
Abstract
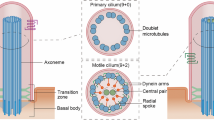
Mammalian Hedgehog (Hh) signal transduction requires a primary cilium, a microtubule-based organelle, and the Gli–Sufu complexes that mediate Hh signalling, which are enriched at cilia tips. Kif7, a kinesin-4 family protein, is a conserved regulator of the Hh signalling pathway and a human ciliopathy protein. Here we show that Kif7 localizes to the cilium tip, the site of microtubule plus ends, where it limits cilium length and controls cilium structure. Purified recombinant Kif7 binds the plus ends of growing microtubules in vitro, where it reduces the rate of microtubule growth and increases the frequency of microtubule catastrophe. Kif7 is not required for normal intraflagellar transport or for trafficking of Hh pathway proteins into cilia. Instead, a central function of Kif7 in the mammalian Hh pathway is to control cilium architecture and to create a single cilium tip compartment, where Gli–Sufu activity can be correctly regulated.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others


References
Bieling, P., Telley, I. A. & Surrey, T. A minimal midzone protein module controls formation and length of antiparallel microtubule overlaps. Cell 142, 420–432 (2010).
Hu, C-K., Coughlin, M., Field, C. M. & Mitchison, T. J. KIF4 regulates midzone length during cytokinesis. Curr. Biol. 21, 815–824 (2011).
Stumpff, J., Wagenbach, M., Franck, A., Asbury, C. L. & Wordeman, L. Kif18A and chromokinesins confine centromere movements via microtubule growth suppression and spatial control of kinetochore tension. Dev. Cell 22, 1017–1029 (2012).
Subramanian, R., Ti, S-C., Tan, L., Darst, S. A. & Kapoor, T. M. Marking and measuring single microtubules by PRC1 and kinesin-4. Cell 154, 377–390 (2013).
Van der Vaart, B. et al. CFEOM1-associated kinesin KIF21A is a cortical microtubule growth inhibitor. Dev. Cell 2013, 10.1016/j.devcel.2013.09.010
Liem, K. F., HE, M., Ocbina, P. J. R. & Anderson, K. V. Mouse Kif7/Costal2 is a cilia-associated protein that regulates Sonic hedgehog signaling. Proc. Natl Acad. Sci. USA 106, 13377–13382 (2009).
Tay, S. Y., Ingham, P. W. & Roy, S. A homologue of the Drosophila kinesin-like protein Costal2 regulates Hedgehog signal transduction in the vertebrate embryo. Development 132, 625–634 (2005).
Cheung, H. O-L. et al. The kinesin protein Kif7 is a critical regulator of Gli transcription factors in mammalian hedgehog signaling. Sci. Signal. 2, ra29 (2009).
Endoh-Yamagami, S. et al. The mammalian Cos2 homolog Kif7 plays an essential role in modulating Hh signal transduction during development. Curr. Biol. 19, 1320–1326 (2009).
Jiang, J. & Hui, C-C. Hedgehog signaling in development and cancer. Dev. Cell 15, 801–812 (2008).
Ingham, P. W., Nakano, Y. & Seger, C. Mechanisms and functions of Hedgehog signalling across the metazoa. Nat. Rev. Genet. 12, 393–406 (2011).
Ingham, P. W. & McMahon, A. P. Hedgehog signalling: Kif7 is not that fishy after all. Curr. Biol. 19, R729–R731 (2009).
Putoux, A. et al. Kif7 mutations cause fetal hydrolethalus and acrocallosal syndromes. Nat. Genet. 43, 601–606 (2011).
Dafinger, C. et al. Mutations in Kif7 link Joubert syndrome with Sonic Hedgehog signaling and microtubule dynamics. J. Clin. Invest. 121, 2662–2667 (2011).
Huangfu, D. et al. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature 426, 83–87 (2003).
Goetz, S. C. & Anderson, K. V. The primary cilium: a signalling centre during vertebrate development. Nat. Rev. Genet. 11, 331–344 (2010).
