
Abstract
Contrast detection thresholds for a central Gabor patch (target) can be modulated by the presence of co-oriented and collinear high contrast Gabors flankers. In foveal vision collinear facilitation can be observed for target-to-flankers relative distances beyond two times the wavelength (λ) of the Gabor’s carrier, while for shorter relative distances (<2λ) there is suppression. These modulatory influences seem to disappear after 12λ. In this study, we measured contrast detection thresholds for different spatial frequencies (1, 4 and 6 cpd) and target-to-flankers relative distances ranging from 6 to 16λ, but with collinear configurations presented in near periphery at 4° of eccentricity. Results showed that in near periphery collinear facilitation extends beyond 12λ for the higher spatial frequencies tested (4 and 6 cpd), while it decays already at 10λ for the lowest spatial frequency used (i.e., 1 cpd). In addition, we found that increasing the spatial frequency the peak of collinear facilitation shifts towards larger target-to-flankers relative distances (expressed as multiples of the stimulus wavelength), an effect never reported neither for near peripheral nor for central vision. The results suggest that the peak and the spatial extent of collinear facilitation in near periphery depend on the spatial frequency of the stimuli used.
Similar content being viewed by others

Introduction
Surrounding elements influence the detection and discrimination of a localized target1,2. There is psychophysical evidence that the contrast detection threshold of a Gabor stimulus can be either increased or reduced by the presence of co-oriented and collinear high contrast Gabor flankers having the same spatial frequency and spatial phase of the target3,4,5. Lateral interactions have been investigated in terms of orientation selectivity4,5, spatial frequency selectivity6 and target-to-flankers relative distances for collinear facilitation4,5; that is, the target-to-flankers relative distance for which the contrast detection threshold of the central target is reduced3. In general, collinear facilitation is observed when flankers are located beyond two times the Gabor carrier’s wavelength (λ) while for shorter distances (<2λ) there is suppression, meaning that the contrast detection thresholds for the central target are higher. It should be noted that in the collinear facilitation literature, distances defined in λ are multiples of the carrier’s wavelength and do not indicate angular distance per se, which is derived by the number of λ and the spatial frequency of the stimulus.
It has been proposed that the anatomical substrates of collinear facilitation are the long-range connections between neurons with similar orientation selectivity in the primary visual cortex (V1)7,8,9,10,11,12,13. This hypothesis is supported by a psychophysical estimation of the propagation speed of collinear facilitation12, that is consistent with measurement from intracellular recording of horizontal connection in V113. The estimated propagation speed of collinear facilitation is ~3 deg/s, that is much slower than feedback and feed forward connections in early visual areas14 and it is consistent with temporal dynamics of lateral conduction of sub-threshold activity elicited by the surround9,13,15. Additionally, physiological studies on integration of responses to a visual target indicate a time window of 200 ms10, in agreement with the integration window for contrast detection16. Consistently, Polat and colleagues17 showed that contrast sensitivity increases from presentation duration of 30 ms to 120 ms, reaching saturation for longer presentation times. Therefore, an input that arrives after a time window of 100–200 ms will no longer be contributing to the target’s response.
Furthermore, there is physiological evidence that horizontal connections are likely to be involved in a variety of contextual modulatory effects: a) in macaque monkeys, inhibition (cooling) of area V2 has no effect on the response to a static texture surround in V1 units18; b) horizontal connections seem to be denser than the feedback projections19; c) both long range striate8,20,21,22,23 and extrastriate7,24 connections are present between units tuned for similar orientation; this is in contrast with Stettler et al.19 who reported that V2–V1 feedback do not occur between units with similar orientation selectivity.
However, collinear facilitation might be also modulated by the activity of higher-level, extrastriate visual areas (e.g., V2, V4). It is well known that the activity of V1 can be modulated by feedback connections controlling the response gain of its target neurons25. There is physiological evidence that inhibition of the extrastriate area V2 produces a decrement of V1 responses14,26. Moreover, neurons in higher-level visual areas present a larger receptive field than V1 neurons, thus pooling information over a larger visual area. Indeed, cell recording in macaque monkeys showed that V2 neurons pool information from an area 5–6 times larger than the integration area of V1 cells27.
On the other hand, other studies do not support the idea that horizontal connections are solely responsible for contextual modulatory effects. For example, in area V1 at 2.5° of eccentricity, an axonal length of 3 mm corresponds to 0.5° in the visual field28, an angular distance much smaller than the extent of modulatory interaction in V129. Consistently, feedback connections, not strictly limited by retinotopy, can gather information from regions that are more far away in the visual field, so they can be a plausible candidate for long-range modulatory effects.
In general, it is plausible that both horizontal and feedback connections contribute to collinear facilitation, with horizontal connections mediating near interactions, such as those located within the summation field, while feedback connections modulate responses to contextual elements located at far distance27,30,31,32.
Overall, these context-related effects seem to be mediated by interactions between cells with a classical receptive field (CRF), organized in hypercolumns and selective for basic features such as orientation, retinal location, spatial phase and spatial frequency.
Similarly to the physiological structure of striate cells, composed by a central excitatory subunit with an antagonistic surround, a proposed corresponding basic unit in visual perception is the Perceptual Field (PF) made up by spatial filters with 2–3 antagonistic elements33,34 with a main component and one or more smaller elements.
Consistent with the role of the CRF in the cytoarchitecture of the visual cortex, the PF represents the first ‘brick’ in the construction of the visual percept. Indeed, previous studies on lateral interactions with Gabor stimuli indicated that collinear facilitation and inhibition mechanisms result from between and within PF stimulation, for facilitatory and suppressory effects, respectively3,35. Therefore, the distance between target and flankers that produces suppression can be used to estimate the size of the PF psychophysically; a question still debated in the case of stimulus presentation in near peripheral vision36. The seminal study of Polat and Sagi3 reported that the foveal range of lateral interactions extends up to 12λ for mid-high spatial frequencies (6–13 cpd), while for flankers placed beyond this distance, the modulation of contrast is marginal or absent4,5,11. Additionally, there is electrophysiological evidence in humans that visually evoked potentials (VEPs) amplitude is maximal for target-to-flankers distances of 1 degree of visual angle for both collinear and orthogonal configurations (i.e., a baseline configuration with flankers orthogonally oriented with respect to the vertical central target)11. Since Polat and Norcia11 used Gabor stimuli with a spatial frequency of 3 cycles per degree (cpd), in their experiment a target-to-flankers distance of 1 deg corresponded to a relative distance of 3λ. This is consistent with psychophysical evidence that the peak of collinear facilitation is located around 3λ3. Polat and Norcia11 also found that response amplitudes of VEPs were significantly lower for the orthogonal configuration, decreasing as a function of distance. Finally, at 4 deg of target-to-flankers angular distance (corresponding to 12λ), no differences in VEPs responses between collinear and orthogonal configuration were observed. As mentioned earlier, this is the same range of collinear facilitation estimated psychophysically in previous studies3.
