
Abstract
Topological protection allows robust transport of localized phenomena such as quantum information, solitons and dislocations. The transport can be either dissipative or non-dissipative. Here, we experimentally demonstrate and theoretically explain the topologically protected dissipative motion of colloidal particles above a periodic hexagonal magnetic pattern. By driving the system with periodic modulation loops of an external and spatially homogeneous magnetic field, we achieve total control over the motion of diamagnetic and paramagnetic colloids. We can transport simultaneously and independently each type of colloid along any of the six crystallographic directions of the pattern via adiabatic or deterministic ratchet motion. Both types of motion are topologically protected. As an application, we implement an automatic topologically protected quality control of a chemical reaction between functionalized colloids. Our results are relevant to other systems with the same symmetry.
Similar content being viewed by others

Introduction
Topological invariants are global properties of a system that remain unchanged by local perturbations. A property that depends only on topological invariants is topologically protected and is very robust against local changes. Topological protection is a promising approach to stabilize quantum computing1 and is used to, for example, maintain robust transport in Hamiltonian systems. Topologically required edge states2 in a bulk system can support transport of quantum mechanical excitations3, classical mechanical solitons4, dislocations5 and gyroscopic waves6. When the edge states are located in a gap of the bulk excitation spectrum, they are protected against scattering into bulk states. Conservation of the Chern number, which is a topological invariant, makes the edge states robust against perturbative interactions. Topological insulators7, which are based on this concept, conduct at the surface but insulate in bulk. In driven Hamiltonian systems, additional invariants, such as the winding number8,9 around quasi energy bands, add to the topological variety of possible transport phenomena.
Transport of a collection of classical particles with different properties, such as size, mobility and so on, usually generates a diffuse broadening of the trajectories. Topological protection might be used to transport a broad distribution of particles without dispersion, despite their different properties. High precision multiparticle transport is an important ingredient in, for example, multifunctional lab-on-a-chip devices10,11.
Topological protection is also possible in driven dissipative (non-Hamiltonian) systems. The interplay between dissipation and topology has been studied in open quantum systems, see for example refs 12, 13 for details. In driven dissipative lattices14,15,16,17,18, transport typically involves the thermal ratchet effect19,20,21,22, that is, biased irreversible jumps between neighbouring potential wells. Complicated correlations between the noise23, disorder24 and many particle interactions25 cause a motion of astonishing complexity. The thermal ratchet mechanism is not robust when transporting simultaneously different types of particles. The complexity makes it hard to maintain control over the transport of one type of particles when adjusting the external drive to control the transport of another particle type.
Here, we show an example of topological protection in a driven dissipative colloidal system. We achieve predictable multiparticle transport of diamagnetic and paramagnetic colloids above a hexagonal magnetic lattice. Using periodic boundary conditions, we describe the unit cell of the lattice as a torus, which defines the action space in which the colloids move. We drive the colloids with periodic modulation loops of an external magnetic field. The direction of the external field defines our control parameter space. The topological correspondence between control and action space is nontrivial, and enables robust, topologically protected, colloidal transport along the lattice vectors. The topological invariant in action space is the set of the two winding numbers around the torus, in close analogy with driven quantum systems8,9. We demonstrate experimentally the robustness of the motion and implement a topologically protected quality control of a chemical reaction between functionalized colloids. We also develop a theoretical framework that fully describes the experimental findings. Our results apply to any hexagonal pattern.
Results
Colloidal model system
We use paramagnetic polystyrene core shell and solid polystyrene colloids of average diameters 2.8 and 3.1 μm, respectively, dispersed in a mixture of diluted water-based ferrofluid. The immersion of the colloids renormalizes their effective susceptibilities such that χp,eff>0 and χd,eff<0 for paramagnetic and diamagnetic colloids, respectively. The colloids immersed in the ferrofluid are placed on top of a magnetically patterned ferrite garnet film (FGF), see Fig. 1a. Spacer beads and a top glass plate create a ferrofluid film of thickness d=4.8 μm. Magnetic boundary conditions at the garnet-ferrofluid and glass-ferrofluid interfaces distort the magnetic field lines (created by the magnetic moments of the colloids) to be parallel to both interfaces. Virtual image dipoles form in the garnet film and the top glass plate and generate a potential that levitates the colloids into the mid-film plane, far away from the FGF, see Fig. 1b. Without the ferrofluid the colloids sediment to the pattern26,27. The FGF is characterized by a hexagonal lattice of magnetic bubble domains magnetized normal to the film (saturation magnetization Ms=17 kA m−1). The bubbles are immersed in a continuous phase of opposite magnetization. In an external field  normal to the film, the bubbles grow on the expense of the continuous phase if
normal to the film, the bubbles grow on the expense of the continuous phase if  >0 and shrink if
>0 and shrink if  <0.
<0.

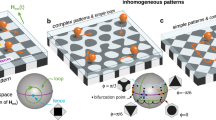
(a) Hexagonal garnet film with lattice constant a=11.6 μm covered with ferrofluid of thickness d=4.8 μm. One Wigner–Seitz unit cell is marked with a dashed line. By adjusting a closed modulation loop of a spatially homogeneous magnetic field Hext(t), we have total control over the transport of paramagnetic (blue) and diamagnetic (green) colloids immersed into the ferrofluid. (b) Lateral view of the system showing the distortion of the dipolar magnetic field (the field of the garnet pattern is omitted here) of an individual particle immersed in ferrofluid. The field distortion pushes the colloidal particle into the midplane of the ferrofluid film. (c) The direction of Hext varies on the surface of a sphere, defining control space  . Control space can be divided into three regions: the north, the tropics and the south. The northern and southern borders separate the tropics from the north and the south, respectively. Each border consists of 12 segments that we number from 0 to 11. The segments join at special points, indicated by empty circles and squares.
. Control space can be divided into three regions: the north, the tropics and the south. The northern and southern borders separate the tropics from the north and the south, respectively. Each border consists of 12 segments that we number from 0 to 11. The segments join at special points, indicated by empty circles and squares.  is an example of a closed modulation loop of Hext that induces transport of diamagnetic particles along the lattice. The loop crosses the northern border through segments 1 and 4.
is an example of a closed modulation loop of Hext that induces transport of diamagnetic particles along the lattice. The loop crosses the northern border through segments 1 and 4.
Control space
We use a homogeneous time-dependent magnetic external field Hext(t) of constant magnitude, Hext=5 kA m−1, to drive the system. Hence, our control space  is the surface of a sphere. Each point on
is the surface of a sphere. Each point on  corresponds to a direction of Hext. For reasons that will become clear later, we can divide
corresponds to a direction of Hext. For reasons that will become clear later, we can divide  in three regions: the north, the tropics and the south, see Fig. 1c. We call the interface between the tropics and the north (south) as the northern (southern) border. Each border is made of 12 segments. We experimentally perform periodic closed modulation loops
in three regions: the north, the tropics and the south, see Fig. 1c. We call the interface between the tropics and the north (south) as the northern (southern) border. Each border is made of 12 segments. We experimentally perform periodic closed modulation loops  of the external magnetic field. The period of the modulation is irrelevant provided that it is large enough such that the particles can follow the changes of the potential generated by Hext. There exist loops that induce intercellular colloidal transport. That is, when
of the external magnetic field. The period of the modulation is irrelevant provided that it is large enough such that the particles can follow the changes of the potential generated by Hext. There exist loops that induce intercellular colloidal transport. That is, when  returns to its initial point, the colloids are not in their initial positions but on a different unit cell.
returns to its initial point, the colloids are not in their initial positions but on a different unit cell.
Experimental phase diagram
Only loops that cross the northern (southern) border of  induce intercellular transport of the diamagnets (paramagnets). We discuss first the motion of the diamagnets. Let
induce intercellular transport of the diamagnets (paramagnets). We discuss first the motion of the diamagnets. Let  =(iN, jN) be a loop in
=(iN, jN) be a loop in  that crosses the ith segment of the northern border from the tropics to the north and returns to the tropics using the jth segment, see an example in Fig. 1c. The experimental phase diagram showing the motion of diamagnetic colloids for all possible modulation loops of type
that crosses the ith segment of the northern border from the tropics to the north and returns to the tropics using the jth segment, see an example in Fig. 1c. The experimental phase diagram showing the motion of diamagnetic colloids for all possible modulation loops of type  =(iN, jN) is shown in Fig. 2a. The precise shape of the loop is irrelevant, a clear sign of the robustness of the transport. Only the segments of the northern border crossed by
=(iN, jN) is shown in Fig. 2a. The precise shape of the loop is irrelevant, a clear sign of the robustness of the transport. Only the segments of the northern border crossed by  and their order is important. We can transport the diamagnets along the six fundamental lattice translations plus intracellular transport. Each direction is represented by a different colour in the phase diagram. The clustering of identical colours indicates the topological protection of the transport direction. A rotation of
and their order is important. We can transport the diamagnets along the six fundamental lattice translations plus intracellular transport. Each direction is represented by a different colour in the phase diagram. The clustering of identical colours indicates the topological protection of the transport direction. A rotation of  by π/3 around the polar axis, that is, from
by π/3 around the polar axis, that is, from  =(iN, jN) to
=(iN, jN) to  =(iN+2, jN+2), is equivalent to rotate the sample by −π/3, and hence changes the transport direction by π/3. Therefore, the sixfold symmetry of the pattern guarantees that if transport is possible along one direction then it must also be possible in the other five directions.
=(iN+2, jN+2), is equivalent to rotate the sample by −π/3, and hence changes the transport direction by π/3. Therefore, the sixfold symmetry of the pattern guarantees that if transport is possible along one direction then it must also be possible in the other five directions.