Matthies, H. J., Baskin, R. J. & Hawley, R. S. Orphan kinesin NOD lacks motile properties but does possess a microtubule-stimulated ATPase activity. Mol. Biol. Cell 12, 4000–4012 (2001).
Sisson, J. C., Ho, K. S., Suyama, K. & Scott, M. P. Costal2, a novel kinesin-related protein in the Hedgehog signaling pathway. Cell 90, 235–245 (1997).
Robbins, D. J. et al. Hedgehog elicits signal transduction by means of a large complex containing the kinesin-related protein costal2. Cell 90, 225–234 (1997).
Klejnot, M. & Kozielski, F. Structural insights into human Kif7, a kinesin involved in Hedgehog signalling. Acta Crystallogr. D 68, 154–159 (2012).
Haycraft, C. J. et al. Gli2 and Gli3 localize to cilia and require the intraflagellar transport protein polaris for processing and function. PLoS Genet. 1, e53 (2005).
Qin, J., Lin, Y., Norman, R. X., Ko, H. W. & Eggenschwiler, J. T. Intraflagellar transport protein 122 antagonizes Sonic Hedgehog signaling and controls ciliary localization of pathway components. Proc. Natl Acad. Sci. USA 108, 1456–1461 (2011).
Wen, X. et al. Kinetics of hedgehog-dependent full-length Gli3 accumulation in primary cilia and subsequent degradation. Mol. Cell. Biol. 30, 1910–1922 (2010).
Tukachinsky, H., Lopez, L. V. & Salic, A. A mechanism for vertebrate Hedgehog signaling: recruitment to cilia and dissociation of SuFu–Gli protein complexes. J. Cell Biol. 191, 415–428 (2010).
Caspary, T., Larkins, C. E. & Anderson, K. V. The graded response to Sonic Hedgehog depends on cilia architecture. Dev. Cell 12, 767–778 (2007).
Janke, C. & Bulinski, J. C. Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat. Rev. Mol. Cell Biol. 12, 773–786 (2011).
Pan, J. & Snell, W. J. Chlamydomonas shortens its flagella by activating axonemal disassembly, stimulating IFT particle trafficking, and blocking anterograde cargo loading. Dev. Cell 9, 431–438 (2005).
Marshall, W. F., Qin, H., Rodrigo Brenni, M. & Rosenbaum, J. L. Flagellar length control system: testing a simple model based on intraflagellar transport and turnover. Mol. Biol. Cell 16, 270–278 (2005).
Cole, D. G. Kinesin-II, coming and going. J. Cell Biol. 147, 463–466 (1999).
Insinna, C., Pathak, N., Perkins, B., Drummond, I. & Besharse, J. C. The homodimeric kinesin, Kif17, is essential for vertebrate photoreceptor sensory outer segment development. Dev. Biol. 316, 160–170 (2008).
Morsci, N. S. & Barr, M. M. Kinesin-3 KLP-6 regulates intraflagellar transport in male-specific cilia of Caenorhabditis elegans. Curr. Biol. 21, 1239–1244 (2011).
Ou, G., Blacque, O. E., Snow, J. J., Leroux, M. R. & Scholey, J. M. Functional coordination of intraflagellar transport motors. Nature 436, 583–587 (2005).
Miki, H., Okada, Y. & Hirokawa, N. Analysis of the kinesin superfamily: insights into structure and function. Trends Cell Biol. 15, 467–476 (2005).
Bringmann, H. et al. A kinesin-like motor inhibits microtubule dynamic instability. Science 303, 1519–1522 (2004).
Brady, S. T. A novel brain ATPase with properties expected for the fast axonal transport motor. Nature 317, 73–75 (1985).
Scholey, J. M., Porter, M. E., Grissom, P. M. & McIntosh, J. R. Identification of kinesin in sea urchin eggs, and evidence for its localization in the mitotic spindle. Nature 318, 483–486 (1985).
Vale, R. D., Reese, T. S. & Sheetz, M. P. Identification of a novel force-generating protein, kinesin, involved in microtubule-based motility. Cell 42, 39–50 (1985).
Bieling, P. et al. Reconstitution of a microtubule plus-end tracking system in vitro. Nature 450, 1100–1105 (2007).
