
Abstract
The treatment of infections caused by methicillin-resistant Staphylococcus aureus is complicated by the emergence of strains with intermediate-level resistance to vancomycin (termed VISA). We have characterised a molecular pathway involved in the in vivo evolution of VISA mediated by the regulatory proteins YycH and YycI. In contrast to their function in other bacterial species, we report a positive role for these auxiliary proteins in regulation of the two-component regulator WalRK. Transcriptional profiling of yycH and yycI deletion mutants revealed downregulation of the ‘WalRK regulon’ including cell wall hydrolase genes atlA and sle1, with functional autolysis assays supporting these data by showing an impaired autolytic phenotype for each deletion strain. Using bacterial-two hybrid assays, we showed that YycH and YycI interact and that YycHI also interacts with the sensor kinase WalK, forming a ternary protein complex. Mutation to YycH or YycI associated with clinical VISA strains had a deleterious impact on the YycHI/WalK complex, suggesting that the interaction is important for the regulation of WalRK. Taken together, we have described a novel antibiotic resistance strategy for the human pathogen S. aureus, whereby YycHI mutations are selected for in vivo leading to reduced WalRK activation, impaired cell wall turnover and ultimately reduced vancomycin efficacy.
Similar content being viewed by others

Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) is a frequent cause of both hospital- and community-acquired infections and invasive disease such as bacteremia is associated with high mortality1. The increased reliance on the glycopeptide antibiotic vancomycin has resulted in the development of strains with reduced vancomycin susceptibility, termed vancomycin-intermediate S. aureus (VISA)2. VISA strains typically emerge as a result of prolonged vancomycin treatment during complex infections such as endocarditis and osteomyelitis and importantly, infections with VISA strains are associated with persistent bacteremia and poor clinical outcomes3,4.
Describing the genetic correlates contributing to VISA has remained challenging. Unlike MRSA, which emerged as a result of the horizontal acquisition of mecA encoding an alternative penicillin binding protein (PBP2a), no molecular ‘silver bullet’ is responsible for the development of VISA5. A landmark study by Mwangi et al. applied comparative genomics to a series of vancomycin-exposed clinical strains and showed that incremental increases in the minimum inhibitory concentration (MIC) of vancomycin correlated with cumulative mutations in diverse genetic pathways6. Subsequent studies have described an abundance of point mutations in regulatory loci that moderate cell wall metabolism including two-component regulatory systems (TCRS, eg. walRK, graSR and vraSR) and differential expression of the ‘cell wall stimulon’ in VISA isolates7,8,9,10,11. In addition to cell wall thickening, VISA is associated with an altered surface protein profile, enhanced capsule and agr dysfunction, each of which likely affects host-pathogen interactions12. It has been postulated that these changes contribute to immune evasion and the bacterial persistence that is observed clinically12,13.
The WalRK TCRS is a critical regulator of cell wall metabolism in S. aureus14. Upon activation by the WalK sensor histidine kinase, the WalR response regulator binds to the promoter sequence and increases the expression of multiple staphylococcal autolysins including atlA, sle1 and lytM14,15. Point mutations in the walRK operon are frequently described for VISA and have also been identified in strains resistant to a second last-line antibiotic, daptomycin16,17. In Bacillus subtilis, YycH and YycI negatively regulate the WalRK ortholog, YycFG18,19. It appears that YycHI form a complex with the sensor histidine kinase YycG, preventing it from activating its cognate response regulator, YycF18,19. Thus far, the function of YycHI in S. aureus and its contribution to the in vivo evolution of VISA is yet to be characterised.
In the present study, we have described a role for YycHI to vancomycin susceptibility in S. aureus. Chromosomal deletion of yycH and/or yycI from a clinical vancomycin-sensitive S. aureus (VSSA) strain was sufficient to produce a heterogenous vancomycin-intermediate (hVISA) phenotype. Transcriptional profiling of the yycH and yycI mutants revealed downregulation of the WalRK ‘regulon’, which was corroborated by reduced autolysis. These data are in contrast to that seen in Bacillus sp. and suggest a role for YycHI in activation of the WalRK TCRS. Using a bacterial two-hybrid approach, we have proposed that complex formation between YycHI and WalK is important for WalRK regulation and that yycHI mutation in clinical S. aureus strains contributes to VISA by disrupting this interaction thereby reducing WalRK activity and impairing cell wall turnover.
Results
Genetic analysis of yycHI mutations in clinical VISA strains
We previously performed whole-genome sequencing of multiple, clinically-derived VISA isolates to characterise the genetic mechanisms of the in vivo evolution of reduced susceptibility to vancomycin in S. aureus9. We identified a novel deletion mutation within yycI, which belongs to the walRK/yycHIJ operon. In S. aureus, this operon consists of walR, which codes for a response regulator, walK, which codes for a sensor histidine kinase, yycH and yycI, which encode putative regulatory proteins and yycJ, which codes for a 5′-3′ exonuclease involved in mismatch repair20 (Fig. 1A). The yycI mutation was one of 13 found between the VSSA progenitor strain A6224 and VISA strain A6226 that emerged after prolonged vancomycin treatment9. The four-nucleotide deletion (ATAA from the position 79–82) resulted in a predicted frameshift from the 27th amino acid and truncation of the protein from 262 amino acids in the VSSA strain (A6224) to 56 amino acids in the VISA strain (A6226) (Fig. 1B). No mutation was observed for the upstream protein, YycH (Fig. 1C). Intriguingly, in a separate study, Mwangi et al. identified 35 mutations between VISA strain JH9 (A8094 in this study) and the VSSA progenitor strain JH1 (A8090 in this study) including a single nucleotide polymorphism (SNP) in yycH6. The G to A substitution at the 81st nucleotide base was predicted to change the tryptophan at position 27 to a premature stop codon, truncating the protein to ~5% of its original length (453 amino acids) (Fig. 1C). No YycI mutation was observed in the VISA strain (A8094) (Fig. 1B). DNA sequencing confirmed the yycHI mutations in A6226 and A8094. Given the degree of truncation and the predicted role for the proteins in WalRK regulation, we hypothesized that yycHI mutation likely contributes to reduced vancomycin susceptibility in clinical VISA strains.

