat spinal cord. Eur J Neurosci 1994; 6: 712–724.
Holmin S, Mathiesen T, Shetye J, Biberfeld P . Intracerebral inflammatory response to experimental contusion. Acta Neurochir (Wien) 1995; 132: 110–119.
Popovich PG, Wei P, Stokes BT . Cellular inflammatory response after spinal cord injury in Sprague–Dawley and lewis rats. J Comp Neur 1997; 377: 443–464.
Fabry Z, Raine CS, Hart MN . Nervous tissue as an immune compartment: the dialect of the immune response in the CNS. Immunol Today 1994; 15: 218–224.
Hauser SL et al. Immunohistochemical staining of normal human brain with monoclonal antibodies that identify lymphocytes, monocytes, and the la antigen. J Neuroimmunol 1983; 5: 197–205.
Oehmicken M . Inflammatory cells in the central nervous system: an integrated concept based on recent research in pathology, immunology, and forensic medicine. Prog Neuropathol 1983; 5: 277–335.
Navratil E et al. Expression of cell adhesion molecules by microvascular endothelial cells in the cortical and subcortical regions of the normal human brain: an immunohistochemical analysis. Neuropathol Appl Neurobiol 1997; 23: 68–80.
Blight AR . Delayed demyelination and macrophage invasion: a candidate for secondary cell damage in spinal cord injury. CNS Trauma 1985; 2: 299–315.
Blight AR . Effects of silica on the outcome from experimental spinal cord injury: implication of macrophages in secondary tissue damage. Neuroscience 1994; 60: 263–273.
Dijkstra CD, de Groot CJ, Huitinga I . The role of macrophages in demyelination. J Neuroimmunol 1992; 40: 183–188.
Giulian D et al. Reactive mononuclear phagocytes release neurotoxins after ischemic and traumatic injury to the central nervous system. J Neurosci Res 1993; 36: 681–693.
Giulian D, Robertson C . Inhibition of mononuclear phagocytes reduces ischemic injury in the spinal cord. Ann Neurol 1990; 27: 33–42.
Hamada Y et al. Roles of nitric oxide in compression injury of rat spinal cord. Free Radic Biol Med 1996; 20: 1–9.
Lee YB et al. Role of tumor necrosis factor-alpha in neuronal and glial apoptosis after spinal cord injury. Exp Neurol 2000; 166: 190–195.
Schwab JM, Brechtel K, Nguyen TD, Schluesener HJ . Persistent accumulation of cyclooxygenase-1 (COX-1) expressing microglia/macrophages and upregulation by endothelium following spinal cord injury. J Neuroimmunol 2000; 111: 122–130.
Popovich PG et al. Depletion of hematogenous macrophages promote partial hindlimb recovery and neuroanatomical repair after experimental spinal cord injury. Exp Neurol 1999; 158: 351–365.
Canella B, Cross AH, Raine CS . Upregulation and coexpression of adhesion molecules correlate with relapsing autoimmune demyelination in the central nervous system. J Exp Med 1990; 172: 1521–1524.
Isaksson J et al. Expression of ICAM-1 and CD11b after experimental spinal cord injury in rats. J Neurotrauma 1999; 16: 165–173.
Engelhardt B, Vestweber D, Hallmann R, Schulz M . E- and P-selectin are not involved in the recruitment of inflammatory cells across the blood–brain barrier in experimental autoimmune encephalomyelitis. Blood 1997; 90: 4459–4472.
Connolly Jr ES et al. Exacerbation of cerebral injury in mice that express the P-selectin gene: identification of P-selectin blockade as a new target for the treatment of stroke. Circ Res 1997; 81: 304–310.
Okada Y et al. P-selectin and intercellular adhesion molecule-1 expression after focal brain ischemia and reperfusion. Stroke 1994; 25: 202–211.
Zhang RL et al. E-selectin in focal cerebral ischemia and reperfusion in the rat. J Cereb Blood Flow Metab 1996; 16: 1126–1136.
Heeman U et al. Mycophenolate mofetil inhibits lymphocyte binding and the upregulation of adhesion molecules in acute rejection of rat kidney allografts. Transpl Immunol 1996; 4: 64–67.
Laurent AF, Dumont S, Poindron P, Muller CD . Mycophenolic acid suppresses protein N-linked glycosylation in human monocytes and their adhesion to endothelial cells and to some substrates. Exp Hemat 1996; 24: 59–67.
Nowak I, Shaw LM . Mycophenolic acid binding to human serum albumin: characterization and relation to pharmacodynamics. Clin Chem 1995; 41: 1011–1017.
Baer PC et al. Effects of mycophenolic acid on human renal proximal and distal tubular cells in vitro. Nephrol Dial Transplant 2000; 15: 184–190.
Blaheta RA et al. Inhibition of endothelial receptor expression and of T-cell ligand activity by mycophenolate mofetil. Transpl Immunol 1998; 6: 251–259.
Acknowledgements
We thank Ms R Schröder, Ms E Oppermann and Ms N Roser for their excellent technical assistance. This study was supported by the DFG (Ha 2721-/1-3), the Paul und Ursula Klein-Stiftung and the Stiftung Friedrichsheim.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Glomsda, B., Blaheta, R. & Hailer, N. Inhibition of monocyte/endothelial cell interactions and monocyte adhesion molecule expression by the immunosuppressant mycophenolate mofetil. Spinal Cord 41, 610–619 (2003). https://doi.org/10.1038/sj.sc.3101512
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.sc.3101512
Keywords
This article is cited by
-
Macrophage lipid accumulation in the presence of immunosuppressive drugs mycophenolate mofetil and cyclosporin A
Inflammation Research (2019)
-
Long-term outcome of mycophenolate mofetil treatment for patients with microscopic polyangiitis: an observational study in Chinese patients
Rheumatology International (2016)
-
In vitro effects of mycophenolic acid on survival, function, and gene expression of pancreatic beta-cells
Acta Diabetologica (2012)