(a) Experimental phase diagram showing the direction and type of motion of the diamagnets for the fundamental loops  =(iN, jN) crossing the northern border in
=(iN, jN) crossing the northern border in  . The same diagram holds for the paramagnets if the modulation loops cross the southern border:
. The same diagram holds for the paramagnets if the modulation loops cross the southern border:  =(iS, jS). Each colour corresponds to a direction of motion, as indicated. Non-textured squares indicate adiabatic motion, and striped textured squares indicate ratchet motion. Empty circles mark the time reversal ratchets. (b) Polarization microscopy images of the pattern and the diamagnetic and paramagnetic colloidal particles at the end of a transport process. Scale bar (yellow rectangle middle image), 10 μm. The path of one paramagnet (blue arrow) and one diamagnet (green arrow) in
=(iS, jS). Each colour corresponds to a direction of motion, as indicated. Non-textured squares indicate adiabatic motion, and striped textured squares indicate ratchet motion. Empty circles mark the time reversal ratchets. (b) Polarization microscopy images of the pattern and the diamagnetic and paramagnetic colloidal particles at the end of a transport process. Scale bar (yellow rectangle middle image), 10 μm. The path of one paramagnet (blue arrow) and one diamagnet (green arrow) in  is depicted in the figure. The pink (cyan) segments of each path indicate the loop in
is depicted in the figure. The pink (cyan) segments of each path indicate the loop in  is on the southern (northern) hemisphere. The outer images show the transport of diamagnets into the x direction and paramagnets into one of the six crystallographic directions, by using modulation loops of type
is on the southern (northern) hemisphere. The outer images show the transport of diamagnets into the x direction and paramagnets into one of the six crystallographic directions, by using modulation loops of type  =(4N, 0N, iS, jS). The middle image is a Franconian folk dance performed by a paramagnetic and diamagnetic couple circulating around a central bubble in opposite sense and with different radius of the hexagon.
=(4N, 0N, iS, jS). The middle image is a Franconian folk dance performed by a paramagnetic and diamagnetic couple circulating around a central bubble in opposite sense and with different radius of the hexagon.
There are two types of motion, adiabatic and deterministic ratchet moves. The phase diagram is a checkerboard of alternating adiabatic- and ratchet-squares. In an adiabatic motion, the diamagnets always travel following the minimum generated by the magnetic potential. Hence, the speed of the modulation determines the speed of the colloids along the full trajectory. In contrast, the speed of the modulation loop does not fully determine the speed of the colloids in a ratchet. At some points during the modulation loop, the diamagnets hop between two minima of the magnetic potential at an intrinsic speed that is uncorrelated to the speed of the modulation.
The adiabatic motion is fully reversible. Reversing the modulation from  =(iN, jN) to
=(iN, jN) to  =(jN, iN) always reverts the direction of motion, and there is no hysteresis when comparing forward and backward trajectories of the colloids. For example, the loop
=(jN, iN) always reverts the direction of motion, and there is no hysteresis when comparing forward and backward trajectories of the colloids. For example, the loop  =(0N, 4N) transports the diamagnets adiabatically to the left, and the reverse loop
=(0N, 4N) transports the diamagnets adiabatically to the left, and the reverse loop  =(4N, 0N) to the right. In a deterministic ratchet motion, reversing the direction of the modulation loop does not usually revert the direction of the transported colloids.
=(4N, 0N) to the right. In a deterministic ratchet motion, reversing the direction of the modulation loop does not usually revert the direction of the transported colloids.  =(0N, 3N), for example, induces a ratchet transporting the diamagnets to the left, but the reverse loop
=(0N, 3N), for example, induces a ratchet transporting the diamagnets to the left, but the reverse loop  =(3N, 0N) does not transport the particles to the right. Only some of the modulation loops induce a time reversal ratchet in which reversing the modulation also reverts the direction of motion. See for example, the loops (0N, 6N) and (6N, 0N) in Fig. 2a. There is always hysteresis in ratchet-like motion between forward and backward trajectories, even in the case of time reversal ratchets.
=(3N, 0N) does not transport the particles to the right. Only some of the modulation loops induce a time reversal ratchet in which reversing the modulation also reverts the direction of motion. See for example, the loops (0N, 6N) and (6N, 0N) in Fig. 2a. There is always hysteresis in ratchet-like motion between forward and backward trajectories, even in the case of time reversal ratchets.
The dynamics we have discussed for the diamagnets on the northern border holds also for the paramagnets on the southern border of  . The phase diagram of the paramagnets is the same as the one of the diamagnets, cf Fig. 2a, if instead of modulation loops of type
. The phase diagram of the paramagnets is the same as the one of the diamagnets, cf Fig. 2a, if instead of modulation loops of type  =(iN, jN) we perform modulation loops of type
=(iN, jN) we perform modulation loops of type  =(iS, jS). That is, loops that cross the southern border of
=(iS, jS). That is, loops that cross the southern border of  from the tropics to the south using segment i and back to the tropics through segment j. An implicit equation to compute the location of the borders is given in the Methods section, and the exact location of the borders is shown in Supplementary Fig. 1.
from the tropics to the south using segment i and back to the tropics through segment j. An implicit equation to compute the location of the borders is given in the Methods section, and the exact location of the borders is shown in Supplementary Fig. 1.
The northern and southern borders of  are well separated. Hence, it is easy to transport the diamagnets and paramagnets successively by using a loop
are well separated. Hence, it is easy to transport the diamagnets and paramagnets successively by using a loop  =(iN, jN, kS, lS). The loop starts on the tropics and goes to the north of
=(iN, jN, kS, lS). The loop starts on the tropics and goes to the north of  crossing the segment iN, then returns to the tropics (jN) and moves to the south (kS). It finally returns to the starting point on the tropics of
crossing the segment iN, then returns to the tropics (jN) and moves to the south (kS). It finally returns to the starting point on the tropics of  crossing the segment lS. In Fig. 2b, we show polarization microscopy images of the combined transport of six representative modulation loops of the form
crossing the segment lS. In Fig. 2b, we show polarization microscopy images of the combined transport of six representative modulation loops of the form  =(4N, 0N, iS, jS). The loops induce adiabatic transport of diamagnets along the x-direction and adiabatic transport of paramagnets along the six possible lattice translations. The trajectories are coloured in pink (cyan) when
=(4N, 0N, iS, jS). The loops induce adiabatic transport of diamagnets along the x-direction and adiabatic transport of paramagnets along the six possible lattice translations. The trajectories are coloured in pink (cyan) when  travels on the northern (southern) hemisphere of
travels on the northern (southern) hemisphere of  .
.
We have total control over the colloidal motion, including the ability to programme complex trajectories. An example is given in the centre of Fig. 2b where we use a complex modulation loop such that the paramagnets and diamagnets perform a traditional Franconian folk dance. Videos showing the colloidal motion are provided in Supplementary Movies 1, 2, 3, 4, 5, 6, 7.
We next develop the theoretical framework needed to explain the experimental observations we have discussed above. An experimental application will be shown at the end of the Results section.
Action space
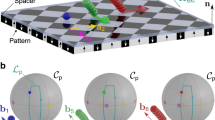
We call the space accessible to the colloids the action space  . Action space is a two-dimensional periodic hexagonal lattice at a fixed elevation above the FGF. Topologically
. Action space is a two-dimensional periodic hexagonal lattice at a fixed elevation above the FGF. Topologically  is a torus if we use periodic boundary conditions at the edges of a unit cell of the lattice, see Fig. 3a. Intercellular transport from one unit cell to the next cell via one of the two lattice vectors in real space is the same as one of the two windings around the torus. Loops
is a torus if we use periodic boundary conditions at the edges of a unit cell of the lattice, see Fig. 3a. Intercellular transport from one unit cell to the next cell via one of the two lattice vectors in real space is the same as one of the two windings around the torus. Loops  in
in  that correspond to intercellular transport of colloids have non-zero winding numbers, and cannot be continuously deformed into a point. That is, lattice translation action loops are non-zero-homotopic. This is not the case in control space. Any modulation loop
that correspond to intercellular transport of colloids have non-zero winding numbers, and cannot be continuously deformed into a point. That is, lattice translation action loops are non-zero-homotopic. This is not the case in control space. Any modulation loop  can be continuously deformed into any other desired modulation loop. For instance, we can continuously deform
can be continuously deformed into any other desired modulation loop. For instance, we can continuously deform  into a point on
into a point on  . Therefore, all loops in
. Therefore, all loops in  are zero-homotopic.
are zero-homotopic.