Genetic organisation of the walRK operon in S. aureus.
(A) The walRK two-component regulatory system is encoded directly upstream of yycH, yycI and yycJ. Predicted transmembrane domains are presented in black. (B) A 262 amino acid protein is coded for by the consensus sequence of yycI in S. aureus. Mutation to yycI in clinical VISA isolate A6226 generates a frameshift (black box) and truncation of the protein to 56 amino acids. (C) A 453 amino acid protein is coded for by the consensus sequence of yycH in S. aureus. A single nucleotide polymorphism in clinical VISA strain A8094 generates a premature stop codon at the 27th amino acid (black box). Black lines represent predicted transmembrane domains for the consensus sequence of each protein. Grey lines represent native residues that remain in each in-frame deletion strain.
YycH and YycI influence vancomycin susceptibility
To determine the exact contribution of yycHI to the development of VISA independent of other mutations, chromosomal deletions of yycH (A8090ΔyycH), yycI (A8090ΔyycI) and yycHI (A8090ΔyycHI) were generated within the genetic context of VSSA clinical isolate, A8090. Vancomycin susceptibility was assessed using Etest and population analysis profiling (PAP). A8090ΔyycH and A8090ΔyycI each showed a reduction in vancomycin susceptibility, with a MIC of vancomycin increasing from 1.0 μg/mL to a maximum of 3.0 μg/mL (range 2.0–3.0 µg/ml) for both mutants, which is in the non-susceptible range (>2 μg/mL) (Table 1) and has been associated with poor patient outcomes21. Notably, the MIC for vancomycin of the double mutant (A8090ΔyycHI) was no greater (Table 1) than for each of the single mutants, suggesting that both YycH and YycI proteins are equally essential for vancomycin susceptibility. PAP analyses are the gold standard for assessing VISA and heterogeneous VISA (hVISA) and provide a more sensitive quantification of resistant sub-populations. Each deletion strain had a greater number of cells tolerant to >2 μg/mL of vancomycin (Fig. 2), which defines the VISA phenotype and predicts for persistent bacteremia and complicated infection outcomes3,22.

Vancomycin susceptibility profiles of A8090 and yycHI mutant strains.
Vancomycin heteroresistance was assessed for A8090, deletion mutants (ΔyycH, ΔyycI and ΔyycHI) and gene reconstituted strains (::yycH and::yycI) using population analysis profiling on heart infusion agar supplemented with increasing concentrations of vancomycin. Data are expressed as mean ± SEM.
Genetic reconstitution of yycH and yycI in situ returned the MIC for vancomycin to within the susceptible range (1.0 μg/mL, Table 1) and each reconstituted strain produced a similar vancomycin population profile to that of the vancomycin-susceptible parent strain (Fig. 2). Together, these data confirm that both YycH and YycI are important for vancomycin susceptibility in S. aureus.
Deletion of yycH or yycI from VSSA strain A8090 did not recreate the MIC of vancomycin for the clinical VISA isolate A8094 (8.0 μg/mL). This suggests that additional mutations in A8094 contribute to the VISA phenotype for this strain and supports the theory that VISA emerge as a result of cumulative mutations during prolonged vancomycin exposure. In addition, the in vivo evolution of VISA in A8094 and A6226 was associated with co-resistance to daptomycin in the absence of daptomycin exposure (Table 1). To determine if yycHI mutation contributed to the daptomycin-nonsusceptible phenotype of these isolates, daptomycin susceptibility testing was performed including PAP analyses. No change in susceptibility was observed for A8090ΔyycH or A8090ΔyycI when compared to A8090 (Table 1, PAP data not shown). It is likely that mutations present in A8094 and A6226 other than those in yycHI contributed to daptomycin resistance in these isolates.
YycHI positively regulates WalRK in S. aureus
To determine how YycHI contributes to antibiotic susceptibility and to understand the impact of these proteins upon WalRK, we compared the transcriptomes of A8090, ΔyycH and ΔyycI using RNA-Seq. Transcriptomes were sequenced using the Illumina sequencing platform yielding an average total read count of 6.67 million reads per sample. The transcriptomes of ΔyycH and ΔyycI were highly similar (only six differentially expressed genes representing 0.2% of the transcriptome [Supplementary Table S1]), suggesting that both proteins function within the same system and are equally important in S. aureus physiology (Fig. 3). When the transcriptome of ΔyycH or ΔyycI was compared to the progenitor strain A8090, a total of 42 genes showed altered expression (≥2-fold) in at least one of the deletion strains. These included 17 downregulated genes (Supplementary Table S2) and 25 upregulated genes (Supplementary Table S3). When each differentially expressed gene was grouped based on Clusters of Orthologous Groups (COG) or Kyoto encyclopedia of genes and genomes (KEGG) pathways, 40% of the genes were predicted to be involved in cell wall, membrane and envelope biosynthesis (Fig. 3A). Furthermore, 21% of genes were predicted to be involved in staphylococcal infection and defence (Fig. 3A). Quantitative digital PCR of select genes confirmed these RNA-Seq data (Supplementary Fig. S1).

Transcriptional comparison of yycH and yycI deletion mutants.