All the studies on collinear facilitation discussed so far focused on foveal presentation. However physiological evidence from cats and monkeys showed that the long-range connections in the foveal projection of V1 area are a possible neural substrate of this phenomenon that may extend to the peripheral projection area7,21. Moreover, single cell recordings in monkeys and macaques showed contextual modulatory effects for stimuli located up to 10° of eccentricity1,10,13 involving possibly both horizontal connections and/or feedback connections from higher visual areas37. Therefore, physiological evidence seem to indicate that collinear facilitation should be present also for near peripheral presentation of the stimuli. Indeed, a series of recent studies36,38,39 showed that collinear facilitation can be observed in the near-periphery of the visual field (4° of eccentricity), with some differences with respect to the foveal vision: a) it emerges at a target-to-flankers relative distance of 6–8λ36,38, approximately 2–3 times the minimum separation for eliciting collinear facilitation in fovea (3λ); b) it shows a preference for lower spatial frequencies39, consistently with the contrast sensitivity function measured in peripheral vision40; c) it is present from a target-to-flankers relative distance of 6λ for stimuli with a spatial frequency of 1 cpd (corresponding to an angular distance of 8 deg), whereas for higher spatial frequencies collinear facilitation emerged at 8λ (corresponding to an angular distance of 4°, 2° and 1.33° for 2, 4 and 6 cpd, respectively39; d) the difference in contrast sensitivity between collinear and orthogonal condition at the critical relative distance of 8λ decreases as spatial frequency increases39, the opposite of what found in fovea where the magnitude of collinear facilitation (measured at the critical relative distance of 3λ) increases as spatial frequency increases6 (differences are summarized in Table 1). However, apart for these few recent studies, collinear facilitation with stimuli presentation in the near-periphery of the visual field has not been properly investigated. In particular, there are no studies on the extent of the range of collinear interactions (expressed as multiples of the stimulus wavelength) in near periphery. Maniglia and colleagues39 tested up to 8λ, while earlier investigations did not test beyond 7λ36,41.
The aim of the present study was to assess the spatial range of collinear facilitation with stimuli presented at 4° of eccentricity for different spatial frequencies.
The first hypothesis, arising from recent studies on near peripheral collinear facilitation36,38, proposed that the overall range of collinear facilitation would be shifted towards higher target-to-flankers relative distances, with the expected decay of collinear facilitation located beyond the foveal limit of 12λ. This hypothesis is consistent with the idea that PFs increase in size with eccentricity36 and fits with previous studies41,42 in which testing target-to-flankers relative distances that are facilitatory in fovea (3–4λ) led to suppression. This suppression is likely due to the 3-elements collinear configuration still falling within the same PF.
The second hypothesis proposed that the facilitatory range of collinear facilitation (i.e., the absolute length of spatial interaction) would be larger. This because each unit responding to flanking stimuli would have a larger PF and consequently would be activated by stimuli located at farther spatial locations, consistently with the cortical magnification factor43.
The third hypothesis, based on Maniglia et al.39, proposed that different spatial frequencies might show different facilitation curves. In particular, low spatial frequencies might have either an overall larger facilitatory range, starting from 6λ and decaying at the same target-to-flankers distance as for higher spatial frequencies; or it could be shifted leftwards with respect to the higher spatial frequencies, decaying at shorter relative distances.
In order to test these three hypotheses, we measured near peripheral collinear facilitation with one low spatial frequency (1cpd), two intermediate spatial frequencies (4 and 6 cpd) and a large range of target-to-flankers relative distances (i.e., from 4λ to 16λ). In order to obtain a reliable baseline measurement, we measured contrast thresholds also in the orthogonal condition; with flankers orthogonally oriented to the target, a configuration that does not elicit collinear interactions3,11 and is commonly used as baseline condition for measuring collinear facilitation38,39,41.
Concerning the first hypothesis, the facilitation range in near periphery appeared to be overall shorter than the facilitation range reported in fovea. The difference between collinear and orthogonal thresholds was not statistically significant at 12λ for 4 and 6 cpd and at 10λ for 1cpd. However, if we consider the facilitation range in a broader term, as the target-to-flankers relative distance at which the facilitation decays completely (i.e., the distance at which the average contrast thresholds for the collinear condition are equal or higher than those of the baseline condition)3, we observe that the facilitation range for 4 cpd decays completely at 16λ, while for 6 cpd it seems to extend beyond that distance.
For the second hypothesis, when compared to the foveal range of lateral interactions (i.e., from 2λ to 12λ), collinear facilitation in the near periphery does not overcome this range, instead it seems to be shorter, especially for the lowest spatial frequency tested (1cpd).
Consistently, concerning the third hypothesis, it seems that collinear facilitation in the near periphery depends at least in part on the spatial frequency, with an overall extent of collinear interactions larger for the higher spatial frequencies tested (8λ–14λ and 8λ–16λ for 4cpd and 6cpd, respectively) and shorter for the lowest spatial frequency (6λ–12λ for 1 cpd).
In addition, the peak of collinear facilitation seems to be shifted towards larger target-to-flankers relative distances for higher spatial frequencies. This phenomenon has never been reported neither for near peripheral nor for foveal presentations. Previous studies on foveal collinear facilitation assumed that the target-to-flankers relative distance at which the peak of facilitation is reached is independent from spatial frequency3,6. The rationale was that 3λ is the shortest relative distance at which flankers fall outside the PF responding to the target, eliciting in turn modulation between (and not within) PFs.