(a) Action space  is the space accessible to the colloids, a hexagonal lattice. Using periodic boundary conditions, action space is topologically a torus. (b) Two-dimensional projection of the stationary manifold
is the space accessible to the colloids, a hexagonal lattice. Using periodic boundary conditions, action space is topologically a torus. (b) Two-dimensional projection of the stationary manifold  , which has genus 7 and it is formed by 16 bijective areas indicated by different colours and listed in c.
, which has genus 7 and it is formed by 16 bijective areas indicated by different colours and listed in c.  ,
,  and
and  are the set of minima, saddle points and maxima of the magnetic potential, respectively. The fence
are the set of minima, saddle points and maxima of the magnetic potential, respectively. The fence  (
( ) separates
) separates  and
and  (
( ), and it is projected onto the northern (southern) border of control space, cf. Fig. 1c. In b, empty squares (circles) on
), and it is projected onto the northern (southern) border of control space, cf. Fig. 1c. In b, empty squares (circles) on  are triple plus
are triple plus  (zero
(zero  ) bifurcation points, at which 4 bijective areas meet. Three out of these bijective areas lie on
) bifurcation points, at which 4 bijective areas meet. Three out of these bijective areas lie on  (
( ) in a
) in a  (
( ) point.
) point.  (b) is an example of a non-zero-homotopic loop that winds around the holes of
(b) is an example of a non-zero-homotopic loop that winds around the holes of  . The corresponding control loop is
. The corresponding control loop is  =(1N, 4N). This loop in action space
=(1N, 4N). This loop in action space  induces intercellular transport of the diamagnets along the −x direction, black arrows in a. The colours in a show the projection of
induces intercellular transport of the diamagnets along the −x direction, black arrows in a. The colours in a show the projection of  and
and  onto action space.
onto action space.
Here, we have demonstrated that there exist modulation loops  in control space that induce either adiabatic or deterministic ratchet intercellular transport of the colloids. That is, there are zero-homotopic loops in
in control space that induce either adiabatic or deterministic ratchet intercellular transport of the colloids. That is, there are zero-homotopic loops in  that induce non-zero-homotopic action loops
that induce non-zero-homotopic action loops  with non-vanishing winding number around the torus. To understand how this is possible, we study theoretically the motion of point dipoles in the magnetic potential generated by the garnet and the external field.
with non-vanishing winding number around the torus. To understand how this is possible, we study theoretically the motion of point dipoles in the magnetic potential generated by the garnet and the external field.
Stationary manifold
The full dynamics is described by a point (Hext,  ) moving in the product phase space
) moving in the product phase space  , where
, where  is the position in action space. The energy landscape is given by the magnetic potential Vm=−χeffμ0H2, with H the total magnetic field and μ0 the vacuum permeability. H is the sum of the external field Hext∈
is the position in action space. The energy landscape is given by the magnetic potential Vm=−χeffμ0H2, with H the total magnetic field and μ0 the vacuum permeability. H is the sum of the external field Hext∈ and the internal field Hg(
and the internal field Hg( ) from the garnet film. The effective susceptibility χeff is positive for the paramagnets and negative for the diamagnets. Therefore, the unique scaled-potential V=H2 is enough to qualitatively describe the motion of both types of colloids. The stable points for the diamagnetic (paramagnetic) colloids are the minima (maxima) of V. The colloids are far away from the garnet film. Hence, we can approximate the potential by its leading non-constant term at large elevations, which is given by:
) from the garnet film. The effective susceptibility χeff is positive for the paramagnets and negative for the diamagnets. Therefore, the unique scaled-potential V=H2 is enough to qualitatively describe the motion of both types of colloids. The stable points for the diamagnetic (paramagnetic) colloids are the minima (maxima) of V. The colloids are far away from the garnet film. Hence, we can approximate the potential by its leading non-constant term at large elevations, which is given by:

