
Abstract
Nutrient deprivation is a stimulus shared by both autophagy and the formation of primary cilia. The recently discovered role of primary cilia in nutrient sensing and signalling motivated us to explore the possible functional interactions between this signalling hub and autophagy. Here we show that part of the molecular machinery involved in ciliogenesis also participates in the early steps of the autophagic process. Signalling from the cilia, such as that from the Hedgehog pathway, induces autophagy by acting directly on essential autophagy-related proteins strategically located in the base of the cilium by ciliary trafficking proteins. Whereas abrogation of ciliogenesis partially inhibits autophagy, blockage of autophagy enhances primary cilia growth and cilia-associated signalling during normal nutritional conditions. We propose that basal autophagy regulates ciliary growth through the degradation of proteins required for intraflagellar transport. Compromised ability to activate the autophagic response may underlie some common ciliopathies.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

References
Mizushima, N., Levine, B., Cuervo, A. M. & Klionsky, D. J. Autophagy fights disease through cellular self-digestion. Nature 451, 1069–1075 (2008)
Hamasaki, M., Shibutani, S. T. & Yoshimori, T. Up-to-date membrane biogenesis in the autophagosome formation. Curr. Opin. Cell Biol. 25, 455–460 (2013)
Yang, Z. & Klionsky, D. J. Mammalian autophagy: core molecular machinery and signaling regulation. Curr. Opin. Cell Biol. 22, 124–131 (2010)
Satir, P., Pedersen, L. B. & Christensen, S. T. The primary cilium at a glance. J. Cell Sci. 123, 499–503 (2010)
Taschner, M., Bhogaraju, S. & Lorentzen, E. Architecture and function of IFT complex proteins in ciliogenesis. Differentiation 83, S12–S22 (2012)
Follit, J. A., Xu, F., Keady, B. T. & Pazour, G. J. Characterization of mouse IFT complex B. Cell Motil. Cytoskeleton 66, 457–468 (2009)
Liem, K. F. et al. The IFT-A complex regulates Shh signaling through cilia structure and membrane protein trafficking. J. Cell Biol. 197, 789–800 (2012)
Rohatgi, R., Milenkovic, L. & Scott, M. P. Patched1 regulates Hedgehog signaling at the primary cilium. Science 317, 372–376 (2007)
Kiprilov, E. N. et al. Human embryonic stem cells in culture possess primary cilia with hedgehog signaling machinery. J. Cell Biol. 180, 897–904 (2008)
Onodera, J. & Ohsumi, Y. Autophagy is required for maintenance of amino acid levels and protein synthesis under nitrogen starvation. J. Biol. Chem. 280, 31582–31586 (2005)
Tanida, I., Minematsu-Ikeguchi, N., Ueno, T. & Kominami, E. Lysosomal turnover, but not a cellular level, of endogenous LC3 is a marker for autophagy. Autophagy 1, 84–91 (2005)
Klionsky, D. J. et al. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy 8, 445–544 (2012)
Pazour, G. J., San Agustin, J. T., Follit, J. A., Rosenbaum, J. L. & Witman, G. B. Polycystin-2 localizes to kidney cilia and the ciliary level is elevated in orpk mice with polycystic kidney disease. Curr. Biol. 12, R378–R380 (2002)
Schneider, L. et al. PDGFRαα signaling is regulated through the primary cilium in fibroblasts. Curr. Biol. 15, 1861–1866 (2005)
Boehlke, C. et al. Primary cilia regulate mTORC1 activity and cell size through Lkb1. Nature Cell Biol. 12, 1115–1122 (2010)
Finetti, F. et al. Intraflagellar transport is required for polarized recycling of the TCR/CD3 complex to the immune synapse. Nature Cell Biol. 11, 1332–1339 (2009)
Ravikumar, B., Moreau, K., Jahreiss, L., Puri, C. & Rubinsztein, D. C. Plasma membrane contributes to the formation of pre-autophagosomal structures. Nature Cell Biol. 12, 747–757 (2010)
Veland, I. R. et al. Inversin/nephrocystin-2 is required for fibroblast polarity and directional cell migration. PLoS ONE 8, e60193 (2013)
Sridhar, S. et al. The lipid kinase PI4KIIIβ preserves lysosomal identity. EMBO J. 32, 324–339 (2012)
Richey, E. A. & Qin, H. Dissecting the sequential assembly and localization of intraflagellar transport particle complex B in Chlamydomonas. PLoS ONE 7, e43118 (2012)
Satir, P. The new biology of cilia: review and annotation of a symposium. Dev. Dyn. 241, 426–430 (2012)
Li, H. et al. Sonic hedgehog promotes autophagy of vascular smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 303, H1319–H1331 (2012)