Experiment 1
The aim of Experiment 1 was to measure the spatial range of collinear facilitation in the near periphery (4° of eccentricity) with low spatial frequency Gabor patches (i.e., 1 cpd). In particular, we assessed the decay of collinear facilitation using a range of target-to-flankers relative distances (i.e., from 6λ to 12λ).
Methods
Apparatus
Stimuli were displayed on a 17” Dell M770 CRT monitor with a refresh rate of 60 Hz. We generated the stimuli with Matlab Psychtoolbox44,45. The screen resolution was 1024 × 768 pixels. Each pixel subtended 1.9 arcmin. The minimum and maximum luminance of the screen were 0.98 cd/m2 and 98.2 cd/m2, respectively and the mean luminance was 47.6 cd/m2. Luminance was measured with a Minolta CS110 (Konica Minolta, Canada). A digital-to-analogue converter (Bits#, Cambridge Research Systems, Cambridge UK) was used to increase the dynamic contrast range (12-bit luminance resolution). A 12-bit gamma-corrected lookup table (LUT) was applied so that luminance was a linear function of the digital representation of the image.
Participants
Two authors (MM and FAJ) and eight naïve observers took part in the experiment. All participants had normal or corrected to normal visual acuity. They sat in a dark room at a distance of 57 cm from the screen. The participant’s head was stabilized using a chinrest. Viewing was binocular. They were instructed to fixate at the center of the screen where a fixation point was always present. All participants took part voluntarily. In addition, all participants gave written informed consent prior to their inclusion in the experiment. This study was conducted in accordance with the Declaration of Helsinki (1964). The experimental protocol was approved by the relevant ethical committee at Centre National de la Recherche Scientifique with our institutional review board (CPP, Comité de Protection des Personnes, protocole 13018–14/04/2014).
Gabor stimuli
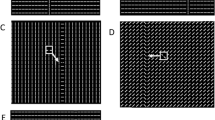
Stimuli were Gabor patches consisting of a cosinusoidal carrier enveloped by a stationary Gaussian and arranged vertically (Fig. 1). The luminance distribution of the Gabor signal was defined as:

Stimuli used in Experiment 1.
(A) Collinear configurations of 1 cpd with target-to-flankers relative distances of 6λ, 8λ, 10λ and 12λ. (B) Orthogonal configurations of 1 cpd with a target-to-flankers relative distance of 6λ, 8λ, 10λ and 12λ. The contrast of the central Gabor patch (i.e., the target) is increased for demonstrative purposes.

where x is the horizontal axis, y the vertical axis, θ is the orientation of the Gabor patch (in radiants), λ is the wavelength of the cosinusoidal carrier and σ is the standard deviation of the Gaussian envelop. In all experiments σ = λ3. Gabor patches had a spatial frequency of 1 cpd. The location of the target relative to the fixation point (0.18 deg) was 4° either in the left or in the right visual hemi-field. A vertical Gabor target was presented flanked above and below by two high-contrast Gabor patches (0.6 Michelson contrast) (Fig. 1). In the collinear configuration target and flankers were vertically oriented (Fig. 1A), whereas in the orthogonal configuration flankers were orthogonally oriented with respect to the target (Fig. 1B). Flankers were located at various distances from the target.
Procedure
Lateral interactions were assessed by comparing the contrast detection thresholds estimated in the collinear and orthogonal configurations as a function of the target-to-flankers relative distance (4λ, 5λ, 6λ, 8λ, 10λ, 12λ and 14λ). In Experiment 1 the spatial frequency tested was 1 cpd. Contrast detection thresholds were measured at 4° of eccentricity. We used a two-interval forced choice task (2IFC) in which participants were required to choose which of the two temporal intervals contained the target. The target was present only in one interval, while the flankers were presented in both intervals. Each interval was presented for 80 ms with an inter-interval delay of 500 ms6,39 (Fig. 2A). The target could be presented either in the left or right visual hemi-field. The temporal interval and the visual hemi-field were randomized on a trial basis (see Fig. 2B).

Schematic representation of the procedure used in Experiment 1.
(A) In the example, the target is shown in the first temporal interval and left visual hemi-filed, whereas only the flankers are displayed in the second temporal interval and in the right visual hemi-field. (B) All the possible combinations of temporal intervals and presentation visual hemi-fields.
The contrast of the target was varied according to a simple 1up-3down staircase46. The starting contrast of the target was set at 0.1 Michelson contrast, increasing of 0.1 log units for each wrong response and decreasing of the same value after three consecutive correct responses. The staircase terminated after either 120 trials or 14 reversals. Contrast thresholds, corresponding to 79% of correct responses, were calculated averaging the contrast values corresponding to the last 6 reversals, regardless the temporal interval in which the target was presented and the presentation visual hemi-field. An acoustic feedback (50 ms tone of 500 Hz) was provided with wrong answers. Observers performed 8 blocks in which the target-to-flankers relative distance and the flankers’ orientation were varied. Observers performed the experiment in one day.
Results
Figure 3 shows the results of Experiment 1. Contrast thresholds (Michelson contrast) and differences between collinear and orthogonal contrast thresholds are shown as a function of the target-to-flankers relative distance.

Results of Experiment 1.
(A) Mean contrast thresholds (Michelson contrast) for collinear and orthogonal configurations as a function of target-to-flankers relative distance. (B) Mean differences between collinear and orthogonal contrast thresholds (i.e., collinear-orthogonal difference) as a function of the target-to-flankers relative distance. The horizontal black dashed line represents a difference equal to zero (i.e., no modulation). Data points below zero represent collinear facilitation. Error bars ± s.e.m.
Spatial extent of collinear facilitation
To estimate the spatial range of collinear facilitation for 1 cpd stimuli, we conducted a repeated-measures ANOVA including as factors the configuration (collinear vs. orthogonal) and the target-to-flankers relative distance (4λ, 5λ, 6λ, 8λ, 10λ, 12λ, 14λ). The ANOVA did not report a significant effect of the target-to-flankers relative distance (F2.34, 21.09 = 1.47, p = 0.251, partial-η2 = 0.14, degrees of freedom and p value were corrected with the Greenhouse-Geisser correction because of sphericity violation), but reported a marginally significant effect of the configuration (F1,9 = 4.62, p = 0.06, partial-η2 = 0.34). The interaction between configuration and target-to-flankers relative distance was significant (F6,54 = 6.65, p = 0.0001, partial-η2 = 0.43). Post-hoc t-tests, using a false discovery rate (FDR) of 0.0547, showed that at 4λ the contrast thresholds in the collinear condition was higher than the orthogonal contrast threshold (Table 2, line 1); this is consistent with the idea that for short relative distances, collinear interactions in the near periphery are mostly inhibitory38,41. On the other hand, collinear thresholds were lower for target-to-flankers relative distance of 6λ and 8λ (see Table 2 and Fig. 3A). These results are consistent with previous studies36,38,39.