(A) RNA sequencing was used to generate lists of genes with differential expression in ΔyycH and ΔyycI compared to parental strain A8090. Each gene was assigned a cluster of orthologous groups (COG) category or Kyoto encyclopedia of genes and genomes (KEGG) pathway category (represented by *). Genes upregulated in each deletion strain are represented in red, whilst genes downregulated in each deletion strain are represented in green. Transcriptional profiles of ΔyycH and ΔyycI were highly similar. (B) Genes predicted to be regulated by WalRK are largely downregulated in ΔyycH and ΔyycI. Genes confirmed to be under the transcriptional control of WalRK are denoted by #14,24,46. Remaining genes are predicted to be under the control of WalRK due to the presence of an upstream putative WalR binding site14,46. (C) Genes involved in host-pathogen interaction were differential expressed in ΔyycH and ΔyycI. Genes encoding surface proteins, adhesins and complement inhibitors were mostly downregulated, whilst the capsule operon was strongly overexpressed in ΔyycH and ΔyycI.
WalR is a critical transcriptional regulator in S. aureus and has been shown to influence the expression of genes involved in cell wall turnover, which explains its role in the development of VISA, as well as a diverse range of genes important for amino acid biosynthesis, central metabolism and virulence14,15,16. However, experimental confirmation of WalR binding to the promoter sequence of specific genes and induction of their expression has only been confirmed for five genes, each of which encode for proteins with autolytic activity (sle1, atlA, ssaA, isaA and lytM)14,15. Based on work in Bacillus, we hypothesised that YycHI binds to WalK and alters its regulatory activity18,19. To determine the direction of YycHI-mediated regulation of WalRK in S. aureus, we interrogated our RNA-seq data for the expression of validated WalR regulated genes. We showed that except for lytM, the remaining four genes (sle1, atlA, ssaA and isaA) were downregulated in both the ΔyycH and ΔyycI deletion mutants when compared to their progenitor strain (Fig. 3B and Supplementary Table S2). Furthermore, whilst upstream WalR binding is yet to be confirmed, walRK induction increases the expression of four CHAP-domain containing amidase/peptidoglycan hydrolase genes (SaurJH1_0704, SaurJH1_0796, SaurJH1_2370 and SaurJH1_2642)14. Of these, SaurJH1_0704 and SaurJH1_2370 were significantly downregulated in ΔyycH and ΔyycI (Fig. 3B and Supplementary Table S2). We also showed, similar to that observed in Bacillus19, that YycH has a growth-phase dependent impact on gene expression that is most pronounced at early stationary compared to mid-exponential phase (Supplementary Fig. S2A). These transcriptomic changes suggest that in contrast to that seen in Bacillus sp., YycHI positively regulates the WalRK TCRS in S. aureus. Importantly, the transcription of walRK was not different for either ΔyycH or ΔyycI when compared to the progenitor strain A8090.
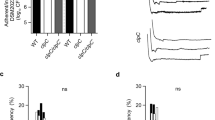
To provide further support to the direction of WalRK regulation by YycHI in S. aureus, we performed functional autolysis assays. It is well established that WalRK is a regulator of autolysis in S. aureus14 and this is supported by the five validated WalR-regulated genes that all encode proteins with autolytic activity14,15. Using Triton X-100 induction, we compared the autolytic rate of ΔyycH and ΔyycI to their progenitor, A8090. Each deletion strain showed impaired autolysis as determined by a reduced decline in OD600 over time (Fig. 4). Gene reconstitution restored the autolytic rate back to that of the vancomycin-susceptible parent strain (A8090) (Fig. 4). Together, the observed differential gene expression and the functional impact on autolysis suggests that YycHI acts as a positive regulator of WalRK in S. aureus. Deletion of either YycH or YycI leads to downregulation of WalR-regulated genes and WalR-mediated autolysis, which likely contributes to reduced susceptibility to the cell-wall acting antibiotic, vancomycin.

Deletion of yycH or yycI reduces autolysis.
Autolysis was determined as the decline in OD600 over time at 30 °C in the presence of Triton X-100. Data are expressed as mean ± SEM (**P < 0.01, *P < 0.05).
Transcriptional effects of yycHI deletion upon genes associated with host-pathogen interaction
In addition to regulating cell wall turnover, recent reports have shown that WalRK indirectly controls the expression of a number of virulence factors via activation of the SaeSR TCRS15. Fittingly, our transcriptomic data highlighted down-regulation of multiple genes that are involved in host-pathogen interaction, a number of which are indirectly regulated by WalR (Fig. 3C)15. These included immunoglobulin-binding proteins encoded by spa and sbi, which were downregulated in both ΔyycH and ΔyycI. In addition, fibrinogen-binding proteins encoded by efb and SaurJH1_1235 were less abundant in ΔyycH and ΔyycI and excreted factors scn and SaurJH1_1240 encoding staphylococcal complement inhibitors were significantly downregulated in ΔyycI (Supplementary Table S2). In contrast, the entire capsule operon was overexpressed in both ΔyycH and ΔyycI (Fig. 3C), which is commonly reported for VISA strains12.
YycH, YycI and WalK form a ternary protein complex
Thus far little is known about the staphylococcal YycH and YycI proteins and their interactions. Work in B. subtilis has showed that YycH and YycI are tethered to the cytoplasmic membrane and physically interact at their respective transmembrane domains (TMDs)18. Using a bacterial-two hybrid system, we assessed the interactions between YycH and YycI from S. aureus. Full-length YycH was found to interact with full-length YycI using this approach (Fig. 5A). We observed that except for the TMDs of each protein (YycI1–53 and YycH1–36), the remainder of the protein was dispensable for their interaction (Fig. 5A). YycH/YycI constructs lacking TMDs did not interact with their respective full-length binding partners (Fig. 5A). These data suggest that staphylococcal YycHI interactions are mediated by the TMDs.