where the sum runs only over the six reciprocal lattice vectors of the second Brillouin zone,  , all of which have magnitude q2. The full expression of V, at any elevation, is given in the Supplementary Note 1.
, all of which have magnitude q2. The full expression of V, at any elevation, is given in the Supplementary Note 1.  and
and  are the components of the external magnetic field in the ferrofluid parallel and normal to the garnet film, respectively. χ is the magnetic susceptibility of the ferrofluid. V is independent of the details of the FGF, and hence the following theory can be transferred to other systems with the same symmetry. For each value of Hext, the stationary points (Hext,
are the components of the external magnetic field in the ferrofluid parallel and normal to the garnet film, respectively. χ is the magnetic susceptibility of the ferrofluid. V is independent of the details of the FGF, and hence the following theory can be transferred to other systems with the same symmetry. For each value of Hext, the stationary points (Hext,  ) are those for which
) are those for which  , where
, where  indicates the gradient in action space. The set of these points forms the stationary manifold
indicates the gradient in action space. The set of these points forms the stationary manifold  , which is a two-dimensional manifold in
, which is a two-dimensional manifold in  . Only if
. Only if  contains non-zero-homotopic loops, we can achieve intercellular transport.
contains non-zero-homotopic loops, we can achieve intercellular transport.  can be viewed as the unification of three submanifolds:
can be viewed as the unification of three submanifolds:  . The Hessian matrix is positive definite in
. The Hessian matrix is positive definite in  (minima of V and hence stable points for the diamagnets), indefinite in
(minima of V and hence stable points for the diamagnets), indefinite in  (unstable saddle points for both colloids) and negative definite in
(unstable saddle points for both colloids) and negative definite in  (maxima of V and hence stable points for the paramagnets). One can show that
(maxima of V and hence stable points for the paramagnets). One can show that  has genus 7 with 3 holes in
has genus 7 with 3 holes in  and 2 holes in each,
and 2 holes in each,  and
and  , see Fig. 3b and Supplementary Fig. 2.
, see Fig. 3b and Supplementary Fig. 2.
Let  be the projection that maps any poinst in
be the projection that maps any poinst in  into control space. A key point is that
into control space. A key point is that  is multifold on
is multifold on  , that is, several points
, that is, several points  are mapped on the same point Hext∈
are mapped on the same point Hext∈ . The north, the south and the tropics of
. The north, the south and the tropics of  , cf. Fig. 1c, have different multiplicity of preimages on
, cf. Fig. 1c, have different multiplicity of preimages on  . The multiplicity changes at the borders of
. The multiplicity changes at the borders of  via generation or annihilation of pairs involving one saddle point and one minimum or one maximum. We can divide
via generation or annihilation of pairs involving one saddle point and one minimum or one maximum. We can divide  into a collection of bijective areas, {n+1, n+2, t+, s+}⊂
into a collection of bijective areas, {n+1, n+2, t+, s+}⊂ , {n01, n02, n03, t01, t02, s01, s02, s03}⊂
, {n01, n02, n03, t01, t02, s01, s02, s03}⊂ and {n−, t−, s−1, s−2}⊂
and {n−, t−, s−1, s−2}⊂ . Each area has exactly one preimage of either the north, the tropics or the south of
. Each area has exactly one preimage of either the north, the tropics or the south of  . The letter indicates if the area is projected onto the north (n), the tropics (t) or the south (s) of
. The letter indicates if the area is projected onto the north (n), the tropics (t) or the south (s) of  . These areas are listed in Fig. 3c (with the colours corresponding to the colouring of
. These areas are listed in Fig. 3c (with the colours corresponding to the colouring of  , Fig. 3b). The first subindex (0, +, −) indicates if the area lies on
, Fig. 3b). The first subindex (0, +, −) indicates if the area lies on  ,
,  or on
or on  . The second subindex labels the areas in case more than one area share the same letter and first subindex.
. The second subindex labels the areas in case more than one area share the same letter and first subindex.
Fences and bifurcation points
We call the boundary between  and
and  (
( ) as the northern
) as the northern  (southern
(southern  ) fence, see Fig. 3b. A segment of
) fence, see Fig. 3b. A segment of  separates a northern area on
separates a northern area on  (n+,1 or n+,2) from a northern area on
(n+,1 or n+,2) from a northern area on  (n0,1, n0,2 or n0,3) and starts and ends at vertices that are bifurcation points. Four different bijective areas in
(n0,1, n0,2 or n0,3) and starts and ends at vertices that are bifurcation points. Four different bijective areas in  meet at a bifurcation point, see Fig. 3b. There are three types of bifurcation points: triple zero bifurcation points
meet at a bifurcation point, see Fig. 3b. There are three types of bifurcation points: triple zero bifurcation points  , where three out of the four areas meeting at the point are on
, where three out of the four areas meeting at the point are on  , and triple plus
, and triple plus  (minus
(minus  ) bifurcation points, where three out of the four areas meeting at the point are on
) bifurcation points, where three out of the four areas meeting at the point are on  (
( ). In total, each fence has 12 bifurcation points that alternate between
). In total, each fence has 12 bifurcation points that alternate between  and
and  or
or  , depending on the fence. No further points where more than two areas meet on
, depending on the fence. No further points where more than two areas meet on  exist. The vertices on the fence are the only bifurcation points on
exist. The vertices on the fence are the only bifurcation points on  . The projection
. The projection  maps each of the 12 segments of
maps each of the 12 segments of  (
( ) onto one segment of the northern (southern) border of
) onto one segment of the northern (southern) border of  , see Fig. 1c.
, see Fig. 1c.  also maps the bifurcation points on
also maps the bifurcation points on  (
( ) onto 12 points at the northern (southern) border of
) onto 12 points at the northern (southern) border of  where two segments join. As
where two segments join. As  is multifold on
is multifold on  , the preimage of the borders of
, the preimage of the borders of  are the fences and other lines that we call the pseudo fences. The preimage of the projection of the bifurcation points are the bifurcation points and other points that we call pseudo bifurcation points. The pseudo fences separate different bijective areas on
are the fences and other lines that we call the pseudo fences. The preimage of the projection of the bifurcation points are the bifurcation points and other points that we call pseudo bifurcation points. The pseudo fences separate different bijective areas on  , and are also divided in 12 segments, which are separated by pseudo bifurcation points. We label the segments of the borders of
, and are also divided in 12 segments, which are separated by pseudo bifurcation points. We label the segments of the borders of  , and the segments of the fences and pseudo fences in
, and the segments of the fences and pseudo fences in  from 0 to 11. A segment i on
from 0 to 11. A segment i on  is projected onto the segment i on
is projected onto the segment i on  . Therefore, if we cross the ith segment of the border in
. Therefore, if we cross the ith segment of the border in  , we cross several ith segments of fences and pseudo fences on
, we cross several ith segments of fences and pseudo fences on  .
.
Adiabatic motion
We next explain the adiabatic transport of diamagnets, similar arguments apply for the paramagnets. To achieve adiabatic transport of diamagnets, we need a modulation loop  with a preimage loop
with a preimage loop  in
in  lying entirely in
lying entirely in  , such that the diamagnets can adiabatically follow the minimum of the magnetic potential. In addition,
, such that the diamagnets can adiabatically follow the minimum of the magnetic potential. In addition,  has to be non-zero-homotopic, that is, it has to wind around at least one of the two holes in
has to be non-zero-homotopic, that is, it has to wind around at least one of the two holes in  . This non-zero-homotopic loop is then projected onto a loop in
. This non-zero-homotopic loop is then projected onto a loop in  that can be non-zero-homotopic, and induce intercellular transport. As we have already shown adiabatic motion along any lattice direction, a=w1a1+w2a2, with ai the basic lattice vectors in
that can be non-zero-homotopic, and induce intercellular transport. As we have already shown adiabatic motion along any lattice direction, a=w1a1+w2a2, with ai the basic lattice vectors in  , is possible. Each transport direction corresponds to a value of the set of the two winding numbers {w1, w2} around the hole in
, is possible. Each transport direction corresponds to a value of the set of the two winding numbers {w1, w2} around the hole in  . Hence, our topological invariant is the set of winding numbers in
. Hence, our topological invariant is the set of winding numbers in  . In
. In  there are 7 holes, and hence 14 winding numbers. The sum of any winding number in
there are 7 holes, and hence 14 winding numbers. The sum of any winding number in  over all loops
over all loops  corresponding to a given loop in
corresponding to a given loop in  is zero since all loops in
is zero since all loops in  are zero-homotopic. We can only achieve a non-zero-homotopic loop in
are zero-homotopic. We can only achieve a non-zero-homotopic loop in  by first joining two zero-homotopic loops in
by first joining two zero-homotopic loops in  , and next disjoining them into two loops with opposite winding numbers. The detailed explanation is shown next.
, and next disjoining them into two loops with opposite winding numbers. The detailed explanation is shown next.
Consider the preimage in  of the modulation loop
of the modulation loop  =(1N, 1N). We show a schematic of
=(1N, 1N). We show a schematic of  and all its preimage loops in
and all its preimage loops in  in Fig. 4a. If
in Fig. 4a. If  is entirely in the tropics of
is entirely in the tropics of  (black loop) there are four zero-homotopic preimage loops on
(black loop) there are four zero-homotopic preimage loops on  . One is in
. One is in  , two in
, two in  and another one in
and another one in  . When
. When  touches the northern border of
touches the northern border of  (red loop), a pair of a minimum and a saddle point is generated at the fence
(red loop), a pair of a minimum and a saddle point is generated at the fence  . As
. As  crosses the northern border of
crosses the northern border of  (yellow loop), the minimum-saddle point pair deforms into a fifth (zero-homotopic) loop on
(yellow loop), the minimum-saddle point pair deforms into a fifth (zero-homotopic) loop on  that crosses the fence
that crosses the fence  . This new loop eventually disjoins into two new loops, one on
. This new loop eventually disjoins into two new loops, one on  and one on
and one on  , when
, when  fully enters the north of
fully enters the north of  (blue loop). At each stage in
(blue loop). At each stage in  , the other four loops on
, the other four loops on  smoothly pass through different pseudo fences on
smoothly pass through different pseudo fences on  . All loops on
. All loops on  produced with modulation loops
produced with modulation loops  =(iN, iN) are zero-homotopic and therefore do not produce transport in
=(iN, iN) are zero-homotopic and therefore do not produce transport in  . The specific bijective areas covered by the loops on
. The specific bijective areas covered by the loops on  depend on the segment of the border that we cross in
depend on the segment of the border that we cross in  . A figure showing the bijective areas that meet at each segment of fences and pseudo fences is given in Supplementary Fig. 3.
. A figure showing the bijective areas that meet at each segment of fences and pseudo fences is given in Supplementary Fig. 3.
 .
.
Schematic of different modulation loops  in control space
in control space  and their corresponding preimage loops on the stationary surface
and their corresponding preimage loops on the stationary surface  . (a)
. (a)  crosses the first segment of the northern border of
crosses the first segment of the northern border of  . When
. When  touches the border (red loop) a pair of a minimum and a saddle point is created in
touches the border (red loop) a pair of a minimum and a saddle point is created in  (red point). When
(red point). When  crosses the border twice (yellow loop), a loop crossing the fence in
crosses the border twice (yellow loop), a loop crossing the fence in  (fence-crossing loop) is created (yellow loop). This fence-crossing loop lies in both
(fence-crossing loop) is created (yellow loop). This fence-crossing loop lies in both  and
and  . (b) We enlarge
. (b) We enlarge  such that it encircles the projection of a triple plus bifurcation point. In
such that it encircles the projection of a triple plus bifurcation point. In  , the fence-crossing loop joins with the loop in
, the fence-crossing loop joins with the loop in  . No loop lies entirely in
. No loop lies entirely in  . (c)
. (c)  encircles the projection of two bifurcation points, one
encircles the projection of two bifurcation points, one  and one
and one  . The fence-crossing loop joins again with another loop that this time lies in
. The fence-crossing loop joins again with another loop that this time lies in  . (d)
. (d)  encircles now the projection of two
encircles now the projection of two  and one
and one  bifurcation points. The four areas meeting at the second
bifurcation points. The four areas meeting at the second  point (n+1, n+2, t+ and n02) were already joined in the fence-crossing loop. As a result, the fence-crossing loop disjoins into two loops, that in this case are non-zero homotopic with opposite winding numbers. One of the disjoint loops lies in
point (n+1, n+2, t+ and n02) were already joined in the fence-crossing loop. As a result, the fence-crossing loop disjoins into two loops, that in this case are non-zero homotopic with opposite winding numbers. One of the disjoint loops lies in  and induces intercellular adiabatic motion. All loops in a–c are zero-homotopic.
and induces intercellular adiabatic motion. All loops in a–c are zero-homotopic.
Let us now deform  such that it finally encircles the projection of a triple plus bifurcation point, see Fig. 4b. The final loop is
such that it finally encircles the projection of a triple plus bifurcation point, see Fig. 4b. The final loop is  =(1N, 2N). When
=(1N, 2N). When  crosses the projection of
crosses the projection of  , the corresponding loop
, the corresponding loop  crossing the fence on
crossing the fence on  joins with the pseudo fence-crossing loop on
joins with the pseudo fence-crossing loop on  . The result is a new loop that crosses the fence and passes through four areas on
. The result is a new loop that crosses the fence and passes through four areas on  . This loop lies in both
. This loop lies in both  and
and  . As no other loop entirely lies on
. As no other loop entirely lies on  , the diamagnets will follow a ratchet motion, leaving the stationary surface
, the diamagnets will follow a ratchet motion, leaving the stationary surface  when the loop crosses the fence towards
when the loop crosses the fence towards  . We will explain the ratchet motion later on. The winding number of the joint fence-crossing loop on
. We will explain the ratchet motion later on. The winding number of the joint fence-crossing loop on  is the sum of the winding numbers of the loops before the joining. In this case, the joining loops are zero-homotopic and hence the joint loop is also zero-homotopic and induces no transport in
is the sum of the winding numbers of the loops before the joining. In this case, the joining loops are zero-homotopic and hence the joint loop is also zero-homotopic and induces no transport in  .
.
In Fig. 4c, we further expand the modulation loop such that it encircles the following projection of a bifurcation point, a  . The final loop is
. The final loop is  =(1N, 3N). In
=(1N, 3N). In  , we again join the fence-crossing loop with a pseudo fence-crossing loop that now lies in
, we again join the fence-crossing loop with a pseudo fence-crossing loop that now lies in  . The result is, as in the previous case, a zero-homotopic fence-crossing loop.
. The result is, as in the previous case, a zero-homotopic fence-crossing loop.
We continue expanding the modulation loop such that it finally encircles the projection of two  points with
points with  =(1N, 4N), see Fig. 4d. Now, all four areas that meet at the second
=(1N, 4N), see Fig. 4d. Now, all four areas that meet at the second  bifurcation point in
bifurcation point in  are already joined in the fence-crossing loop. Therefore, crossing this bifurcation point disjoins the fence-crossing loops in two loops. The disjoint loops are no longer zero-homotopic. They have winding numbers with equal magnitude but opposite sign such that the sum is zero. One of the disjoint loops lies entirely in
are already joined in the fence-crossing loop. Therefore, crossing this bifurcation point disjoins the fence-crossing loops in two loops. The disjoint loops are no longer zero-homotopic. They have winding numbers with equal magnitude but opposite sign such that the sum is zero. One of the disjoint loops lies entirely in  , crosses the segments 1 and 4 of the pseudo fence between n+2 and t+ and winds around the holes in
, crosses the segments 1 and 4 of the pseudo fence between n+2 and t+ and winds around the holes in  . This loop is projected into a non-zero-homotopic loop in
. This loop is projected into a non-zero-homotopic loop in  that induces adiabatic transport of the diamagnets along the −x direction.
that induces adiabatic transport of the diamagnets along the −x direction.
Encircling the next projection of a  point,
point,  =(1N, 6N), joins again the loop in
=(1N, 6N), joins again the loop in  with a fence-crossing loop and creates a ratchet motion. The adiabatic transport is recovered by encircling a further projection of a
with a fence-crossing loop and creates a ratchet motion. The adiabatic transport is recovered by encircling a further projection of a  with
with  =(1N, 8N). This disjoins the fence-crossing loop and generates a new non-zero-homotopic loop in
=(1N, 8N). This disjoins the fence-crossing loop and generates a new non-zero-homotopic loop in  . This new loop crosses segments and pseudo fences in
. This new loop crosses segments and pseudo fences in  that are different than the previous non-zero-homotopic loop, and induces transport in a different lattice direction.
that are different than the previous non-zero-homotopic loop, and induces transport in a different lattice direction.
Deterministic ratchet motion
We next explain why the deterministic ratchet is topologically protected and its fundamental role in the phase diagram. A ratchet motion occurs if there is no loop that lies entirely on  . In this case, the minimum of the magnetic potential that transports the diamagnets disappears at the fence, and the particles leave the stationary manifold
. In this case, the minimum of the magnetic potential that transports the diamagnets disappears at the fence, and the particles leave the stationary manifold  jumping to another minimum.
jumping to another minimum.
Our modulation is adiabatic, that is, the relaxation time of the colloids in the cage around the minimum is orders of magnitude faster than the period of the modulation. Hence, if the diamagnets are on  , they follow the minimum of the potential with a dynamics given by the modulation. If, on the contrary, the diamagnets are not on
, they follow the minimum of the potential with a dynamics given by the modulation. If, on the contrary, the diamagnets are not on  , they move along the path of steepest descend of an effectively frozen potential in
, they move along the path of steepest descend of an effectively frozen potential in  . This path brings the diamagnets back to
. This path brings the diamagnets back to  .
.
Consider again the modulation loop  =(1N, 2N) that encircles the projection of a
=(1N, 2N) that encircles the projection of a  point and creates a ratchet. In Fig. 5a, we plot the loop in
point and creates a ratchet. In Fig. 5a, we plot the loop in  and the corresponding fence-crossing loop in
and the corresponding fence-crossing loop in  . We start
. We start  in the tropics of
in the tropics of  . In
. In  , the diamagnets follow the segment in t+ of the loop. Next,
, the diamagnets follow the segment in t+ of the loop. Next,  crosses the first segment of the northern border, and the diamagnets cross the first segment of the pseudo fence between t+ and n+2 in
crosses the first segment of the northern border, and the diamagnets cross the first segment of the pseudo fence between t+ and n+2 in  . Finally,
. Finally,  crosses the second segment of the border. At this point, the loop in
crosses the second segment of the border. At this point, the loop in  touches the fence. The minimum in n+2 that adiabatically transported the particles annihilates with a saddle point and disappears. The colloids leave the stationary surface
touches the fence. The minimum in n+2 that adiabatically transported the particles annihilates with a saddle point and disappears. The colloids leave the stationary surface  at the annihilation site. Diamagnets follow now the path of steepest descend and are brought back to
at the annihilation site. Diamagnets follow now the path of steepest descend and are brought back to  through the return site, see Fig. 5a. Hence, the fence-crossing loop in
through the return site, see Fig. 5a. Hence, the fence-crossing loop in  can be divided into accessible and inaccessible parts. The particles can stay only in the accessible part. The path of steepest descend connecting the annihilation and the return sites is topologically trivial. It cannot change the homotopy class of the adiabatic loop that emerges by taking the accessible part of the loop and gluing both ends, annihilation and return sites, together, see Fig. 5a. The reason is that the path of steepest descend develops in a continuous manner from the bifurcation point
can be divided into accessible and inaccessible parts. The particles can stay only in the accessible part. The path of steepest descend connecting the annihilation and the return sites is topologically trivial. It cannot change the homotopy class of the adiabatic loop that emerges by taking the accessible part of the loop and gluing both ends, annihilation and return sites, together, see Fig. 5a. The reason is that the path of steepest descend develops in a continuous manner from the bifurcation point  . To understand this, imagine we make
. To understand this, imagine we make  smaller and smaller but always encircling the projection of
smaller and smaller but always encircling the projection of  . Then, annihilation and return sites come closer and closer to each other, and eventually meet at the bifurcation point. This argument holds for any other ratchet motion in the system. A ratchet loop is always topologically protected by an adiabatic loop. Both loops have the same homotopy class, and therefore the same direction of motion.
. Then, annihilation and return sites come closer and closer to each other, and eventually meet at the bifurcation point. This argument holds for any other ratchet motion in the system. A ratchet loop is always topologically protected by an adiabatic loop. Both loops have the same homotopy class, and therefore the same direction of motion.