Howard, J. & Hyman, A. A. Dynamics and mechanics of the microtubule plus end. Nature 422, 753–758 (2003).
Howard, J. & Hyman, A. A. Growth, fluctuation and switching at microtubule plus ends. Nat. Rev. Mol. Cell Biol. 10, 569–574 (2009).
Bhogaraju, S. et al. Molecular basis of tubulin transport within the cilium by IFT74 and IFT81. Science 341, 1009–1012 (2013).
Humke, E. W., Dorn, K. V., Milenkovic, L., Scott, M. P. & Rohatgi, R. The output of Hedgehog signaling is controlled by the dynamic association between suppressor of fused and the Gli proteins. Genes Dev. 24, 670–682 (2010).
Piao, T. et al. A microtubule depolymerizing kinesin functions during both flagellar disassembly and flagellar assembly in Chlamydomonas. Proc. Natl Acad. Sci. USA 106, 4713–4718 (2009).
Niwa, S. et al. KIF19A is a microtubule-depolymerizing kinesin for ciliary length control. Dev. Cell (2012)doi:10.1016/j.devcel.2012.10.016
Helenius, J., Brouhard, G., Kalaidzidis, Y., Diez, S. & Howard, J. The depolymerizing kinesin MCAK uses lattice diffusion to rapidly target microtubule ends. Nature 441, 115–119 (2006).
Akhmanova, A. & Steinmetz, M. O. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat. Rev. Mol. Cell Biol. 9, 309–322 (2008).
Dentler, W. L. & Rosenbaum, J. L. Flagellar elongation and shortening in Chlamydomonas. III. structures attached to the tips of flagellar microtubules and their relationship to the directionality of flagellar microtubule assembly. J. Cell Biol. 74, 747–759 (1977).
Pedersen, L. B. et al. Chlamydomonas IFT172 is encoded by FLA11, interacts with CrEB1, and regulates IFT at the flagellar tip. Curr. Biol. 15, 262–266 (2005).
Scholey, J. M. & Anderson, K. V. Intraflagellar transport and cilium-based signaling. Cell 125, 439–442 (2006).
Zhang, X. M., Ramalho-Santos, M. & McMahon, A. P. Smoothened mutants reveal redundant roles for Shh and Ihh signaling including regulation of L/R asymmetry by the mouse node. Cell 105, 781–792 (2001).
Matise, M. P., Epstein, D. J., Park, H. L., Platt, K. A. & Joyner, A. L. Gli2 is required for induction of floor plate and adjacent cells, but not most ventral neurons in the mouse central nervous system. Development 125, 2759–2770 (1998).
Ocbina, P. J. R. & Anderson, K. V. Intraflagellar transport, cilia, and mammalian Hedgehog signaling: analysis in mouse embryonic fibroblasts. Dev. Dyn. 237, 2030–2038 (2008).
Liem, K. F. et al. The IFT-A complex regulates Shh signaling through cilia structure and membrane protein trafficking. J. Cell Biol. 197, 789–800 (2012).
Hyman, A. A. Preparation of marked microtubules for the assay of the polarity of microtubule-based motors by fluorescence. J. Cell Sci. Suppl. 14, 125–127 (1991).
Acknowledgements
We thank L. Gunther-Cummins, P. Satir (Albert Einstein College of Medicine, Bronx, NY), K. Uryu (Rockefeller University) for assistance with TEM, and Alan Hall for use of a microscope for TIRF imaging. We thank H. Bazzi, A. Parrish and T. Bestor for helpful comments on the manuscript. We thank B. Tsou and W-J. Wang for technical support and helpful discussion. We thank C.C. Hui (Sick Children’s Hospital) for providing Kif7−/− mice. We thank J. Eggenschwiler (University of Georgia) and A. Salic (Harvard University) for antibodies. We thank N. Lampen for help with scanning electron microscopy and the Memorial Sloan-Kettering Cancer Center core facilities and the Rockefeller University Bio-Imaging Resource Center for help with imaging. We thank R. Hendrickson and H. Erdjument-Bromage for help with mass spectrometry. This work was supported by National Institutes of Health grants NS044385 to K.V. Anderson and GM65933 to T.M. Kapoor and the MSKCC Cancer Center Support Grant P30 CA008748.