YycH, YycI and WalK interactions as determined by bacterial-two-hybrid analysis.
(A) β-galactosidase assays were performed for E. coli co-expressing bacterial two-hybrid fusions of full-length YycH (H) and YycI (I) as well as fragments containing the transmembrane domain of each protein (H1–36 and I1–53) or lacking each respective transmembrane domain (H40–444 and I36–262). (B) β-galactosidase assays were also performed for E. coli expressing combinations of WalK (K), YycH (H), YycI (I) and YycHI (HI). (C) YycH mutation associated with VISA strain A8094 (YycHA8094) reduced the strength of interaction with YycI and YycI mutation associated with VISA strain A6226 (YycIA6226) reduced the strength of interaction with YycH as determined by β-galactosidase activity. (D) Similarly, mutations associated with A8094 (YycHIA8094) and A6226 (YycHIA6226) reduced the strength of the complex formed between YycH, YycI and WalK. No interaction controls were generated using pKT25 or pUT18C without gene inserts (represented by -). Data are presented as mean ± SEM (*P < 0.05).
To determine whether YycH/YycI interact with the sensor kinase WalK, we first tested them individually. Similar to our observations for vancomycin susceptibility, YycH or YycI alone were not sufficient to produce a quantifiable interaction with WalK (Fig. 5B), suggesting that YycH and YycI are required as a protein complex before they can adequately interact with WalK. To test this hypothesis, full-length YycH, YycI and WalK were co-expressed in Escherichia coli BTH101; a quantifiable interaction was observed (Fig. 5B).
yycHI mutations associated with clinical VISA strains disrupt the interaction between YycH, YycI and WalK
In addition to yycHI mutations described in this report, numerous diverse walK point mutations have been described in VISA strains16,17. To date, it is unclear how these mutations impact on the activity of WalRK or the relationship with YycHI. As such, we next investigated the impact of VISA associated walK/yycHI mutations upon the interaction between YycH, YycI and WalK using our bacterial two-hybrid system. WalK proteins containing amino acid substitutions associated with diverse VISA isolates were assessed including G233D from JKD6008, Q371Δ from A8392 and R263C/S273N from A8118 (Table 1). These WalK mutations had no effect on the interaction with YycHI (data not shown). We next assessed the impact of yycHI mutations from the clinical VISA strains A8094 and A6226 (Fig. 1). The interaction between YycH and YycI was reduced 6-fold when the mutated YycH from A8094 was assessed and the interaction was abolished when the mutated YycI from A6226 was included in the analysis (Fig. 5C). Most importantly, the interaction with the kinase, WalK was significantly reduced when mutated YycH or YycI proteins from either A8094 or A6226 were included (Fig. 5D). Together, these data suggest that reduced vancomycin susceptibility in our clinical VISA strains is due to impaired interaction of the YycHI protein complex with WalK, leading to reduced expression of the WalR regulon and reduced WalR-mediated autolysis. A proposed model for this is shown in Fig. 6.

Proposed model for the impact of YycHI mutation upon the activation of WalKR in the context of VISA.
(A) In vancomycin-sensitive strains, YycH and YycI physically interact with the sensor kinase to activate the WalRK TCRS. WalR is phosphorylated by WalK leading to transcription of target genes. (B) Mutation to either YycH or YycI occurring in clinical VISA strains has a negative impact on complex formation, resulting in reduced expression of WalR-regulated genes via an unknown mechanism. Reduced WalR activity contributes to reduced autolysis, impaired cell wall turnover and ultimately reduced vancomycin (VAN) susceptibility.
Discussion
Defining the complete molecular pathway to reduced vancomycin susceptibility in S. aureus has remained challenging owing to the heterogeneous nature of the genetic mutations associated with VISA. We and others have used whole-genome sequencing of clinical isolates to guide targeted genetic manipulation experiments to determine the exact contribution of select genes to reduced susceptibility to vancomycin, independent of other mutations9,16. By engineering yycH and yycI deletions from a vancomycin-susceptible strain, we have described for the first time the contribution of these genes to the development of reduced vancomycin susceptibility and we confirmed their importance by restoring the vancomycin-susceptible phenotype via gene reconstitution (Fig. 2).
The function of YycH and YycI in S. aureus is typically inferred from their role in Bacillus sp. where they negatively regulate the WalRK ortholog YycFG18,19. WalRK/YycFG is highly conserved across low GC content gram-positive bacteria and is essential in S. aureus most likely due to its crucial role in regulating cell wall metabolism23,24. Due to its role in cell wall maintenance, it is not surprising that WalRK mutation is commonly described for VISA16,17. Two distinct mechanisms have been identified that infer a role for WalRK in the evolution of VISA. Firstly, IS256 insertion in the promoter of the operon has been shown to reduce vancomycin susceptibility and secondly, point mutations within walK and walR contribute to vancomycin tolerance in clinical VISA strains16,25,26. What remains unclear, however, is the impact of these mutations upon the activity of WalRK. Jansen et al. showed that IS256 insertion led to increased expression of walRK whereas more recently, McEvoy et al. showed that upstream IS256 insertion decreased expression of the TCRS25,26. Similarly, transcriptional profiling of VISA-associated walK and walR point mutants revealed no clear directionality for the activity of the TCRS based on contrasting expression of isaA and atlA, each of which are induced by WalR16. In this study, we have described a third mechanism of WalRK-mediated VISA development, via mutation to auxiliary proteins YycHI. Along with reduced vancomycin susceptibility, deletion of yycH or yycI resulted in reduced autolysis and downregulation of genes under the transcriptional control of WalRK, including atlA, sle1 and CHAP domain amidase/peptidoglycan hydrolase genes SaurJH1_0704 and SaurJH1_2370 (Fig. 3B). Taken together these data suggest that in contrast to Bacillus sp., YycHI serves as an activator of WalRK in S. aureus and that the VISA phenotype associated with yycHI mutation correlates to reduced activity of the system, as opposed to enhanced activity, which was previously proposed6,9,27.