(a) A loop in  encircling the projection of a triple plus bifurcation point and the corresponding fence-crossing loop in
encircling the projection of a triple plus bifurcation point and the corresponding fence-crossing loop in  . (b) Same as in a but for a modulation loop in the opposite direction. The arrows in a,b indicate the starting points and the directions of the loops in
. (b) Same as in a but for a modulation loop in the opposite direction. The arrows in a,b indicate the starting points and the directions of the loops in  and
and  . The violet dashed loop is an adiabatic loop topologically equivalent to the deterministic ratchet loop. It is formed by gluing the annihilation and return sites of the ratchet loop. (c) Fraction of transported colloids f as a function of the magnetic susceptibility χ of the ferrofluid. f is computed by counting how many colloids out of 100 have been successfully transported after a modulation loop. The vertical dotted line approximately marks the transition between transport and non-transport phases.
. The violet dashed loop is an adiabatic loop topologically equivalent to the deterministic ratchet loop. It is formed by gluing the annihilation and return sites of the ratchet loop. (c) Fraction of transported colloids f as a function of the magnetic susceptibility χ of the ferrofluid. f is computed by counting how many colloids out of 100 have been successfully transported after a modulation loop. The vertical dotted line approximately marks the transition between transport and non-transport phases.
Let us now revert the direction of  , see Fig. 5b. The accessible part of the loop and the annihilation and return sites change. The forward and backward adiabatic loops that protect the ratchets are different, but induce transport in opposite directions. Therefore, the ratchet is time reversal. Reverting the modulation reverts the colloidal motion. There is, however, hysteresis since forward and backward loops differ in the path of steepest descend, and in the segments being crossed in
, see Fig. 5b. The accessible part of the loop and the annihilation and return sites change. The forward and backward adiabatic loops that protect the ratchets are different, but induce transport in opposite directions. Therefore, the ratchet is time reversal. Reverting the modulation reverts the colloidal motion. There is, however, hysteresis since forward and backward loops differ in the path of steepest descend, and in the segments being crossed in  . Usually, forward and backward loops are protected by adiabatic loops that induce transport in different, non-opposite, directions, resulting in a non-time reversal ratchet.
. Usually, forward and backward loops are protected by adiabatic loops that induce transport in different, non-opposite, directions, resulting in a non-time reversal ratchet.
Ratchets play a fundamental role in the system. The homotopy class of an adiabatic loop, which lies on  , cannot be changed by continuous deformations. Therefore, the direction of transport cannot change if the motion remains adiabatic (note that all neighbouring adiabatic loops in the phase diagram of Fig. 2a induce transport in the same direction). It is only via ratchets that we can change the homotopy class of a loop and hence the transport direction. See, for example, in Fig. 2a, the ratchet loop
, cannot be changed by continuous deformations. Therefore, the direction of transport cannot change if the motion remains adiabatic (note that all neighbouring adiabatic loops in the phase diagram of Fig. 2a induce transport in the same direction). It is only via ratchets that we can change the homotopy class of a loop and hence the transport direction. See, for example, in Fig. 2a, the ratchet loop  =(1N, 2N) (protected by the adiabatic loop (1N, 1N)) and the ratchet loop (1N, 3N) (protected by (1N, 4N)). The topological transition that changes the transport direction occurs when
=(1N, 2N) (protected by the adiabatic loop (1N, 1N)) and the ratchet loop (1N, 3N) (protected by (1N, 4N)). The topological transition that changes the transport direction occurs when  encircles the projection of a
encircles the projection of a  point (Supplementary Fig. 4).
point (Supplementary Fig. 4).
Theory and experiments are in perfect agreement. The above theory predicts exactly the same phase diagram we have found experimentally, cf Fig. 2a. In addition, we have also performed Brownian dynamic simulations of paramagnetic and diamagnetic particles moving in the potential given by equation (1). The simulations are also in perfect agreement with the theory and the experiments. The simulation allows us to introduce thermal noise in the system. We have verified that the topological protection is very robust against thermal fluctuations. When the noise is very high, such that it erases the energy landscape, the topological protection is lost. The degradation of the topological protection starts at both interfaces between different types of motion in the phase diagram: adiabatic-ratchet interface and the interface between deterministic ratchets along different directions.
The transport direction is also robust against other perturbations, such as the precise shape and speed of the modulation loop, changes in size, mobility and magnetic susceptibility of the colloids, and changes in the pattern that do not affect its symmetry (for example, the shape of the bubbles). Most strikingly, the directions of the ratchets are protected, that is, the topology of the stationary surface determines not only the direction of the adiabatic motion but also of the non-equilibrium ratchet motion.
There are always operations that break the topological protection. In our system, we can break the protection by changing the topology of  as we describe next.
as we describe next.
Elevation above the garnet
We return now to the experiments. We have discussed the transport of colloids at elevations z far away from the garnet film so that the potential is given by equation (1). At low elevations, the field created by the garnet is very strong compared with the external magnetic field and the potential is given by that of the pattern alone. In this situation, the different parts of  are disconnected manifolds and have a trivial topology (spheres) missing the requirements for topological transport.
are disconnected manifolds and have a trivial topology (spheres) missing the requirements for topological transport.
Depending on the dilution of the ferrofluid, the image-dipole potential may or may not overcome the gravitational potential. Hence, controlling the ferrofluid susceptibility gives direct control over the colloidal elevation z above the garnet film. Consider a loop in  that induces lattice translations if the colloids are at high elevations. By performing the same loop for different ferrofluid-water compositions, and hence varying the susceptibility χ, we can observe the transition from non-zero-homotopic loops in
that induces lattice translations if the colloids are at high elevations. By performing the same loop for different ferrofluid-water compositions, and hence varying the susceptibility χ, we can observe the transition from non-zero-homotopic loops in  to zero-homotopic loops. The results are shown in Fig. 5c. The topological transition takes place at χ≈0.1. For χ≲0.1, the particles descend below a critical elevation zc, and the transition to the non-transport phase occurs. Above zc the effects of the hexagonal pattern are the same for any z, and topologically protected modulation loops work for any hexagonal pattern, independently of the fine details. By decreasing the elevation below zc, we remove the holes of
to zero-homotopic loops. The results are shown in Fig. 5c. The topological transition takes place at χ≈0.1. For χ≲0.1, the particles descend below a critical elevation zc, and the transition to the non-transport phase occurs. Above zc the effects of the hexagonal pattern are the same for any z, and topologically protected modulation loops work for any hexagonal pattern, independently of the fine details. By decreasing the elevation below zc, we remove the holes of  inducing a topological transition. This plays the role of gap closure in the dispersion relation of wave-like systems28.
inducing a topological transition. This plays the role of gap closure in the dispersion relation of wave-like systems28.
Application
We use the topological protection to implement an experimental internal quality control of a chemical reaction. We consider the hybridization reaction between two complementary single-stranded oligo nucleotides of DNA, which we attach to the paramagnetic and diamagnetic colloids. If the hybridization is successful, the paramagnet (p) and diamagnet (d) irreversibly bind to form a quadrupole (q)