Jimenez-Sanchez, M. et al. The Hedgehog signalling pathway regulates autophagy. Nature Comm. 3, 1200 (2012)
Wang, Y., Han, C., Lu, L., Magliato, S. & Wu, T. Hedgehog signaling pathway regulates autophagy in human hepatocellular carcinoma cells. Hepatology 58, 995–1010 (2013)
Petralia, R. S. et al. Sonic hedgehog promotes autophagy in hippocampal neurons. Biol. Open 2, 499–504 (2013)
Singh, R. et al. Autophagy regulates lipid metabolism. Nature 458, 1131–1135 (2009)
Auteri, J. S., Okada, A., Bochaki, V. & Dice, J. F. Regulation of intracellular protein degradation in IMR-90 human diploid fibroblasts. J. Cell. Physiol. 115, 167–174 (1983)
Huang, B. Q. et al. Isolation and characterization of cholangiocyte primary cilia. Am. J. Physiol. Gastrointest. Liver Physiol. 291, G500–G509 (2006)
Marzella, L., Ahlberg, J. & Glaumann, H. Isolation of autophagic vacuoles from rat liver: morphological and biochemical characterization. J. Cell Biol. 93, 144–154 (1982)
Wada, K. et al. Application of photoacoustic microscopy to analysis of biological components in tissue sections. Chem. Pharm. Bull. (Tokyo) 34, 1688–1693 (1986)
Acknowledgements
We thank N. Mizushima and G. Pazour for providing MEFs and I. R. Veland and S. T. Christensen for providing the plasmid for expression of GFP–inversin. We also thank the personnel at the Analytical Imaging Facility for their technical assistance with TEM, J. Kraut and N. Rodriguez for their early assistance in cilia staining optimization and S. Kaushik for critically reviewing this manuscript. This work was supported by grants from the National Institute of Health National Institute on Aging AG031782 and AG038072, National Institute of Diabetes and Digestive and Kidney Diseases DK098408 (to A.M.C.), Institut National de la Santé et de la Recherche Medicale (to P.C. and I.B.), Institut National du Cancer and L’Agence Nationale de la Recherche (to P.C.), and the generous support of Robert and Renee Belfer (to A.M.C.). O.P. was supported by a Basque Government Postdoctoral Fellowship.
Author information
Authors and Affiliations
Contributions
O.P. designed and performed most of the experiments, analysed the data and contributed to writing the paper; I.O. and I.B. performed and analysed the experiments related to Hh signalling; B.P. performed the electron microscopy studies and morphometric analysis; S.S. performed the cytosolic vesicle experiments; A.D.-C. assisted with cell culture; P.C. conceived the part of the study related to Hh signalling, provided interpretation to the data and contributed to the writing and revision of the paper; B.H.S. and P.S. set the bases for the rationale of the study, provided feedback in the interpretation of the data and revised the written manuscript; A.M.C. coordinated the study, designed experiments, analysed data and contributed to the writing and revision of the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
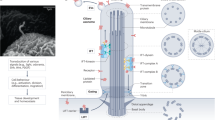
Extended Data Figure 1 Schematic model of the interplay between autophagy and ciliogenesis.
Starvation conditions (serum−) increase intraflagellar transport (IFT) and ciliogenesis and Hedgehog (Hh) signalling. We show in this work that both events increase autophagosome formation interdependently. ATG16L is trafficked in IFT20-enriched vesicles to the base of the cilia where most ATG localize. Arrival of ATG16L to this ATG assembly and/or its transport along the axoneme seems to initiate the elongation of ATG5 structures and presence of lipid-binding ATGs (such as LC3 and GABARAP) in the cilia and at the plasma membrane. The cilium also contains under these conditions molecules of SMO that are mobilized to the ciliary membrane after release of the inhibitory effect of Ptc. Activated SMO induces the downstream transcription factors GLI1/GLI2 which further favours recruitment and assembly of ATGs in the base of the cilium. Normal nutritional conditions (serum+) lead to decreased Hh and IFT and suppression of starvation-induced autophagy. Basal autophagy contributes to maintain ciliogenesis to a minimum through the degradation of IFT20, which in turns reduces trafficking of ATG16L into the ATG assembly at the base of the cilium, preventing induction of ciliary autophagy.