The regulatory impact of YycHI in B. subtilis was found to be as a result of direct interaction with the kinase, YycG18,19. A report investigating YycH and YycI from S. aureus failed to provide evidence of an interaction with WalK using a detergent-micelle-model28. Here, we have used a bacterial two-hybrid system to show that YycH or YycI in isolation was not sufficient to produce an interaction with WalK but when all three proteins were present, a ternary protein complex formed (Fig. 5B). This correlates with what has been described for B. subtilis using immunoprecipitation assays, whereby YycH only co-precipitated with YycG when YycI was present and vice versa29. YycH and YycI appear indispensible for the regulation of WalK, which explains why minimal transcriptional or phenotypic differences were observed between the yycH, yycI and yycHI deletion strains in this study. In prokaryotes, a number of TCRSs are regulated by interaction with auxiliary proteins. For example, the PII protein from E. coli acts as a repressor of NtrBC by blocking the autokinase domain and enhancing the phosphatase activity of the sensor kinase NtrB30. In addition, SaePQ serves as a repressor of the staphylococcal virulence regulator SaeSR by activating the phosphatase activity of the sensor kinase SaeS31. Whilst we have shown a physical interaction between YycHI and WalK, further experiments are required to understand exactly how YycHI serves to regulate WalRK in S. aureus.
The TMDs of YycH and YycI appeared to be important for protein-protein interactions (Fig. 5A). The yycH mutation associated with the clinical VISA strain, A8094 resulted in TMD truncation, which impacted on the interaction with YycI, as well as complex formation with WalK (Figs 1 and 5). Similarly, the yycI mutation associated with the clinical VISA strain, A6226 resulted in a frameshift within the TMD that led to similar effects upon interaction with YycH and WalK (Figs 1 and 5). Thus, based on current evidence, yycHI VISA mutations impact on WalRK activation as a direct result of reduced interaction with the sensor kinase WalK.
When compared to infections caused by VSSA, VISA are more likely to be associated with prolonged bacteremia and less likely to cause acute clinical instability such as septic shock32,33. In the current study, whilst we did not observe altered expression of major staphylococcal toxins, we did observe differential transcription of genes that impact on interaction with the host (Fig. 3C). Surface protein A (coded for by spa) is a trigger for platelet aggregation, which stimulates the production of platelet microbiocidal proteins34. Deceased expression of spa, as seen for our yycH/I deletion strains, likely results in reduced stimulation of platelet microbiocidal proteins that are important for host-defense against S. aureus bloodstream infection. As a corollary, increased expression of the capsule operon contributes to resistance to opsinophagocytosis by human polymorphonuclear leukocytes and enhanced bacterial persistence in a murine model of S. aureus infection35.
In keeping with the notion of YycHI as an activator of WalRK, the expression of Ig-binding B domain containing protein (sbi), complement inhibitors (scn, SaurJH1_1240) and fibrinogen binding proteins (efb, SauJH1_1235) were downregulated in this study, each of which have been shown to be positively regulated upon WalR overexpression15. Constitutive activation of WalR has also been shown to promote bacterial clearance and increase neutrophil recruitment in vivo15. Taken together, it is plausible that mutations to yycHI and reduced WalRK activation may contribute to a ‘stealth’ strategy coined by Gardete et al. whereby VISA alter the expression of virulence determinants to facilitate immune evasion and persistence within the context of infection27; a hypothesis that requires further exploration.
In summary, our data highlight a novel role for YycH and YycI to the development of reduced vancomycin susceptibility in S. aureus. YycHI mutations associated with VISA directly impacted upon interaction with the sensor kinase WalK and yycH/yycI deletion resulted in reduced expression of genes under its control. This indicates a positive role for YycHI proteins in regulating the WalRK TCRS in S. aureus.
Methods
Bacterial strains, plasmids and culture conditions
S. aureus strains were grown in Heart Infusion (HI) broth or agar and are listed in Table 1. E. coli strains were grown in LB broth or agar and are listed in Table S4. For the selection of strains carrying plasmids (listed in Table S4), media was supplemented with ampicillin (100 μg/mL), carbenicillin (100 μg/mL), kanamycin (50 μg/mL) or chloramphenicol (25 μg/mL).
Genetic manipulation
The E. coli/S. aureus shuttle vector pKOR1 was used to generate in-frame deletions of yycH (A8090ΔyycH), yycI (A8090ΔyycI) and yycHI (A8090ΔyycHI) as described previously36. Approximately one kilobase up- and down-stream of each gene was amplified using primers listed in Table S4. Products were cloned into pKOR1 using BP Clonase (Invitrogen), then used to transform E. coli DH5α (NEB). Each plasmid was passaged through S. aureus strain RN4220 before being electroporated into A809037. Integration of the vector was stimulated by growing cells in the presence of chloramphenicol at 42 °C followed by vector excision at 30 °C (no antibiotic). Counter-selection was achieved using 1 μg/mL anhydrotetracycline (ATc).