We want to emulate the conditional command: if the reaction is successful, then transport the product q along a given direction aq, otherwise transport the educts p and d along directions ap and ad, respectively.
We have already shown how to induce topologically protected transport of the educts (dipoles). The product of the reaction is a quadrupole that senses the quadrupolar potential Vq=−( V)2. The modulation loops
V)2. The modulation loops  and
and  for the transport of the educts can be chosen such that they do not affect the motion of the quadrupoles. We also find an appropriate modulation loop
for the transport of the educts can be chosen such that they do not affect the motion of the quadrupoles. We also find an appropriate modulation loop  that transports the quadrupoles in the required direction without affecting the dipoles.
that transports the quadrupoles in the required direction without affecting the dipoles.
The paramagnets and diamagnets reside on opposite parts ( and
and  ) of the stationary surface and, in the presence of the pattern, do not approach each other in
) of the stationary surface and, in the presence of the pattern, do not approach each other in  to allow the hybridization. To bring the colloids together, we perform a non-adiabatic hybridization loop
to allow the hybridization. To bring the colloids together, we perform a non-adiabatic hybridization loop  around the equator of
around the equator of  with a very high angular frequency. Hence, the colloids cannot follow the instantaneous potential and feel an almost flat averaged potential. Effectively, we erase the potential of the pattern such that the weak dipolar attractive interaction is enough to bring paramagnets and diamagnets together. The colloids meet in a bubble and rotate around each other such that hybridization is possible. After hybridization, the bond is strong enough to resist the magnetic stress exerted by the potential of the next modulation loops such that the bond is irreversible. The particle pair remains inseparable and behaves like a stable quadrupole. The entire modulation is a loop of the type
with a very high angular frequency. Hence, the colloids cannot follow the instantaneous potential and feel an almost flat averaged potential. Effectively, we erase the potential of the pattern such that the weak dipolar attractive interaction is enough to bring paramagnets and diamagnets together. The colloids meet in a bubble and rotate around each other such that hybridization is possible. After hybridization, the bond is strong enough to resist the magnetic stress exerted by the potential of the next modulation loops such that the bond is irreversible. The particle pair remains inseparable and behaves like a stable quadrupole. The entire modulation is a loop of the type  , see Fig. 6a. We show the transport of the colloids after a successful and a non-successful hybridization in Fig. 6b,c, respectively. Videos are provided in Supplementary Movies 8, 9. This quality control is internal as we do not change the modulation loop after we have measured externally whether the reaction was successful. The quality control works without active intervention.
, see Fig. 6a. We show the transport of the colloids after a successful and a non-successful hybridization in Fig. 6b,c, respectively. Videos are provided in Supplementary Movies 8, 9. This quality control is internal as we do not change the modulation loop after we have measured externally whether the reaction was successful. The quality control works without active intervention.

(a) Schematic of the hybridization reaction and the emulation of a conditional command that transport each type of particle in different directions. Polarization images of a successful and non-successful reaction are shown in b,c, respectively. The trajectories of the diamagnets, paramagnets and quadrupoles are highlighted in pink, cyan and yellow, respectively. The green (red) circle indicates the area where the dipoles meet and the hybridization takes place (fails). Scale bars (b,c), 10 μm.
Discussion
When the modulation loop is in the north of  , the magnetic potential presents two minima, one in n+1 and one in n+2 that are projected onto different points on
, the magnetic potential presents two minima, one in n+1 and one in n+2 that are projected onto different points on  . No colloidal transport between the minima exists as the potential barrier is too high. Phase space is hence divided into different nonergodic regions, and thermal equilibration only occurs over the cage around each minimum. The cage can only be left in a ratchet-like motion when the modulation loop touches the fence. Hence, our ratchet is associated with an ergodic-nonergodic transition, and might serve as a model for the cage effect in supercooled liquids29 and glasses30.
. No colloidal transport between the minima exists as the potential barrier is too high. Phase space is hence divided into different nonergodic regions, and thermal equilibration only occurs over the cage around each minimum. The cage can only be left in a ratchet-like motion when the modulation loop touches the fence. Hence, our ratchet is associated with an ergodic-nonergodic transition, and might serve as a model for the cage effect in supercooled liquids29 and glasses30.
Dissipation has been used in open quantum systems12 to isolate topologically protected edge modes from a bath. The final state of the edge modes is nevertheless dissipation free. In contrast, our system must be driven to maintain the transport. Moreover, the ratchet effect (crucial to change the direction of motion) is intrinsically dissipative and cannot, in general, be described with an effective Hamiltonian.
In one-dimensional systems, one can prove that thermal ratchets31, in which the potential evolves in time adiabatically, are always time reversal ratchets. We have shown here an example in two dimensions of a non-time reversal ratchet in an adiabatically evolving potential.
The construction of the stationary surface  and the mappings to action and control space is completely general and can be used for any potential. Other potentials might or might not support topologically protected transport, depending on the topological properties of
and the mappings to action and control space is completely general and can be used for any potential. Other potentials might or might not support topologically protected transport, depending on the topological properties of  . Our results are directly transferable to any system with hexagonal symmetry, and a potential proportional to the square of a field, which satisfies the Laplace equation.
. Our results are directly transferable to any system with hexagonal symmetry, and a potential proportional to the square of a field, which satisfies the Laplace equation.
High-quality magnetic bubbles lattices, like the one we have used here, have been studied extensively32 and hence the technology for its fabrication is well known. In addition, we note that any patterned substrates, such as lithographic magnetic patterns33, will induce similar transport.
Methods
Experimental preparation and measurements
The FGF films were grown by Tom Johansen (Oslo) via liquid phase epitaxy. We use the water-based ferrofluid EMG 707 from FerroTec GmbH, Germany. We dilute the ferrofluid with water. The final magnetic susceptibility is χ≈0.6. The time-dependent magnetic field is generated by three coils, following the ideas presented in ref. 34. Each coil controls the magnetic field along one of the three Cartesian axis. The current through the coils is provided by three phase-locked channels of programmable waveform generators (TTi TG1244) via three bipolar (KEPCO 20–50GL) amplifiers. The system is monitored via polarization microscopy. The pattern is visualized via the polar Faraday effect, and the colloids via ordinary transmission microscopy. Modulation loops in control space are programmed on a computer and transferred to the waveform generators.
For the hybridization reaction, we functionalize the colloids with streptavidin. The diamagnets and paramagnets are immersed separately into two solutions of biotinylated and complementary oligonucleotides. The complementary sequences are 5′-/5Bio/TCACTCAGTACGATATGCGGCACAG-3′ and 5′-/5Bio/CTGTGCCGCATATCGTACTGAGTGA-3′.
Topology of the stationary manifold
We first find the projection of the bifurcation points and the fences onto action space. Next, we map action space into control space, so that we obtain the projection of the fences and bifurcation points in  . With these projections we compute the vertices v=96, edges e=124 and areas a=16 of
. With these projections we compute the vertices v=96, edges e=124 and areas a=16 of  . The Euler characteristic of
. The Euler characteristic of  is χ(
is χ( )=v−e+a=−12, and it has genus g(
)=v−e+a=−12, and it has genus g( )=1−χ(
)=1−χ( )/2=7. The mapping of
)/2=7. The mapping of  into
into  also allows the determination of how the bijective areas are glued in
also allows the determination of how the bijective areas are glued in  . Further details are given below.
. Further details are given below.
Projection of the fence
We use coordinates

in action space  , where a1 and a2 are the basic lattice vectors of the hexagonal lattice. In control space, we use coordinates
, where a1 and a2 are the basic lattice vectors of the hexagonal lattice. In control space, we use coordinates

where the azimuthal angle ϕ is measured with respect to the direction of a1 and the polar angle θ with respect to the z-direction. Consider the unit vectors  (x1, x2)=∂iHg/|∂iHg|, i=1, 2, where Hg is the magnetic field of the garnet film. As we have seen, the leading term of the magnetic potential is V∝Hext·Hg and the stationary points are those for which
(x1, x2)=∂iHg/|∂iHg|, i=1, 2, where Hg is the magnetic field of the garnet film. As we have seen, the leading term of the magnetic potential is V∝Hext·Hg and the stationary points are those for which  . Then, a point (Hext,
. Then, a point (Hext,  ) in
) in  is stationary, and hence lies on
is stationary, and hence lies on  , if the direction of Hext is perpendicular to both
, if the direction of Hext is perpendicular to both  and
and  . Therefore, a point in
. Therefore, a point in  with coordinates (x1, x2) in the basis (a1, a2) has two stationary preimages in
with coordinates (x1, x2) in the basis (a1, a2) has two stationary preimages in  that correspond to external magnetic fields
that correspond to external magnetic fields  . The superscript (s) in
. The superscript (s) in  indicates that this field makes the point (x1, x2) in
indicates that this field makes the point (x1, x2) in  stationary. Consider now the Hessian matrix,
stationary. Consider now the Hessian matrix,