Extended Data Figure 2 Ciliogenesis and LC3 in different cell types.
a, Immunoblot for IFT20 and IFT88 and quantification in MEFs control (Ctrl) or knocked down (−) for IFT20 (**P = 0.007, n = 7), or kidney epithelial cells (KECs) wild-type (WT) or knockout for IFT88 (Ift88−/−; **P = 0.0003, n = 3). ADU, arbitrary densitometric units (n = 3). b, Rates of degradation of long-lived proteins in the same cells expressed as a percentage of total proteolysis (top) or as a percentage of lysosomal degradation (sensitive to inhibition of lysosomal proteases) (*P = 0.021, n = 3). c, Ciliated cells in MEFs (**P = 0.003, n = 5), KECs (*P = 0.011, *P = 0.021, n = 5) and retinal ganglion cells (RGC-5; **P = 0.004, ##P = 0.0002, n = 5) maintained in the presence or absence of serum. d, Immunoblot for LC3 in MEFs and KECs maintained in the presence of serum and treated or not with protease inhibitors (PI). e, Single-channel images and merge of WT and Ift88−/− KECs transfected with the mCherry–GFP–LC3 reporter. f, Immunofluorescence for LC3 in WT and Ift88−/− KECs upon serum removal. Quantification of LC3-positive puncta (*P = 0.04, n = 3). Scale bars, 10 µm. Mean ± s.e.m. unless otherwise stated.
Extended Data Figure 3 Autophagy in IFT88 and IFT20 double-knockdown cells.
a, Immunoblot for the indicated proteins of MEFs Ctrl or knockdown for IFT88 (IFT88−) or for IFT88 and IFT20 (IFT20−/IFT88−). b, Percentage of ciliated cells after 24-h serum starvation and representative images of cilia (acetylated tubulin, green) and basal body (γ-tubulin, red) in the same cells(IFT88−, **P = 0.00001; IFT20-88−, **P = 0.0006, n = 5). Arrows indicate cilia. c, LC3 flux immunoblot in the same cell lines maintained in presence or absence of serum. d, Quantification of LC3-II levels (S+, **P = 0.0008, **P = 0.004, n = 4; S−, *P = 0.034, n = 5) and LC3-II flux (S+, n = 4; S−, *P = 0.016, n = 5) by densitometry of blots as the ones shown in c. Mean ± s.d. e, LC3 flux in a retinal ganglion cell line (RGC-5) and in KEC treated or not with platelet-derived growth factor (PDGF). Scale bars, 10 µm. Mean ± s.e.m. unless otherwise stated.
Extended Data Figure 4 Ciliary hedgehog signalling modulates autophagy.
a, Scheme of chemical and genetic approaches to modulate hedgehog (Hh) signalling. Both the agonist purmorphamine (Purmo) and genetic ablation of the Patched-1 receptor (Ptc−/−) result in the recruitment of SMO to the cilia and initiation of expression of GLI factors. Knockdown of SMO− or treatment with the SMO antagonist cyclopamine (Cyclo) suppress activation of downstream effectors. b, mRNA expression of Hh downstream effector genes relative after PDGF-induced ciliary resorption relative to untreated Serum− cells (Gli1, **P = 0.0006, **P = 0.007; Gli2, **P = 0.0001; Ptc, **P = 0.002; n = 3). c, mRNA levels for downstream target genes of Hh signalling measured by RT–PCR after the indicated treatments (Smo; Ctrl *P = 0.028, purmorphamine *P = 0.027; Gli1, ctrl **P = 0.006, purmorphamine *P = 0.037; Bcl-2, Ctrl **P = 0.007; n = 3). d, Immunofluorescence for LC3 in control cells treated or not with purmorphamine, and in Ptc−/− cells. Right, quantification (n = 3). e, mCherry–GFP–LC3 reporter in cells treated with purmorphamine. f, LC3 flux in IFT88− MEFs treated with purmorphamine. Right, quantification of LC3-II flux (Mean ± s.d.; n = 3). g, Ptc mRNA levels in MEF WT and Ptc1−/− (n = 3). h, Immunofluorescence for acetylated tubulin in MEF WT and Ptc−/−. i, Percentage ciliated cells in WT and Ptc−/− MEFs (*P = 0.014, ##P = 0.0009, n = 3). j, Relative mRNA expression by RT–PCR of Hh downstream effector genes in control and myc–GLI1 cells (Gli1, **P = 0.0017; Bcl2 **P = 0.004; Ptc **P = 0.0006; Hhip *P = 0.03; Gli2 *P = 0.028; n = 3). k, LC3 flux immunoblot in MEFs treated or not with Cyclo and PI. Quantification of LC3-II flux (n = 3). Differences with Ctrl (*) or with Serum + (#) are significant for P < 0.05. n.s., statistically non-significant. Mean ± s.e.m. unless otherwise stated. Scale bars, 10 µm.