Gene reconstitution in situ was performed for A8090ΔyycH and A8090ΔyycI using the E. coli/S. aureus shuttle vector pIMAY38. Silent PvuI restriction sites were engineered into each reconstituted gene using splicing with overlap extension PCR (SOE-PCR)38,39. The reconstituted genes were introduced into pIMAY, generating pIMAY::yycH and pIMAY::yycI (Table S4) and the vectors were electroporated into A8090ΔyycH and A8090ΔyycI, respectively. Integration, plasmid excision and counter-selection were performed as described38 generating A8090::yycH and A8090::yycI (Table 1). To distinguish each strain from A8090, a 3.8 kb fragment encompassing yycHI was amplified from A8090, A8090::yycH and A8090::yycI and the products were digested with PvuI. A8090 produced one fragment whereas A8090::yycH and A8090::yycI each produced two (data not shown).
Antibiotic susceptibility testing and autolysis assays
Minimum inhibitory concentrations (MICs) were determined using Etest (bioMérieux) as per manufacturer’s specifications. To assess for antibiotic heteroresistance, population analysis profiling was performed using the method of Wootton et al.9,40.
Autolytic activity was assessed using Triton X-100 induction41. S. aureus cells were grown to an optical density at 600 nm (OD600) of 0.8, chilled then pelleted by centrifugation. Cells were washed once with PBS then resuspended in PBS (pH 7.4) supplemented with 0.05% Triton X-100 to an OD600 of 1.0. Autolysis was defined as the decline in OD600 over time.
RNA extraction, sequencing and digital PCR
For RNA sequencing, total RNA was extracted from early stationary phase (OD600 7.0, Fig. S2B), as the impact of YycHI upon WalR-dependent gene expression in Bacillus sp. was most pronounced at this growth stage19. RNA was stabilized in situ using RNAlater (QIAGEN) and samples were frozen at −80 °C. S. aureus cells were thawed at ambient temperature then disrupted using a Precellys 24 tissue homogenizer (Bertin Technologies). RNA was purified using an RNeasy mini kit (QIAGEN), with on-column DNAse I (QIAGEN) digestion. RNA from two independent extractions was forwarded to the Beijing Genomics Institute for sequencing using Illumina HiSeq 2000. Reads were mapped to the A8090 genome (GenBank accession NC_009632) and gene expression was quantified using the Reads Per Kilobase Per Million (RPKM) method42. Differential gene expression between strains was determined using NOISeq43. A fold change ratio of >2.0 with a q-value ≥0.8 was considered significant as described elsewhere43. To validate the differential gene expression data generated by RNA sequencing, digital PCR was performed for sle1, SaurJH1_2073, capL, spa and efb (primers listed in Table S5). cDNA (500 ng) was synthesized using Superscript III reverse transcriptase (Invitrogen). Digital PCR was performed as described by the manufacturer (Bio-Rad) for three biological replicates and gene expression was normalized against gyrB expression.
Bacterial two-hybrid (BTH) analysis
Bacterial two-hybrid vectors were constructed and assays performed as described previously (Table S4)44. Coding sequences of proteins of interest were fused to either T25 (pKT25) or T18 (pUT18C) of adenylate cyclase. Each construct was verified by DNA sequencing. To determine if protein fragments interacted, combinations of pKT25 and pUT18C based plasmids were co-transformed into E. coli strain BTH101, which is adenylate cyclase deficient. Interaction was observed via functional complementation of adenylate cyclase and expression of the lac reporter44. The relative strength of interaction was quantified by measuring β-galactosidase activity using o-nitrophenol-β-galactoside (ONPG) as a substrate, as described elsewhere45. β-galactosidase assays were performed in triplicate for 4 biological replicates. Significance was determined by Mann-Whitney U test with a significance level of P < 0.05.
Published during review process
While this paper was being reviewed, a manuscript was published by Poupel O et al. (Poupel et al., 2016 doi: 10.1371/journal.pone.0151449) that supported the key findings from this study. Deletion of yycHI from the S. aureus HG001 genetic background (vancomycin MIC 3 µg/ml) resulted in a decrease in vancomycin susceptibility (MIC 4 µg/ml) which was restored upon complementation. The authors concluded that the YycHI complex does not act as a repressor of WalRK in S. aureus by analyzing autolysis and the expression of atlA, sle and saouhsc00773 in yycH and yycI deletion mutants during the exponential phase of growth (OD 600 nm 1.0). By assessing global gene expression at a later point in the growth cycle (OD 600 nm 7.0) we have now shown that in contrast to Bacillus, YycHI induces the expression of genes under the transcriptional control of WalRK in S. aureus.
Additional Information
How to cite this article: Cameron, D. R. et al. Vancomycin susceptibility in methicillin-resistant Staphylococcus aureus is mediated by YycHI activation of the WalRK essential two-component regulatory system. Sci. Rep. 6, 30823; doi: 10.1038/srep30823 (2016).
References
Cosgrove, S. E. et al. Comparison of mortality associated with methicillin-resistant and methicillin-susceptible Staphylococcus aureus bacteremia: a meta-analysis. Clin. Infect. Dis. 36, 53–59 (2003).
Howden, B. P., Davies, J. K., Johnson, P. D., Stinear, T. P. & Grayson, M. L. Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin-intermediate and heterogeneous vancomycin-intermediate strains: resistance mechanisms, laboratory detection and clinical implications. Clin. Microbiol. Rev. 23, 99–139 (2010).
Charles, P. G., Ward, P. B., Johnson, P. D., Howden, B. P. & Grayson, M. L. Clinical features associated with bacteremia due to heterogeneous vancomycin-intermediate Staphylococcus aureus. Clin. Infect. Dis. 38, 448–451 (2004).
Howden, B. P. et al. Treatment outcomes for serious infections caused by methicillin-resistant Staphylococcus aureus with reduced vancomycin susceptibility. Clin. Infect. Dis. 38, 521–528 (2004).
Hartman, B. J. & Tomasz, A. Low-affinity penicillin-binding protein associated with beta-lactam resistance in Staphylococcus aureus. J. Bacteriol. 158, 513–516 (1984).