Figure 3B shows the mean differences between the contrast thresholds estimated in the collinear configuration and those estimated in the orthogonal configuration for each target-to-flankers relative distance. For the sake of simplicity we will refer to the difference between collinear and orthogonal threshold as “collinear-orthogonal difference”. A repeated-measures ANOVA on the mean collinear-orthogonal differences and including as factor the target-to-flankers relative distance reported a significant effect of the separation (F6,54 = 6.65, p = 0.0001, partial-η2 = 0.43). Simple within-subjects contrasts showed that the collinear-orthogonal difference at 4λ was significantly different than the collinear-orthogonal differences calculated for all the other target-to-flankers relative distances (Table 3). In addition, reverse Helmert within-subjects contrasts showed a significant difference between the collinear-orthogonal difference at 6λ and 5λ (F1,9 = 13.82, p = 0.005, partial-η2 = 0.61) and a significant difference between the collinear-orthogonal difference at 8λ and 6λ (F1,9 = 7.71, p = 0.022, partial-η2 = 0.46).
Finally, we performed seven two-sided one-sample t-tests using a FDR of 0.05. The t-tests were performed between collinear-orthogonal differences, for each target-to-flankers relative distance and zero (i.e., no modulation). The t-tests showed that collinear-orthogonal differences were significantly lower than zero for 6λ and 8λ and significantly higher than zero for 4λ (Table 4).
Experiment 2
The aim of Experiment 2 was to measure collinear facilitation in the near periphery (4° of eccentricity) with mid-high spatial frequencies (i.e., 4 and 6 cpd). In particular, this experiment was conducted to assess whether higher spatial frequencies show a different range of collinear facilitation. In Experiment 1 and in our previous study39 we found that for 1 cpd, collinear facilitation emerged at shorter target-to-flankers relative distance than for higher spatial frequencies (i.e., 6λ for 1cpd, ~8λ for 2 and 3 cpd39) and vanishes around 10λ (against the 12λ of the fovea3). Such reduced range of collinear facilitation might be due to the temporal integration window for the central target10, so that inputs coming from the flankers fail to reach the target within the temporal constraint of contrast integration6,12. Since in the lateral interactions paradigm3 lower spatial frequencies would lead to greater angular distances between target and flankers, it is possible that the reduced range of collinear facilitation depends on the failure of temporal signal integration between flankers and target. In order to test whether higher spatial frequencies would lead to a larger collinear facilitation range, we tested collinear facilitation at 4 and 6 cpd for a number of target-to-flankers relative distances.
Methods
One author (MM) and a new sample of nine naïve observers took part in Experiment 2. Stimuli and apparatus were the same as in Experiment 1. The procedure was the same as used in Experiment 1 except that we used spatial frequencies of 4 and 6 cpd and target-to-flankers relative distances ranging from 6λ to 16λ (i.e., 6λ, 8λ, 10λ, 12λ, 14λ and 16λ).
Results
Extent of collinear facilitation for higher spatial frequency
Figure 4 shows contrast thresholds (panel A) and mean collinear-orthogonal differences (panel B) as a function of the target-to-flankers relative distance. A repeated-measures ANOVA on the contrast thresholds and including as factors the spatial frequency (4 cpd vs. 6 cpd), the configuration (collinear vs. orthogonal) and the target-to-flankers relative distance (6λ, 8λ, 10λ, 12λ, 14λ, 16λ), reported a significant effect of the spatial frequency (F1,9 = 26.66, p = 0.001, partial-η2 = 0.75) and a significant effect of the configuration (F1,9 = 9.62, p = 0.013, partial-η2 = 0.52). In addition to better investigate the pattern of lateral interaction for mid-high spatial frequencies we conducted two repeated-measures ANOVA separately for 4 and 6 cpd.

(A,B) Mean contrast detection thresholds (Michelson contrast) for collinear and orthogonal configurations as a function of the target-to-flankers relative distance for 4 cpd (panel A) and 6 cpd (panel B). (C) Mean collinear-orthogonal difference for each target-to-flankers relative distance and for 4 and 6 cpd. The horizontal black dashed line represents a difference equal to zero (i.e., no modulation). Data points below zero represent collinear facilitation. Error bars ± s.e.m.
For 4 cpd the ANOVA reported a significant effect of the configuration (F1,9 = 8.43, p = 0.017, partial-η2 = 0.48), but not a significant effect of the target-to-flankers relative distance (F1.17, 10.6 = 0.47, p = 0.54, partial-η2 = 0.05) or a significant interaction configuration x target-to-flankers relative distance (F1.6, 14.5 = 1.35, p = 0.28, partial-η2 = 0.13).
For 6 cpd, the ANOVA also reported only a significant effect of the configuration (F1,9 = 6.11, p = 0.035, partial-η2 = 0.40), but not a significant effect of the target-to-flankers relative distance (F2.35, 21.2 = 0.793, p = 0.48, partial-η2 = 0.08) or a significant interaction configuration x target-to-flankers relative distance (F2.21, 19.94 = 0.66, p = 0.54, partial-η2 = 0.07).
Despite the interaction was not significant, we also conducted a series of paired t-tests between the contrast thresholds estimated in the collinear and orthogonal configurations, separately for each target-to-flankers relative distance38,39 and spatial frequency. Significant paired t-tests are summarized in Table 5. Overall, the results showed that for both spatial frequencies (i.e., 4 and 6 cpd) the contrast thresholds in the collinear condition were lower for target-to-flankers relative distance of 8λ and 10λ. In addition, the difference between contrast thresholds estimated in the collinear and orthogonal configurations at 4 cpd for 12λ was close to significance (t(9) = −2.20, p = 0.055, d = 0.69).