Author information
Authors and Affiliations
Contributions
M.H. designed, carried out and analysed experiments and assembled figures. R.S. and M.H. carried out and analysed the microtubule dynamics assays. F.B. carried out IFT assays. T.O. helped with TIRF microscopy. K.F.L. did the TEM. T.M.K. designed, supervised and analysed the microtubule dynamics assays. R.S. and T.M.K. helped write and edit the manuscript. K.V.A. designed, supervised and analysed experiments. M.H. and K.V.A. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Kif7L130P encodes a stable protein that does not localize to cilium tips.
(a) Immunoblot with Kif7 antibody in cell lysates of wild-type, Kif7L130P and Kif7−/− MEFs. (b) Kif7 (green) is absent from Kif7L130P cilia regardless of pathway activation. Acetylated α-tubulin (red) stains cilia and γ-tubulin (blue) stains basal body. Arrows indicate Kif7 at the transition zone. Scale bar, 1 μm. (c) Kif7 localizes to cilium tip in Smo−/− and (d) Gli2−/−Gli3−/− double mutant MEFs. Acetylated α-tubulin (red) stains cilia and γ-tubulin (blue) in (c) and magenta in (d) stains basal body. Scale bar, 5 μm. (e) GM130 (green), a Golgi membrane protein, appears normal in Kif7L130P MEFs. DAPI in blue. Scale bar, 17 μm. (f) In contrast to Kif7 knockdown cells14, normal numbers of centrosomes and cilia are present in wild-type and Kif7L130P MEFs. Acetylated α-tubulin (red) marks axonemal microtubules and γ-tubulin (magenta) marks basal bodies. DAPI is in blue. n = 165 cilia were analysed and were pooled from 4 independent experiments. Error bars represent the s.d., P > 0.0001 by student t-test. Scale bar, 22 μm.
Supplementary Figure 2 Kif7L130P mutant cells have long cilia and abnormal axonemal structure.
(a) The diameter of proximal and distal region was measured from neural tube cilia from e10.5 wild-type and Kif7L130P embryos. The width of proximal cilia was 0.21 ± 0.02 μm in wild-type and 0.20 ± 0.02 μm in Kif7L130P. The width of distal cilia was 0.12 ± 0.02 μm in wild-type and 0.08 ± 0.01 μm in Kif7L130P (n = 50 cilia were measured for each genotype; P < 0.001 by one-way ANOVA analysis). Error bars represent the s.d. (b) Low power s.e.m. of the node of an e8.0 wild-type embryo (black square). (c) Node cilia from e8.0 wild-type and Kif7L130P mutant embryos. Scale bar, 2 μm. (d) TEM images of transverse sections of Kif7L130P neural tubule cilia (4 examples). Arrows indicate singlet microtubules. Scale bars, 50 nm.
Supplementary Figure 3 Defects of tubulin modification in Kif7−/− mutant MEFs.
Tubulin acetylation (red) and glutamylation (green) are reduced at the distal segment of cilia in Kif7−/− MEFs compared to wild type. Scale bar, 1 μm.
Supplementary Figure 4 Kif7 associates with microtubule in vivo.
(a) Protein domain analysis of mouse Kif7 and KIF4. Motor domain is in green, neck in pink and coiled-coil in purple. Note the long linker between the neck and the first coiled-coil of Kif7, which may inhibit its ability to act as a processive motor. Arrows and numbers indicate the positions of respective domains in amino acid. (b) Co-immunoprecipitation of Flag- and GFP-tagged full-length Kif7 in 293T cells. (c) Expression of C-terminal GFP-tagged Kif7-full length and Kif7560 constructs in HEK293T cells visualized by confocal fluorescent microscope. DAPI in blue. Scale bar, 8 μm. (d) Deletion analysis of Kif7 for in vivo microtubule bundling activity and dimerization.
Supplementary Figure 5 Purified Kif7560-GFP directly binds to microtubules.
