
Abstract
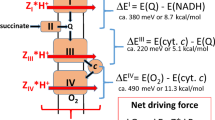
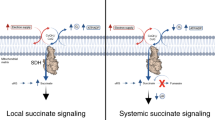
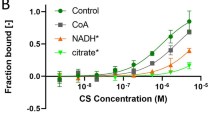
CALCIUM ions are known to inhibit oxidative phosphorylation in isolated mitochondria. As reported previously1, in the case of substrates oxidized via diphosphopyridine nucleotide, this inhibition can be reversed by the addition of the pyridine nucleotide, adenosine triphosphate and manganous ions. Treatment with the pyridine nucleotide alone partially restores only the respiration; but addition of all three factors is necessary in order to re-establish both oxidation and phosphorylation.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others
References
Lindberg, O., and Ernster, L., Nature, 173, 1038 (1954).
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
ERNSTER, L., LINDBERG, O. & LÖW, H. Reconstruction of Coupled Phosphorylation in the Succinic Oxidase System. Nature 175, 168–169 (1955). https://doi.org/10.1038/175168a0
Issue Date:
DOI: https://doi.org/10.1038/175168a0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