Additionally, we conducted a repeated-measures ANOVA, separately for 4 and 6 cpd, on the collinear-orthogonal differences including as factor the target-to-flankers relative distance. The ANOVA did not report a significant effect of the target-to-flankers relative distance for both spatial frequencies (4 cpd: F1.6, 14.5 = 1.35, p = 0.28, partial-η2 = 0.13; 6cpd: F2.16,19.5 = 0.66, p = 0.54, partial-η2 = 0.068). Finally, we also compared the collinear-orthogonal differences with respect to zero. The results showed that for both spatial frequencies (i.e., 4 and 6 cpd) collinear-orthogonal differences were significantly lower than zero at 8λ and 10λ (Table 6). In addition, for 4 cpd the difference between threshold elevation and zero at 12λ was close to significance (t(9) = −2.20, p = 0.055, d = 0.69).
Peak of collinear facilitation
There is psychophysical evidence that foveal collinear facilitation peaks at target-to-flankers relative distance of 3λ3,4,5,11 and this regardless the spatial frequency used6. In an additional analysis we fitted a lognormal function on the mean collinear-orthogonal differences in order to estimate the peak of collinear facilitation (i.e., the lowest point of the curve) in near periphery. The non-linear fit was conducted using SciPy for Python48,49. The lognormal function used was:

where μ represents the value at which the curve reaches its minimum, σ corresponds to the shape of the function and α is the scale of the function. We estimated the minimum values for the function that best fitted the obtained data. At 1 cpd the minimum values obtained was −0.0046 ± 0.013 s.d. and it was found at 7.52 ± 0.29 s.d. Thus, the peak of facilitation corresponds to a target-to-flankers relative distance of 7.52λ. This is consistent with our previous study in the 1 cpd condition39.
Equation (2) was also fitted to collinear-orthogonal differences of Experiment 2 in order to estimate the peak of collinear facilitation for spatial frequencies of 4 and 6 cpd. The estimated minimum values were −0.0297 ± 0.009 s.d. at 8.75 ± 0.54 s.d. for 4 cpd and −0.0302 ± 0.011 s.d. at 9.62 ± 0.77 s.d. for 6 cpd. Thus, the peak of facilitation for 4 and 6 cpd was located at a target-to-flankers relative distance of 8.75λ and 9.62λ, respectively. In these two cases, in order to establish more precisely the location of the peak of collinear facilitation, we excluded from the fitting procedure the mean collinear-orthogonal difference in correspondence of 16λ. This is because at 16λ there seems to be some residual collinear facilitation, but contrast thresholds (and consequently collinear-orthogonal differences) obtained at this target-to-flankers relative distance are quite noisy and the collinear facilitation estimated is not statistically significant. To establish the goodness of fit of our data on the obtained lognormal curves, we performed a post-hoc goodness of fit. Our values were quite close to those expected by those predicted by the function (1 cpd: χ2 = −0.002, p > 0.05; 4 cpd: χ2 = −0.0026, p > 0.05; 6 cpd: χ2 = − 0.012, p > 0.05). This corroborates our idea that the lognormal distribution explain quite well the behaviour of the results at different spatial frequencies. Figure 5 shows collinear-orthogonal differences for each spatial frequency and relative fits.

Collinear-orthogonal differences as a function of the target-to-flankers relative distance for 1 (light grey dots), 4 (grey dots) and 6 cpd (black dots) configurations.
Lognormal fits are also reported as continuous lines in the corresponding colours whose standard deviations correspond to the surrounding semi-transparent area. Vertical dashed lines correspond to the minimum values of the fitted function for each spatial frequency.
Discussion
In this study we tested the extent of collinear facilitation for configurations presented at 4° of eccentricity (i.e., near periphery). The rationale was that since previous studies showed that collinear facilitation in the near periphery of the visual field emerges at target-to-flankers relative distances 2–3 times larger than in the fovea36,38,39, the spatial extent of facilitation in the near periphery might be different with respect to the fovea.
In Experiment 1 we used low spatial frequency stimuli (1 cpd) and the results showed collinear facilitation at a target-to-flankers relative distance of 6λ36,39, decaying at 10λ. Overall, this range is shorter than the typical range of collinear facilitation found in fovea, extending from 1.5–2λ to 12λ3,11.
In Experiment 2 we used stimuli with higher spatial frequencies (i.e., 4 and 6 cpd). The rationale was that the magnitude of collinear contrast sensitivity in the near periphery seems to differ depending on the spatial frequency used, being weaker for high spatial frequencies36,38,39. Results showed that collinear facilitation emerges at a target-to-flankers relative distance of 8λ for both spatial frequencies, returning to baseline at 12λ (i.e., collinear contrast thresholds not significantly lower than orthogonal contrast thresholds).
Overall, collinear facilitation, defined as the target-to-flankers relative distance at which collinear contrast thresholds are significantly lower than orthogonal contrast thresholds, seems to differ between low spatial frequency (1cpd) and mid-high spatial frequencies (4 and 6cpd) both in terms of spatial range and magnitude of the effect. In particular, based on the curve fitting with the lognormal function, for the lowest spatial frequency facilitation peaked at 7.52λ, while for 4 and 6 cpd, it peaked at 8.75λ and 9.62λ, respectively. Moreover, the facilitatory effect of collinear flankers (expressed as the difference between collinear and orthogonal contrast thresholds) for 1 cpd is one order of magnitude smaller than 4 and 6 cpd, while the suppressory effect at shorter target-to-flankers relative distances is much stronger for 1 cpd than for 4 and 6 cpd.
However, it should be noted that when Polat and Sagi3 psychophysically described the phenomenon of lateral interactions and collinear facilitation, they did not perform statistical analysis to support their findings, rather the authors indicated 12λ as the target-to-flankers relative distance at which contrast thresholds return to baseline. Thus, most of these effects have been previously described without a statistical cut-off to define formally the range of foveal facilitation and this makes difficult to compare our results with previous findings in foveal vision. Accordingly, if we adopt a more descriptive approach, considering the extent of collinear facilitation as the maximum target-to-flankers relative distance at which the difference between collinear and orthogonal contrast thresholds approaches zero, we can observe how collinear thresholds for 4 cpd are on average lower than the orthogonal thresholds for all the target-to-flankers relative distances tested, except for the last one, returning above zero at 16λ. On the other hand, for 6 cpd, collinear thresholds are lower than orthogonal thresholds across the entire range of relative distances tested.