Mwangi, M. M. et al. Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proc. Natl. Acad. Sci. USA 104, 9451–9456 (2007).
Cui, L., Neoh, H. M., Shoji, M. & Hiramatsu, K. Contribution of vraSR and graSR point mutations to vancomycin resistance in vancomycin-intermediate Staphylococcus aureus. Antimicrob. Agents Chemother. 53, 1231–1234, 10.1128/AAC.01173-08 (2009).
Cui, L. et al. An RpoB mutation confers dual heteroresistance to daptomycin and vancomycin in Staphylococcus aureus. Antimicrob. Agents Chemother. 54, 5222–5233 (2010).
Cameron, D. R. et al. Serine/Threonine Phosphatase Stp1 Contributes to Reduced Susceptibility to Vancomycin and Virulence in Staphylococcus aureus. J. Infect. Dis. 205, 1677–1687, 10.1093/infdis/jis252 (2012).
Howden, B. P. et al. Genomic analysis reveals a point mutation in the two-component sensor gene graS that leads to intermediate vancomycin resistance in clinical Staphylococcus aureus. Antimicrob. Agents Chemother. 52, 3755–3762 (2008).
McAleese, F. et al. Overexpression of genes of the cell wall stimulon in clinical isolates of Staphylococcus aureus exhibiting vancomycin-intermediate- S. aureus-type resistance to vancomycin. J. Bacteriol. 188, 1120–1133 (2006).
Howden, B. P. et al. Different bacterial gene expression patterns and attenuated host immune responses are associated with the evolution of low-level vancomycin resistance during persistent methicillin-resistant Staphylococcus aureus bacteraemia. BMC Microbiol 8, 39 (2008).
Cameron, D. R., Howden, B. P. & Peleg, A. Y. The interface between antibiotic resistance and virulence in Staphylococcus aureus and its impact upon clinical outcomes. Clin. Infect. Dis. 53, 576–582, 10.1093/cid/cir473 (2011).
Dubrac, S., Boneca, I. G., Poupel, O. & Msadek, T. New insights into the WalK/WalR (YycG/YycF) essential signal transduction pathway reveal a major role in controlling cell wall metabolism and biofilm formation in Staphylococcus aureus. J. Bacteriol. 189, 8257–8269 (2007).
Delaune, A. et al. The WalKR system controls major staphylococcal virulence genes and is involved in triggering the host inflammatory response. Infect. Immun. 80, 3438–3453, 10.1128/IAI.00195-12 (2012).
Howden, B. P. et al. Evolution of multidrug resistance during Staphylococcus aureus infection involves mutation of the essential two component regulator WalKR. PLoS Pathog 7, e1002359, 10.1371/journal.ppat.1002359 (2011).
Shoji, M. et al. walK and clpP Mutations Confer Reduced Vancomycin Susceptibility in Staphylococcus aureus. Antimicrob. Agents Chemother. 55, 3870–3881 (2011).
Szurmant, H., Mohan, M. A., Imus, P. M. & Hoch, J. A. YycH and YycI interact to regulate the essential YycFG two-component system in Bacillus subtilis. J. Bacteriol. 189, 3280–3289 (2007).
Szurmant, H., Bu, L., Brooks, C. L., 3rd & Hoch, J. A. An essential sensor histidine kinase controlled by transmembrane helix interactions with its auxiliary proteins. Proc. Natl. Acad. Sci. USA 105, 5891–5896, 10.1073/pnas.0800247105 (2008).
Yang, H. et al. Evidence that YycJ is a novel 5′-3′ double-stranded DNA exonuclease acting in Bacillus anthracis mismatch repair. DNA repair 12, 334–346, 10.1016/j.dnarep.2013.02.002 (2013).
van Hal, S. J., Lodise, T. P. & Paterson, D. L. The clinical significance of vancomycin minimum inhibitory concentration in Staphylococcus aureus infections: a systematic review and meta-analysis. Clin. Infect. Dis. 54, 755–771, 10.1093/cid/cir935 (2012).
Casapao, A. M. et al. Clinical outcomes in patients with heterogeneous vancomycin-intermediate Staphylococcus aureus (hVISA) bloodstream infection. Antimicrob. Agents Chemother. 10.1128/AAC.00380-13 (2013).
Dubrac, S., Bisicchia, P., Devine, K. M. & Msadek, T. A matter of life and death: cell wall homeostasis and the WalKR (YycGF) essential signal transduction pathway. Mol. Microbiol. 70, 1307–1322, 10.1111/j.1365-2958.2008.06483.x (2008).
Delaune, A. et al. Peptidoglycan crosslinking relaxation plays an important role in Staphylococcus aureus WalKR-dependent cell viability. PLoS One 6, e17054, 10.1371/journal.pone.0017054 (2011).
Jansen, A. et al. Role of insertion elements and yycFG in the development of decreased susceptibility to vancomycin in Staphylococcus aureus. Int. J. Med. Microbiol. 297, 205–215 (2007).
McEvoy, C. R. et al. Decreased vancomycin susceptibility in Staphylococcus aureus caused by IS256 tempering of WalKR expression. Antimicrob. Agents Chemother. 57, 3240–3249, 10.1128/AAC.00279-13 (2013).
Gardete, S. et al. Genetic pathway in acquisition and loss of vancomycin resistance in a methicillin resistant Staphylococcus aureus (MRSA) strain of clonal type USA300. PLoS Pathog 8, e1002505, 10.1371/journal.ppat.1002505 (2012).
Turck, M. & Bierbaum, G. Purification and activity testing of the full-length YycFGHI proteins of Staphylococcus aureus. PLoS One 7, e30403, 10.1371/journal.pone.0030403 (2012).