(a) Purified Kif7560-GFP for in vitro analysis. Asterisk indicates bacterial protein Arna (Bifunctional polymyxin resistance protein Arna). (b) 70 nM Kif7560-GFP binds to X-rhodamine-labelled microtubules in the presence of 1 mM MgATP or 2 mM AMP-PNP. Scale bar, 5 μm.
Supplementary Figure 6 Induced microtubule destabilization by purified Kif7560-GFP.
Examples of fragmented microtubules observed when X-rhodamine-labeled microtubules were incubated with 500 nM of Kif7560-GFP in the presence of 1 mM MgATP.
Supplementary Figure 7 Kif7L130P mutant cilia have defects in IFT dynamics.
(a–b) Representative kymographs generated from time lapse imaging of wild-type and (C–D) Kif7L130P primary cilium (Supplementary Videos 1 and 2). (a) and (c) are taken from Fig. 3. The base and tip of the cilium are indicated with arrowheads. Asterisks indicate stationary IFT88 puncta associated with the initiation of retrograde trafficking. Horizontal scale bar (distance), 1 μm. Vertical scale bar (time), 7.5 s. Enlarged portion of kymographs are highlighted (yellow rectangles). Asterisks show the position of stationary IFT88 puncta. Our kymograph analysis shows IFT88–GFP (4/200 retrograde trains) switches tracks in the wild-type cilia very rarely, whereas significantly more trains (79/214 retrograde trains) switch tracks in Kif7 mutants. (e) Numbers of stationary IFT88–GFP puncta per cilium in wild-type and Kif7L130P (n = 50 cilia were imaged for each condition and were pooled from 5 independent experiments; P < 0.0001 by student t-test). The error bars represent s.d.
Supplementary Figure 8 Uncropped western blots referring to Supplementary Figs 1a and 4b.
Supplementary Fig. 1a Kif7 protein level in wild-type, Kif7L130P and Kif7−/− MEFs.γ-tubulin (asterisk) was used as loading control. Supplementary Fig. 4b co-immunoprecipitation of Flag- and GFP-tagged full-length Kif7 in 293T cells. Membranes were blotted with anti-Flag and anti-GFP antibodies, respectively.
Supplementary information
Supplementary Information
Supplementary Information (PDF 1494 kb)
Live imaging of IFT88–GFP in wild-type cilia.
IFT visualized by time-lapse imaging of a wild-type primary cilium expressing IFT88–GFP. Images are taken every 250 ms for 30 s. Movie shows 10 frames s−1. (MOV 1813 kb)
Live imaging of IFT88–GFP in Kif7L130P mutant cilia.
IFT visualized by time-lapse imaging of a Kif7L130P primary cilium expressing IFT88–GFP. Images are taken every 250 ms for 30 s. Movie shows 10 frames s−1. (MOV 1607 kb)
Effect of Kif7560-GFP on dynamic microtubules.
Kif7560-GFP (70 nM; 1 mM MgATP) and dynamic X-rhodamine-microtubule visualized by time-lapse TIRF microscopy. Images are taken every 5 s for 15 min. Movie shows 15 frames s−1. (MOV 8959 kb)
Rights and permissions
About this article

Cite this article
He, M., Subramanian, R., Bangs, F. et al. The kinesin-4 protein Kif7 regulates mammalian Hedgehog signalling by organizing the cilium tip compartment. Nat Cell Biol 16, 663–672 (2014). https://doi.org/10.1038/ncb2988
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb2988
This article is cited by
-
Molecular diagnosis and novel genes and phenotypes in a pediatric thoracic insufficiency cohort
Scientific Reports (2023)
-
The role of Hedgehog and Notch signaling pathway in cancer
Molecular Biomedicine (2022)
-
A kinesin mimics DNA
Nature Cell Biology (2022)
-
Cytoskeletal regulation of a transcription factor by DNA mimicry via coiled-coil interactions
Nature Cell Biology (2022)
-
Exploring Sonic Hedgehog Cell Signaling in Neurogenesis: Its Potential Role in Depressive Behavior
Neurochemical Research (2021)