Within this framework, the estimation of 3 deg of spatial extent for collinear facilitation11 is too short to explain the effect we found at 8λ with 1cpd stimuli, for which the angular distance between the target and flankers is 8 deg. Similarly, the foveal limit of 12λ is overcome for the highest spatial frequencies tested (i.e., 6 cpd). Interestingly, the peak of collinear facilitation seems to shift rightwards, i.e., towards larger target-to-flankers relative distances, but shorter angular distances. This is because σ = λ, so higher spatial frequencies are characterized by shorter target-to-flankers distances in terms of angular distance. This phenomenon has never been reported in previous studies with foveal or peripheral stimulus presentation. A shorter range of collinear facilitation is expected for lower spatial frequencies because of the neural integration time; that is, neural signals from flankers located at more than 10 deg of visual angle (i.e., 10λ with 1 cpd stimuli) would fail to reach the neuron responding to the target within the contrast integration time window. Polat6 showed that the magnitude of collinear facilitation in fovea increases with increasing spatial frequency. The author argued that facilitation for low spatial frequencies is reduced because of the slow propagation speed of the flankers input, failing in combining with the target’s input within the temporal integration window, estimated to be around 200 ms10,50. Moreover, since collinear facilitation seems involved in contour integration33,51, the preference for high spatial frequencies can be explained in terms of ecological values, since intermediate and high spatial frequencies are more involved in this process and more trained by everyday life6,52. However, Maniglia et al.39 reported how collinear facilitation with stimuli presented in the near periphery of the visual field (4° of eccentricity) seems to show an inverse pattern, with a preference for lower spatial frequencies, consistent with the known spatial frequency tuning of this portion of the visual field53. Therefore, it seems also that the temporal integration window and/or the propagation speed are different between the fovea and the near periphery of the visual field. Polat6 reported how estimation of cortico-cortical propagation, provided by psychophysics studies12,54 is about 3 deg/s. However, while these estimations both in terms of temporal integration window and propagation speed may be plausible and consistently verified in foveal vision, they are not suitable to explain the effect observed in the present study, in which we reported statistically significant facilitation at 1 cpd for 8λ and up to 10λ with 4 cpd (Fig. 6).

Spatial range of near peripheral collinear facilitation expressed in angular distance (degrees of visual angle) for 1 and 4 cpd.
Only the target-to-flankers relative distances for which we showed a significant difference between collinear and orthogonal conditions are reported. Collinear facilitation is scaled for the wavelength of the stimuli used (emerging at 6λ and 8λ for the spatial frequencies shown) independently from the angular distance.
The angular distance between target and flankers in the classical lateral interaction paradigm is related to the spatial frequency of the Gabor stimuli used (since σ = λ), so in the case of spatial frequency of 1 cpd, a target-to-flankers relative distance of 8λ is four times larger, in terms of visual angle, than a target-to-flankers relative distance of 8λ with 4 cpd stimuli (Fig. 6).
If the integration time is 200 ms and the propagation speed from flankers’ location is 3 deg/s, then for 1 cpd and target-to-flankers relative distance of 8λ, the input from the flankers would arrive at the target location after 2.66 seconds (being the angular distance between flankers and target 8 deg), while for 4 cpd and target-to-flankers relative distance of 10λ (target-to-flankers angular distance of 2.5 deg), it would be 830 ms, that is beyond the integration time window10.
However, more systematic studies taking into account the cortical magnification factor, the propagation speed for near peripheral stimulation and the integration time for near peripheral contrast response are needed in order to shed light on this phenomenon.
As reported in the introduction section, one of the proposed anatomical substrate for collinear facilitation are the long-range horizontal connections between units in the primary visual cortex sharing the same orientation selectivity7,21,55,56,57. Several studies have proven that neurons can synchronize their firing rate with a millisecond precision58. This effect was reported for spatially separated units and involves stimuli similar to those used to investigate collinear facilitation. Consequently, it is possible that practice can improve collinear facilitation in the near periphery and uncover a larger range of facilitation also for low spatial frequencies, since previous studies with foveal presentation already showed that collinear facilitation can be strengthened through practice4,5,59,60. Interestingly, Polat6 also proposed that the propagation time may be slower in the periphery. However, this is somewhat inconsistent with the data presented here, showing that facilitation may arise for target-to-flankers angular distances of 8 deg.
An alternative explanation is that propagation speed is the same for horizontal long-range connection between units coding central and near peripheral vision. In this case, the size of PFs is larger, so that each unit analyses a bigger portion of the visual field and anatomically close neurons in the visual cortex are responsible of a wider portion of the visual field. Therefore, anatomically close neurons can be activated by stimuli located in more distant spatial locations. This is consistent with the definition of the cortical magnification factor43. Overall, our data show that when presenting near peripheral stimuli, propagation time is fast enough to lead to the integration of signals coming from the flankers for distances of 8λ with 1 cpd stimuli (i.e., for an angular distance between flankers and target of 8 deg).
However, a strong alternative explanation for the effects we reported would take into account feedback mechanisms from higher-level visual areas. One of the main reasons is the angular distances at which contextual modulations are still present (8 deg). Such large interactions cannot be easily explained in terms of horizontal connections in early visual cortex alone, suggesting the involvement of extrastriate areas, whose units present receptive fields up to 6 times bigger than V127. Moreover, as reported in the introduction section, the retinal extent of contextual modulation in V1 is wider than the area encompassed by the average axonal length alone28.
Future psychophysical studies might reveal that this range of collinear facilitation can be extended by practice, promoting synaptic synchronization for larger target-to-flankers relative distances, as already showed for foveal presentation4.