Extended Data Figure 5 Presence of ATGs at the primary cilia.
a, Co-immunostaining for the indicated autophagy-related proteins (ATGs; green) and acetylated tubulin (red) of mouse kidney epithelial cells maintained in the absence of serum for 24 h. b, 3D reconstruction of the co-staining for ATG16L and acetylated tubulin in the cilia. ATG14 is shown an as example of absence of colocalization. 0.2-µm Z-stack are shown from the surface to the bottom part of the cilium. c, Staining with ATG16L or acetylated tubulin, and 3D images in cells transiently transfected with either GFP–LC3 or GFP–inversin and in GFP–inversin 3T3 stable cell lines. d, Immunostaining for ATG16L in NRK Ctr or knockdown for ATG16L transiently transfected with GFP–inversin to highlight the primary cilia. Individual channels, merge and 3D reconstitution of the co-staining are shown. e, Immunoblot for ATG16L in ATG16L− NRK cells. Scale bars, 10 µm.
Extended Data Figure 6 Autophagy-related proteins associate with the basal body in a serum-dependent manner.
a, b, Co-immunostaining for ATGs and gamma tubulin of kidney epithelial cells (KECs) maintained in serum-free (a) or serum-supplemented media (b). Arrows, colocalization (yellow) or no colocalization (white) in the centriole. c, Co-immunostaining of ATG7 (green) with gamma tubulin (red) in WT and Ift88−/− KECs maintained in the presence or absence of serum for 24 h. Scale bars, 10 µm.
Extended Data Figure 7 Changes in intracellular distribution of ATGs in kidney epithelial cells with defective IFT.
a, Immunostaining for ATG7, ATG14, LC3 and GABARAP in WT and Ift88−/− kidney epithelial cells (KECs) maintained in the presence or absence of serum for 24 h. b, ATG5 co-immunostaining (green) with gamma tubulin or acetylated tubulin (red) in KECs. Arrows indicate clusters of ATG5. c, Immunostaining for ATG5 in WT and Ift88−/− KECs maintained in the absence of serum for 24 h. d, Immunostaining for ATG5 (green) and acetylated tubulin (red) in WT or Ptc−/− MEFs. Yellow arrow, ATG5 over ciliary structures. Scale bars, 10 µm.
Extended Data Figure 8 Interaction of IFT20 with ATGs.
a, Co-immunostaining for ATG16L and γ-tubulin in kidney epithelial cells (KECs) WT or Ift88−/− maintained in the presence or absence of serum for 24 h. Arrows, colocalization (yellow) or no colocalization (white) at the basal body (BB). b, Percentage of WT MEFs untreated (−), treated with purmorphamine (*P = 0.01, n = 4) or Ptc−/− (*P = 0.005, n = 4) showing colocalization of ATG16L and BB in the absence of serum. c, Co-immunostaining for IFT20 and ATG16L in the same cells as in a. Insets show split channels of boxed areas at higher magnification. Arrows, colocalization. Right: quantification of the colocalization (##P = 0.004, **P = 0.004, n = 4). d, Co-immunostaining for IFT20 (red) with VPS15, ATG7 and ATG14 (green) in WT KECs during starvation. Insets show boxed areas at higher magnification. Yellow arrows, colocalization. Percentage of colocalization is indicated. e–g, Immunoblot for the indicated proteins after coimmunoprecipitation for IFT20 (e, f) or ATG14 (g) in the same cells. h, Immunogold electron microscopy for ATG16L and IFT20 in isolated cytosolic vesicles. Full-field images of double immunogold staining for IFT20 (10 nm) and ATG16L (15 nm) captured by transmission electron microscopy. ATG16L alone (yellow), IFT20 alone (blue), both proteins (red). Right image shows absence of gold particles in a region of vesicle-free film. Mean ± s.e.m. unless otherwise stated. Scale bars, 10 µm.
Extended Data Figure 9 Effect of blockage of autophagy on ciliogenesis.
a, Co-immunostaining for acetylated tubulin and gamma tubulin in MEFs from WT or ATG5 null mice (Atg5−/−) maintained in the absence of serum for the indicated periods of time. Arrows, cilia. b, Immunostaining for acetylated and gamma tubulin in MEFs control or knocked down (−) for ATG7, and maintained in the presence or absence of serum. Arrows, cilia. c, Quantification of the percentage of ciliated cells in ATG7− (n = 5). d, Immunostaining for acetylated tubulin in KEC control or ATG14−. e, Quantification of the percentage of ciliated cells in ATG14− cells. Cells from Ift88 null mice are used as negative control in e, (n = 4). f, Percentage of ciliated MEFs upon treatment with 3-methyladenine (3MA) or rapamycin (Rapa) in the presence or absence of serum (n = 3). g, Percentage of ciliated retinal ganglionar cells (RGC-5) at the indicated times of treatment with 3MA or rapamycin in the presence of serum (n = 3). Scale bars, 10 µm. Arrows; cilia. All values are mean ± s.e.m. unless otherwise stated. Differences with Ctrl (*) or with Serum+ (#) are significant for P < 0.05. n.s., statistically non-significant.