When crossing the fence on  from
from  to
to  , a saddle point changes to a minimum. Hence, the determinant of the above Hessian matrix must vanish at the fence,
, a saddle point changes to a minimum. Hence, the determinant of the above Hessian matrix must vanish at the fence,  . In this way, we find an implicit equation for the projection of the fences in action space. Both fences,
. In this way, we find an implicit equation for the projection of the fences in action space. Both fences,  and
and  , are projected into the same region in
, are projected into the same region in  with coordinates (x1,f, x2,f) given implicitly by:
with coordinates (x1,f, x2,f) given implicitly by:

where

The projection of the fence  in control space, that is, the northern border on
in control space, that is, the northern border on  is given by:
is given by:

where

The coordinates of the southern border on  are then obtained via the transformation θ→π−θ and ϕ→ϕ−π. Supplementary Fig. 1 shows the projection of the fences in the ϕ−θ plane of control space.
are then obtained via the transformation θ→π−θ and ϕ→ϕ−π. Supplementary Fig. 1 shows the projection of the fences in the ϕ−θ plane of control space.
Projection of bifurcation points
Four bijective areas meet at a bifurcation point in  . Four segments (two fence segments and two pseudo-fence segments) form the branches that bifurcate in a bifurcation point in
. Four segments (two fence segments and two pseudo-fence segments) form the branches that bifurcate in a bifurcation point in  . If we follow the fence
. If we follow the fence  and cross a bifurcation point, then either the minimum or the saddle point that meet at the fence (depending on the type of bifurcation point) changes the bijective area to which it belongs. If we cross the projection of a triple plus bifurcation point in
and cross a bifurcation point, then either the minimum or the saddle point that meet at the fence (depending on the type of bifurcation point) changes the bijective area to which it belongs. If we cross the projection of a triple plus bifurcation point in  from the tropics to the north, then in
from the tropics to the north, then in  a minimum undergoes a pitchfork bifurcation into two minima and one saddle point. An equivalent bifurcation happens if we cross the projection of a triple zero bifurcation point, in which case the roles of saddle points and minima are reversed.
a minimum undergoes a pitchfork bifurcation into two minima and one saddle point. An equivalent bifurcation happens if we cross the projection of a triple zero bifurcation point, in which case the roles of saddle points and minima are reversed.
The mathematical condition for a bifurcation point is as follows. Let v0 be the eigenvector of the Hessian matrix, cf. equation (5), at the fence with eigenvalue 0. Then, a bifurcation point is a fence point that satisfies

Solving the above equation, we find the projection in  of a triple plus bifurcation point lying on the fence
of a triple plus bifurcation point lying on the fence  at (θ, ϕ)=(π/3, π). The coordinates in
at (θ, ϕ)=(π/3, π). The coordinates in  of the projection of a triple zero bifurcation point in
of the projection of a triple zero bifurcation point in  are (θ, ϕ)=(0.381π, 7π/6). The other projections of bifurcation points belonging to the fence
are (θ, ϕ)=(0.381π, 7π/6). The other projections of bifurcation points belonging to the fence  are obtained, for symmetry reasons, via rotations around the z axis by multiples of π/3. Using the transformation θ→π−θ and ϕ→ϕ−π, one finds the projection of the bifurcation points in
are obtained, for symmetry reasons, via rotations around the z axis by multiples of π/3. Using the transformation θ→π−θ and ϕ→ϕ−π, one finds the projection of the bifurcation points in  .
.
Bijective areas and the genus of 

We can thus map each point in  onto two opposing points in
onto two opposing points in  . The mapping of a point in
. The mapping of a point in  onto the two points in
onto the two points in  will fall either in the north and the south (one point in each region), or both points fall into the tropics of
will fall either in the north and the south (one point in each region), or both points fall into the tropics of  . This gives the projections
. This gives the projections  of the bijective areas of
of the bijective areas of  into action space. Hence, we can see how the bijective areas are glued together in
into action space. Hence, we can see how the bijective areas are glued together in  and
and  . A bijective area is a connected preimage of either the north, the south or the tropics of control space. That is, there is a one-to-one correspondence between the bijective area in
. A bijective area is a connected preimage of either the north, the south or the tropics of control space. That is, there is a one-to-one correspondence between the bijective area in  and its corresponding region in
and its corresponding region in  . Any loop in
. Any loop in  lying entirely in a bijective area is projected onto a zero homotopic loop in
lying entirely in a bijective area is projected onto a zero homotopic loop in  . Hence, in order to achieve intercellular transport, a loop must cross different bijective areas. The neighbouring bijective areas in
. Hence, in order to achieve intercellular transport, a loop must cross different bijective areas. The neighbouring bijective areas in  are shown in Supplementary Fig. 3. We can use it to construct the sequence of bijective areas for a given
are shown in Supplementary Fig. 3. We can use it to construct the sequence of bijective areas for a given  . For example, consider the loop
. For example, consider the loop  =(1N, 4N). We start in the tropics, where there is only one minimum which is located in t+. Segment 1N connects t+ to n+2, and segment 4N connects n+2 to t+, which closes the loop.
=(1N, 4N). We start in the tropics, where there is only one minimum which is located in t+. Segment 1N connects t+ to n+2, and segment 4N connects n+2 to t+, which closes the loop.
To compute the Euler characteristic of  and hence its genus, we need to count the vertices, edges and bijective areas, as detailed next. Topologically, the north of
and hence its genus, we need to count the vertices, edges and bijective areas, as detailed next. Topologically, the north of  is a simply connected area (that is, all loops are zero homotopic) with 12 edges (segments of the borders) and 12 vertices (projection of bifurcation points). Each point in the north of
is a simply connected area (that is, all loops are zero homotopic) with 12 edges (segments of the borders) and 12 vertices (projection of bifurcation points). Each point in the north of  has 6 preimages on
has 6 preimages on  . Hence, the north of
. Hence, the north of  contributes with 6 bijective areas, 12 × 6=72 edges and 12 × 6=72 vertices to
contributes with 6 bijective areas, 12 × 6=72 edges and 12 × 6=72 vertices to  . A similar contribution comes from the south of
. A similar contribution comes from the south of  . The tropics of
. The tropics of  is a non-simply connected area, for example, the equator is zero homotopic in
is a non-simply connected area, for example, the equator is zero homotopic in  but not in the tropics of
but not in the tropics of  . To easily compute the Euler characteristic, we need simply connected areas. We make the tropics of
. To easily compute the Euler characteristic, we need simply connected areas. We make the tropics of  simply connected by cutting it along a longitude that connects the projection of two bifurcation points, one in each border of
simply connected by cutting it along a longitude that connects the projection of two bifurcation points, one in each border of  . The total number of edges of the tropics is thus 12+12+2=26 and the total number of vertices is 12+12+2=26. There are 4 preimages of the tropics on
. The total number of edges of the tropics is thus 12+12+2=26 and the total number of vertices is 12+12+2=26. There are 4 preimages of the tropics on  . Hence, the total contribution of the tropics of
. Hence, the total contribution of the tropics of  to
to  is 4 bijective areas, 4 × 26=104 edges and 4 × 26=104 vertices.
is 4 bijective areas, 4 × 26=104 edges and 4 × 26=104 vertices.
Next, we glue the bijective areas to form  . Two unglued edges are glued to form a single edge such that the number of glued edges of
. Two unglued edges are glued to form a single edge such that the number of glued edges of  is (72+72+104)/2=124. Regarding the vertices, we have 72+72+104=248 before gluing them in
is (72+72+104)/2=124. Regarding the vertices, we have 72+72+104=248 before gluing them in  . We have to subtract 8 vertices that were artificially produced by cutting the tropics of
. We have to subtract 8 vertices that were artificially produced by cutting the tropics of  in order to have a simply connected area. We have then 248−8=240 unglued vertices. There are two types of vertices on
in order to have a simply connected area. We have then 248−8=240 unglued vertices. There are two types of vertices on  : bifurcation points where 4 bijective areas meet, and pseudo-bifurcation points where 2 bijective areas meet. There are 24 bifurcation points on
: bifurcation points where 4 bijective areas meet, and pseudo-bifurcation points where 2 bijective areas meet. There are 24 bifurcation points on  . Hence, we need 4 × 24=96 unglued vertices to glue together the bijective areas at the bifurcation points. The remaining 240−96=144 unglued vertices are glued in pairs to form 144/2=72 pseudo bifurcation points. The total number of vertices on
. Hence, we need 4 × 24=96 unglued vertices to glue together the bijective areas at the bifurcation points. The remaining 240−96=144 unglued vertices are glued in pairs to form 144/2=72 pseudo bifurcation points. The total number of vertices on  is the sum of the number of bifurcation and pseudo bifurcation points: 24+72=96.
is the sum of the number of bifurcation and pseudo bifurcation points: 24+72=96.
Finally, the Euler characteristic of  is χ(
is χ( )=96−124+16=−12 and the genus of
)=96−124+16=−12 and the genus of  is g(
is g( )=1−χ(
)=1−χ( )/2=7. Similar arguments can be used to calculate the genus of the submanifolds that form
)/2=7. Similar arguments can be used to calculate the genus of the submanifolds that form  , that is,
, that is,  ,
,  and
and  . We show in Supplementary Fig. 2 a plaster model of
. We show in Supplementary Fig. 2 a plaster model of  .
.
Computer simulations
We simulate the motion of point dipoles moving in the potential given by equation (1) using Brownian dynamic simulations. The equation of motion is