A further question that should be addressed is whether the minimum target-to-flankers relative distance necessary to elicit collinear facilitation increases with increasing eccentricity. To our knowledge, collinear facilitation in the periphery of the visual field has been tested mainly at 4° of eccentricity36,38,41,42,61 and overall not beyond 6° 42. Assuming that the size of PFs increases as a function of eccentricity, we might expect that facilitation emerges at larger target-to-flankers relative distances for more eccentric spatial locations. However, in the case of large angular distances, the propagation of the signal from the flankers to the target might not be fast enough to reach the target location within the integration time window, thus interfering with collinear facilitation. Further studies might address this issue by training subjects on larger target-to-flankers relative distances.
Additional Information
How to cite this article: Maniglia, M. et al. The spatial range of peripheral collinear facilitation. Sci. Rep. 5, 15530; doi: 10.1038/srep15530 (2015).
References
Kapadia, M. K., Ito, M., Gilbert, C. D. & Westheimer, G. Improvement in visual sensitivity by changes in local context: parallel studies in human observers and in V1 of alert monkeys. Neuron 15, 843–856 (1995).
Bonneh, Y. & Sagi, D. Effects of spatial configuration on contrast detection. Vision research 38, 3541–3553 (1998).
Polat, U. & Sagi, D. Lateral interactions between spatial channels: suppression and facilitation revealed by lateral masking experiments. Vision research 33, 993–999 (1993).
Polat, U. & Sagi, D. Spatial interactions in human vision: from near to far via experience-dependent cascades of connections. Proceedings of the National Academy of Sciences of the United States of America 91, 1206–1209 (1994).
Polat, U. & Sagi, D. The architecture of perceptual spatial interactions. Vision research 34, 73–78 (1994).
Polat, U. Effect of spatial frequency on collinear facilitation. Spatial vision 22, 179–193, doi: 10.1163/156856809787465609 (2009).
Gilbert, C. D. & Wiesel, T. N. Columnar specificity of intrinsic horizontal and corticocortical connections in cat visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience 9, 2432–2442 (1989).
Ts’o, D. Y., Gilbert, C. D. & Wiesel, T. N. Relationships between horizontal interactions and functional architecture in cat striate cortex as revealed by cross-correlation analysis. The Journal of neuroscience: the official journal of the Society for Neuroscience 6, 1160–1170 (1986).
Grinvald, A., Lieke, E. E., Frostig, R. D. & Hildesheim, R. Cortical point-spread function and long-range lateral interactions revealed by real-time optical imaging of macaque monkey primary visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience 14, 2545–2568 (1994).
Polat, U., Mizobe, K., Pettet, M. W., Kasamatsu, T. & Norcia, A. M. Collinear stimuli regulate visual responses depending on cell’s contrast threshold. Nature 391, 580-584, doi: 10.1038/35372 (1998).
Polat, U. & Norcia, A. M. Neurophysiological evidence for contrast dependent long-range facilitation and suppression in the human visual cortex. Vision research 36, 2099–2109 (1996).
Cass, J. R. & Spehar, B. Dynamics of collinear contrast facilitation are consistent with long-range horizontal striate transmission. Vision research 45, 2728–2739, doi: 10.1016/j.visres.2005.03.010 (2005).
Bringuier, V., Chavane, F., Glaeser, L. & Fregnac, Y. Horizontal propagation of visual activity in the synaptic integration field of area 17 neurons. Science 283, 695–699 (1999).
Hupe, J. M. et al. Feedback connections act on the early part of the responses in monkey visual cortex. Journal of neurophysiology 85, 134–145 (2001).
Rossi, A. F., Desimone, R. & Ungerleider, L. G. Contextual modulation in primary visual cortex of macaques. The Journal of neuroscience: the official journal of the Society for Neuroscience 21, 1698–1709 (2001).
Watson, A. B., Barlow, H. B. & Robson, J. G. What does the eye see best? Nature 302, 419–422 (1983).
Polat, U., Sterkin, A. & Yehezkel, O. Spatio-temporal low-level neural networks account for visual masking. Advances in cognitive psychology / University of Finance and Management in Warsaw 3, 153–165, doi: 10.2478/v10053-008-0021-4 (2007).
Hupe, J. M., James, A. C., Girard, P. & Bullier, J. Response modulations by static texture surround in area V1 of the macaque monkey do not depend on feedback connections from V2. Journal of neurophysiology 85, 146–163 (2001).
Stettler, D. D., Das, A., Bennett, J. & Gilbert, C. D. Lateral connectivity and contextual interactions in macaque primary visual cortex. Neuron 36, 739–750 (2002).
Hirsch, J. A. & Gilbert, C. D. Synaptic physiology of horizontal connections in the cat’s visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience 11, 1800–1809 (1991).
Malach, R., Amir, Y., Harel, M. & Grinvald, A. Relationship between intrinsic connections and functional architecture revealed by optical imaging and in vivo targeted biocytin injections in primate striate cortex. Proceedings of the National Academy of Sciences of the United States of America 90, 10469–10473 (1993).
Schwarz, C. & Bolz, J. Functional specificity of a long-range horizontal connection in cat visual cortex: a cross-correlation study. The Journal of neuroscience: the official journal of the Society for Neuroscience 11, 2995–3007 (1991).
Weliky, M., Kandler, K., Fitzpatrick, D. & Katz, L. C. Patterns of excitation and inhibition evoked by horizontal connections in visual cortex share a common relationship to orientation columns. Neuron 15, 541–552 (1995).
Shmuel, A., Korman, M., Harel, M., Grinvald A. & Malach R. Relationship of feedback connections from area V2 to orientation domains in area V1 of the primate. Society Neuroscience Abstract 24, 767 (1998).
Shao, Z. & Burkhalter, A. Different balance of excitation and inhibition in forward and feedback circuits of rat visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience 16, 7353–7365 (1996).
Mignard, M. & Malpeli, J. G. Paths of information flow through visual cortex. Science 251, 1249–1251 (1991).
Angelucci, A. et al. Circuits for local and global signal integration in primary visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience 22, 8633–8646 (2002).
Dow, B. M., Snyder, A. Z., Vautin, R. G. & Bauer, R. Magnification factor and receptive field size in foveal striate cortex of the monkey. Experimental brain research 44, 213–228 (1981).
Levitt, J. B. & Lund, J. S. Contrast dependence of contextual effects in primate visual cortex. Nature 387, 73–76, doi: 10.1038/387073a0 (1997).