Extended Data Figure 10 Ultrastructure of the primary cilia in autophagy-deficient cells.
Scanning electron microscopy images of embryonic fibroblasts from WT and ATG5-defective mice (Atg5−/−) grown in the absence or presence of serum. a, Surface of the different cells to highlight lower levels of villi in the Atg5−/− cells. Red arrow, primary cilia. b, Details of primary cilia. Arrows, cilia-associated vesicles. Arrowhead, ciliary pocket. c–e, Morphometric analysis of the cilia; cilia diameter (S−, **P = 0.0002, n = 28; S+, **P = 0.026, n = 24) (c), area of the ciliary pocket (S−, **P = 0.009, n = 28; S+, **P = 0.0001; n = 24) (d), and exosome diameter and number of exosomes per cilia (e) in WT and Atg5−/− MEFs. Mean ± s.d.
Extended Data Figure 11 Enhanced ciliogenesis in autophagy-deficient cells.
a, Co-immunostaining for SMO and acetylated tubulin in WT and Atg5−/− MEFs treated or not with purmorphamine. Arrows, colocalization (yellow) or no colocalization (white). b, Immunofluorescence for IFT20 in MEF WT and Atg5−/−. Arrows, IFT20 cytosolic vesicles. Scale bars, 10 µm. All values are mean ± s.e.m. unless otherwise stated. c, Immunoblot for the indicated proteins in WT MEFs treated or not with ammonium chloride and leupeptin (N/L) and collected at different times after serum removal. d, Time-course of changes in IFT20 protein levels in WT and Atg5−/− cells during serum removal relative to levels in serum supplemented WT MEFs. Time-course of changes in LC3-II flux is plotted as discontinuous line.
Supplementary information
Supplementary Information
This file contains Supplementary Discussions 1-4 and additional references. (PDF 181 kb)
Localization of VPS15 at the axoneme
Serial images through the Z-stacks of the 3D reconstituted axoneme of kidney epithelial cells after 24 h in the absence of serum and co-immunostained for VPS15 (green) and acetylated tubulin (red). (MOV 4671 kb)
Localization of ATG16L at the axoneme
Serial images through the Z-stacks of the 3D reconstituted axoneme of kidney epithelial cells after 24 h in the absence of serum and co-immunostained for ATG16L (green) and acetylated tubulin (red). (MOV 3144 kb)
Localization of AMBRA1 at the axoneme
Serial images through the Z-stacks of the 3D reconstituted axoneme of kidney epithelial cells after 24 h in the absence of serum and co-immunostained for AMBRA1 (green) and acetylated tubulin (red). (MOV 4703 kb)
Localization of LC3 at the axoneme
Serial images through the Z-stacks of the 3D reconstituted axoneme of kidney epithelial cells after 24 h in the absence of serum and co-immunostained for LC3 (green) and acetylated tubulin (red). (MOV 4233 kb)
Localization of GABARAP at the axoneme
Serial images through the Z-stacks of the 3D reconstituted axoneme of kidney epithelial cells after 24 h in the absence of serum and co-immunostained for GABARAP (green) and acetylated tubulin (red). (MOV 2314 kb)
Rights and permissions
About this article
Cite this article
Pampliega, O., Orhon, I., Patel, B. et al. Functional interaction between autophagy and ciliogenesis. Nature 502, 194–200 (2013). https://doi.org/10.1038/nature12639
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature12639
This article is cited by
-
Emerging roles of prominin-1 (CD133) in the dynamics of plasma membrane architecture and cell signaling pathways in health and disease
Cellular & Molecular Biology Letters (2024)
-
Metabolism of asparagine in the physiological state and cancer
Cell Communication and Signaling (2024)
-
Abnormal accumulation of extracellular vesicles in hippocampal dystrophic axons and regulation by the primary cilia in Alzheimer’s disease
Acta Neuropathologica Communications (2023)
-
Primary cilia sense glutamine availability and respond via asparagine synthetase
Nature Metabolism (2023)
-
Ligand-dependent hedgehog signaling maintains an undifferentiated, malignant osteosarcoma phenotype
Oncogene (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