where t is the time,  is the position of the dipoles in
is the position of the dipoles in  , ξ is the friction coefficient and η is a Gaussian random force with a variance given by the fluctuation-dissipation theorem. The plus (minus) sign in front of the potential holds for the diamagnetic (paramagnetic) colloids. The equation of motion is integrated in time with a standard Euler algorithm. We use a time step T/dt≈2·105 with T the period of a modulation loop
, ξ is the friction coefficient and η is a Gaussian random force with a variance given by the fluctuation-dissipation theorem. The plus (minus) sign in front of the potential holds for the diamagnetic (paramagnetic) colloids. The equation of motion is integrated in time with a standard Euler algorithm. We use a time step T/dt≈2·105 with T the period of a modulation loop  . Simulations fully reproduce the experimental phase diagram.
. Simulations fully reproduce the experimental phase diagram.
Data availability
The data that supports the findings of this study are available from the corresponding author upon request.
Additional information
How to cite this article: Loehr, J. et al. Topological protection of multiparticle dissipative transport. Nat. Commun. 7:11745 doi: 10.1038/ncomms11745 (2016).
References
Freedman, M. H., Kitaev, A., Larsen, M. J. & Wang, Z. Topological quantum computation. Bull. Amer. Math. Soc. 40, 31–38 (2003).
Kane, C. L. & Lubensky, T. C. Topological boundary modes in isostatic lattices. Nat. Phys 10, 39–45 (2014).
Hasan, M. Z. & Kane, C. L. Colloquium: topological insulators. Rev. Mod. Phys. 82, 3045–3067 (2010).
Chen, B. G., Upadhyaya, N. & Vitelli, V. Nonlinear conduction via solitons in a topological mechanical insulator. Proc. Natl Acad. Sci. USA 111, 13004–13009 (2014).
Paulose, J., Chen, B. G. & Vitelli, V. Topological modes bound to dislocations in mechanical metamaterials. Nat. Phys. 11, 153–156 (2015).
Nash, L. M. et al. Topological mechanics of gyroscopic metamaterials. Proc. Natl Acad. Sci. USA 112, 14495–14500 (2015).
Shen, S.-Q. Topological Insulators: Dirac Equation in Condensed Matters Springer Science & Business Media (2013).
Kitagawa, T., Berg, E., Rudner, M. & Demler, E. Topological characterization of periodically driven quantum systems. Phys. Rev. B. 82, 235114 (2010).
Rudner, M. S., Lindner, N. H., Berg, E. & Levin, M. Anomalous edge states and the bulk-edge correspondence for periodically driven two-dimensional systems. Phys. Rev. X 3, 031005 (2013).
Ghodbane, M. et al. Development and validation of a microfluidic immunoassay capable of multiplexing parallel samples in microliter volumes. Lab Chip 15, 3211–3221 (2015).
Gao, L., Tahir, M. A., Virgin, L. N. & Yellen, B. B. Multiplexing superparamagnetic beads driven by multi-frequency ratchets. Lab Chip. 11, 4214–4220 (2011).
Diehl, S., Rico, E., Baranov, M. A. & Zoller, P. Topology by dissipation in atomic quantum wires. Nat. Phys 7, 971–977 (2011).
Weimer, H., Müller, M., Lesanovsky, I., Zoller, P. & Büchler, H. P. A Rydberg quantum simulator. Nat. Phys 6, 382–388 (2010).
Olson, C. J., Reichhardt, C. & Nori, F. Nonequilibrium dynamic phase diagram for vortex lattices. Phys. Rev. Lett. 81, 3757–3760 (1998).
Korda, P. T., Taylor, M. B. & Grier, D. G. Kinetically locked-in colloidal transport in an array of optical tweezers. Phys. Rev. Lett. 89, 128301 (2002).
Reichhardt, C. & Nori, F. Phase locking, devil’s staircases, Farey trees, and Arnold tongues in driven vortex lattices with periodic pinning. Phys. Rev. Lett. 82, 414–417 (1999).
Kolton, A. B., Domínguez, D. & Grønbech-Jensen, N. Mode locking in ac-driven vortex lattices with random pinning. Phys. Rev. Lett. 86, 4112–4115 (2001).
Bohlein, T., Mikhael, J. & Bechinger, C. Observation of kinks and antikinks in colloidal monolayers driven across ordered surfaces. Nat. Mater. 11, 126–130 (2012).
Reimann, P. Brownian motors: noisy transport far from equilibrium. Phys. Rep. Rev. Phys. Lett. 361, 57–265 (2002).
Köhler, S., Lehmann, J. & Hänggi, P. Driven quantum transport on the nanoscale. Phys. Rep. Rev. Phys. Lett. 406, 379–443 (2005).
Simon, S. M., Peskin, C. S. & Oster, G. F. What drives the translocation of proteins. Proc. Natl Acad. Sci. USA 89, 3770–3774 (1992).
Jülicher, F., Ajdari, A. & Prost, J. Modeling molecular motors. Rev. Mod. Phys. 69, 1269–1281 (1997).
Doering, C. R., Horsthemke, W. & Riordan, J. Nonequilibrium fluctuation-induced transport. Phys. Rev. Lett. 72, 2984–2987 (1994).
Popescu, M. N., Arizmendi, C. M., Salas-Brito, A. L. & Family, F. Disorder induced diffusive transport in ratchets. Phys. Rev. Lett. 85, 3321–3324 (2000).
Savel’ev, S., Marchesoni, F. & Nori, F. Stochastic transport of interacting particles in periodically driven ratchets. Phys. Rev. E 70, 061107 (2004).
Tierno, P., Johansen, T. H. & Fischer, T. h. M. Localized and delocalized motion of colloidal particles on a magnetic bubble lattice. Phys. Rev. Lett. 99, 038303 (2007).
Tierno, P., Reddy, S. V., Roper, M. G., Johansen, T. H. & Fischer, T. h. M. Transport and separation of biomolecular cargo on paramagnetic colloidal particles in a magnetic ratchet. J. Phys. Chem. B. 112, 3833–3837 (2008).
Zhang, F., MacDonald, A. H. & Mele, E. J. Valley Chern numbers and boundary modes in gapped bilayer graphene. Proc. Natl Acad. Sci. USA 110, 10546–10551 (2013).
Götze, W. & Sjögren, L. Relaxation processes in supercooled liquids. Rep. Prog. Phys 55, 241–376 (1992).
Doliwa, B. & Heuer, A. Cage effect, local anisotropies, and dynamic heterogeneities at the glass transition: A computer study of hard spheres. Phys. Rev. Lett. 80, 4915–4918 (1998).
Sinitsyn, N. A. The stochastic pump effect and geometric phases in dissipative and stochastic systems. J. Phys. A 42, 193001 (2009).
Bobeck, A. H., Bonyhard, P. I. & Geusic, J. E Magnetic bubbles.-emerging new memory technology. Proc. IEEE 63, 1176–1195 (1975).
Terris, B. D. & Thomson, T. Nanofabricated and self-assembled magnetic structures as data storage media. J. Phys. D Appl. Phys 38, R199–R222 (2005).
Martin, J. E. & Solis, K. J. Fully alternating, triaxial electric or magnetic fields offer new routes to fluid vorticity. Soft Matter 11, 241–254 (2015).
Acknowledgements
We thank Ingrid Bauer, Matthias Schmidt, Pietro Tierno and Antonio Ortiz-Ambriz for illuminating discussions and critical reading of the manuscript. We thank Thomas Hauenstein and Konrad Stern for playing the Franconian Landler with Johannes Loehr. Publication costs have been partially funded by the Profilfeld Polymer- und Kolloidforschung of the University of Bayreuth.
Author information
Authors and Affiliations
Contributions
J.L. performed the experiments, designed the modulation loops and played the accordion in movie ‘Franconian Landler.avi’. M.L. developed the topological relation between  ,
,  and
and  . D.de las H. had the idea to use Brownian dynamics to simulate the motion. Simulations were performed by A.E. and led to the discovery of the topological protection of the ratchet. T.M.F. had the idea of the experiments. J.L., D.de las H. and T.M.F. wrote the manuscript.
. D.de las H. had the idea to use Brownian dynamics to simulate the motion. Simulations were performed by A.E. and led to the discovery of the topological protection of the ratchet. T.M.F. had the idea of the experiments. J.L., D.de las H. and T.M.F. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Figures 1-4 and Supplementary Note 1 (PDF 6251 kb)
Supplementary Movie 1
Paramagnetic and diamagnetic colloids both moving along the x-axis. (AVI 502 kb)
Supplementary Movie 2
Diamagnetic colloids moving along the x-axis and paramagnetic colloids moving at an angle of π/3 with respect to the motion of the diamagnets. (AVI 254 kb)
Supplementary Movie 3
Diamagnetic colloids moving along the x-axis and paramagnetic colloids moving at an angle of 2π/3 with respect to the motion of the diamagnets. (AVI 360 kb)
Supplementary Movie 4
Diamagnetic colloids moving along the x-axis and paramagnetic colloids moving at an angle of π with respect to the motion of the diamagnets. (AVI 258 kb)
Supplementary Movie 5
Diamagnetic colloids moving along the x-axis and paramagnetic colloids moving at an angle of 4π/3 with respect to the motion of the diamagnets. (AVI 167 kb)
Supplementary Movie 6
Diamagnetic colloids moving along the x-axis and paramagnetic colloids moving at an angle of 5π/3 with respect to the motion of the diamagnets. (AVI 145 kb)
Supplementary Movie 7
Diamagnetic and paramagnetic colloids performing an adiabatic Franconian Landler. (AVI 971 kb)
Supplementary Movie 8
Quality control of a hybridization reaction. A successful hybridization of a paramagnet and a diamagnet results in the formation of a quadrupole. Diamagnets, paramagnets, and quadrupoles move along different directions. (AVI 1662 kb)
Supplementary Movie 9
Quality control of a hybridization reaction. An unsuccessful hybridization of a diamagnet and a paramagnet. The motion direction of paramagnets and diamagnets is unaffected. (AVI 1825 kb)
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Loehr, J., Loenne, M., Ernst, A. et al. Topological protection of multiparticle dissipative transport. Nat Commun 7, 11745 (2016). https://doi.org/10.1038/ncomms11745
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/ncomms11745
This article is cited by
-
Disorder scattering in classical flat channel transport of particles between twisted magnetic square patterns
Communications Physics (2024)
-
Simultaneous and independent topological control of identical microparticles in non-periodic energy landscapes
Nature Communications (2023)
-
Enhanced colloidal transport in twisted magnetic patterns
Communications Physics (2022)
-
Emergent colloidal currents across ordered and disordered landscapes
Communications Physics (2021)
-
Noether’s theorem in statistical mechanics
Communications Physics (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