Fukushima, T., Szurmant, H., Kim, E. J., Perego, M. & Hoch, J. A. A sensor histidine kinase co-ordinates cell wall architecture with cell division in Bacillus subtilis. Mol. Microbiol. 69, 621–632, 10.1111/j.1365-2958.2008.06308.x (2008).
Ninfa, A. J. & Jiang, P. PII signal transduction proteins: sensors of alpha-ketoglutarate that regulate nitrogen metabolism. Curr. Opin. Microbiol. 8, 168–173, 10.1016/j.mib.2005.02.011 (2005).
Jeong, D. W. et al. The auxiliary protein complex SaePQ activates the phosphatase activity of sensor kinase SaeS in the SaeRS two-component system of Staphylococcus aureus. Mol. Microbiol. 86, 331–348, 10.1111/j.1365-2958.2012.08198.x (2012).
Yoon, Y. K., Kim, J. Y., Park, D. W., Sohn, J. W. & Kim, M. J. Predictors of persistent methicillin-resistant Staphylococcus aureus bacteraemia in patients treated with vancomycin. J. Antimicrob. Chemother. 65, 1015–1018, 10.1093/jac/dkq050 (2010).
Soriano, A. et al. Influence of vancomycin minimum inhibitory concentration on the treatment of methicillin-resistant Staphylococcus aureus bacteremia. Clin. Infect. Dis. 46, 193–200, 10.1086/524667 (2008).
Yeaman, M. R. & Bayer, A. S. Antimicrobial peptides from platelets. Drug resistance updates: reviews and commentaries in antimicrobial and anticancer chemotherapy 2, 116–126, 10.1054/drup.1999.0069 (1999).
Luong, T. T. & Lee, C. Y. Overproduction of type 8 capsular polysaccharide augments Staphylococcus aureus virulence. Infect. Immun. 70, 3389–3395 (2002).
Bae, T. & Schneewind, O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid 55, 58–63 (2006).
Lee, J. C. Electrotransformation of Staphylococci. Methods Mol. Biol. 47, 209–216, 10.1385/0-89603-310-4:209 (1995).
Monk, I. R., Shah, I. M., Xu, M., Tan, M. W. & Foster, T. J. Transforming the untransformable: application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. mBio 3, 10.1128/mBio.00277-11 (2012).
Ho, S. N., Hunt, H. D., Horton, R. M., Pullen, J. K. & Pease, L. R. Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene 77, 51–59 (1989).
Wootton, M. et al. A modified population analysis profile (PAP) method to detect hetero-resistance to vancomycin in Staphylococcus aureus in a UK hospital. J. Antimicrob. Chemother. 47, 399–403 (2001).
Passalacqua, K. D., Satola, S. W., Crispell, E. K. & Read, T. D. A mutation in the PP2C phosphatase gene in a Staphylococcus aureus USA300 clinical isolate with reduced susceptibility to vancomycin and daptomycin. Antimicrob. Agents Chemother. 56, 5212–5223, 10.1128/AAC.05770-11 (2012).
Mortazavi, A., Williams, B. A., McCue, K., Schaeffer, L. & Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nature methods 5, 621–628, 10.1038/nmeth.1226 (2008).
Tarazona, S., Garcia-Alcalde, F., Dopazo, J., Ferrer, A. & Conesa, A. Differential expression in RNA-seq: a matter of depth. Genome Res. 21, 2213–2223, 10.1101/gr.124321.111 (2011).
Karimova, G., Pidoux, J., Ullmann, A. & Ladant, D. A bacterial two-hybrid system based on a reconstituted signal transduction pathway. Proc. Natl. Acad. Sci. USA 95, 5752–5756 (1998).
Baker, S. J., Daniels, C. & Morona, R. PhoP/Q regulated genes in Salmonella typhi identification of melittin sensitive mutants. Microb. Pathog. 22, 165–179, 10.1006/mpat.1996.0099 (1997).
Dubrac, S. & Msadek, T. Identification of genes controlled by the essential YycG/YycF two-component system of Staphylococcus aureus. J. Bacteriol. 186, 1175–1181 (2004).
Acknowledgements
This work was supported by the Australian National Health and Medical Research Council (NHMRC) project grant (APP1008973). D.R.C. was supported by an Australian Postgraduate Award and a Monash University Postgraduate Publication Award. A.Y.P. was supported by an Australian NHMRC R.D. Wright Biomedical Fellowship (APP1047916). We thank Ian Monk for the generous gift of the pIMAY plasmid. This work was presented in part at the International Symposium on Staphylococci and Staphylococcal Infections (ISSSI), Chicago, USA, August 2014.
Author information
Authors and Affiliations
Contributions
D.R.C. and A.Y.P. designed the project. D.R.C., J.-H.J., X.K. and D.J.F. performed the experiments. All authors interpreted the data. D.R.C. and A.Y.P. wrote the manuscript. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Cameron, D., Jiang, JH., Kostoulias, X. et al. Vancomycin susceptibility in methicillin-resistant Staphylococcus aureus is mediated by YycHI activation of the WalRK essential two-component regulatory system. Sci Rep 6, 30823 (2016). https://doi.org/10.1038/srep30823
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep30823
This article is cited by
-
Environmental conditions dictate differential evolution of vancomycin resistance in Staphylococcus aureus
Communications Biology (2021)
-
Characterization of methicillin-resistant Staphylococcus aureus through genomics approach
3 Biotech (2020)
-
Staphylococcus aureus biofilm organization modulated by YycFG two-component regulatory pathway
Journal of Orthopaedic Surgery and Research (2019)
-
Zinc-binding to the cytoplasmic PAS domain regulates the essential WalK histidine kinase of Staphylococcus aureus
Nature Communications (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