Brown, H. A., Allison, J. D., Samonds, J. M. & Bonds, A. B. Nonlocal origin of response suppression from stimulation outside the classic receptive field in area 17 of the cat. Visual neuroscience 20, 85–96 (2003).
Cavanaugh, J. R., Bair, W. & Movshon, J. A. Nature and interaction of signals from the receptive field center and surround in macaque V1 neurons. Journal of neurophysiology 88, 2530–2546, doi: 10.1152/jn.00692.2001 (2002).
Chisum, H. J., Mooser, F. & Fitzpatrick, D. Emergent properties of layer 2/3 neurons reflect the collinear arrangement of horizontal connections in tree shrew visual cortex. The Journal of neuroscience: the official journal of the Society for Neuroscience 23, 2947–2960 (2003).
Polat, U. & Tyler, C. W. What pattern the eye sees best. Vision research 39, 887–895 (1999).
Watson, A. B. Transfer of contrast sensitivity in linear visual networks. Visual neuroscience 8, 65–76 (1992).
Zenger, B. & Sagi, D. Isolating excitatory and inhibitory nonlinear spatial interactions involved in contrast detection. Vision research 36, 2497–2513 (1996).
Lev, M. & Polat, U. Collinear facilitation and suppression at the periphery. Vision research 51, 2488–2498, doi: 10.1016/j.visres.2011.10.008 (2011).
Angelucci, A. & Bullier, J. Reaching beyond the classical receptive field of V1 neurons: horizontal or feedback axons? J Physiol Paris 97, 141–154 (2003).
Maniglia, M. et al. Reducing crowding by weakening inhibitory lateral interactions in the periphery with perceptual learning. PloS one 6, e25568, doi: 10.1371/journal.pone.0025568 (2011).
Maniglia, M., Pavan, A. & Trotter, Y. The effect of spatial frequency on peripheral collinear facilitation. Vision research 107, 146–154, doi: 10.1016/j.visres.2014.12.008 (2015).
Johnston, A. Spatial scaling of central and peripheral contrast-sensitivity functions. Journal of the Optical Society of America. A, Optics and image science 4, 1583–1593 (1987).
Shani, R. & Sagi, D. Eccentricity effects on lateral interactions. Vision research 45, 2009–2024, doi: 10.1016/j.visres.2005.01.024 (2005).
Williams, C. B. & Hess, R. F. Relationship between facilitation at threshold and suprathreshold contour integration. Journal of the Optical Society of America. A, Optics, image science and vision 15, 2046–2051 (1998).
Horton, J. C. & Hoyt, W. F. The representation of the visual field in human striate cortex. A revision of the classic Holmes map. Archives of ophthalmology 109, 816–824 (1991).
Brainard, D. H. The Psychophysics Toolbox. Spatial vision 10, 433–436 (1997).
Pelli, D. G. The VideoToolbox software for visual psychophysics: transforming numbers into movies. Spatial vision 10, 437–442 (1997).
Levitt, H. Transformed up-down methods in psychoacoustics. The Journal of the Acoustical Society of America 49, Suppl 2:467+ (1971).
Benjamini, Y. H. & Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the royal statistical society. Series B (Methodological) 57, 289–300 (1995).
Millman, K. J. A. & M. Python for Scientists and Engineers. Computing in Science & Engineering 13, 9–12 (2011).
Oliphant, T. E. Python for Scientific Computing. Computing in Science & Engineering 9, 10–20 (2007).
Albrecht, D. G. Visual cortex neurons in monkey and cat: effect of contrast on the spatial and temporal phase transfer functions. Visual neuroscience 12, 1191–1210 (1995).
Kovacs, I. Gestalten of today: early processing of visual contours and surfaces. Behav Brain Res 82, 1–11. (1996).
Dakin, S. C. & Hess, R. F. Spatial-frequency tuning of visual contour integration. Journal of the Optical Society of America. A, Optics, image science and vision 15, 1486–1499 (1998).
Rovamo, J. & Virsu, V. An estimation and application of the human cortical magnification factor. Experimental brain research 37, 495–510 (1979).
Polat, U. & Sagi, D. Temporal asymmetry of collinear lateral interactions. Vision research 46, 953–960, doi: 10.1016/j.visres.2005.09.031 (2006).
Gilbert, C. D. & Wiesel, T. N. Intrinsic connectivity and receptive field properties in visual cortex. Vision research 25, 365–374 (1985).
Gilbert, C. D. & Wiesel, T. N. The influence of contextual stimuli on the orientation selectivity of cells in primary visual cortex of the cat. Vision research 30, 1689–1701 (1990).
Weliky, M. & Katz, L. C. Functional mapping of horizontal connections in developing ferret visual cortex: experiments and modeling. The Journal of neuroscience: the official journal of the Society for Neuroscience 14, 7291–7305 (1994).
Lowel, S. & Singer, W. Selection of intrinsic horizontal connections in the visual cortex by correlated neuronal activity. Science 255, 209–212 (1992).
Polat, U., Ma-Naim, T., Belkin, M. & Sagi, D. Improving vision in adult amblyopia by perceptual learning. Proceedings of the National Academy of Sciences of the United States of America 101, 6692–6697, doi: 10.1073/pnas.0401200101 (2004).
Polat, U. & S. D. In Maturational Windows and Adult Cortical Plasticity Vol. XXIV (ed Kovács I. & Julesz B. (eds.)) 111–125 (Addison-Wesley, 1995).
Giorgi, R. G., Soong, G. P., Woods, R. L. & Peli, E. Facilitation of contrast detection in near-peripheral vision. Vision Res 44, 3193–3202 (2004).
Acknowledgements
Author M.M. was supported by the Fouassier Foundation (France) and the CerCo, Toulouse (France). Authors would like to thank Samy Rima for English proofreading.
Author information
Authors and Affiliations
Contributions
M.M. and A.P. designed and implemented the experiments. M.M. collected the data. M.M., F.A.J. and A.P. analysed data, interpreted the results and wrote the main manuscript. Y.T. reviewed the manuscript text. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Maniglia, M., Pavan, A., Aedo-Jury, F. et al. The spatial range of peripheral collinear facilitation. Sci Rep 5, 15530 (2015). https://doi.org/10.1038/srep15530
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep15530
This article is cited by
-
Correlations between the metabolic profile and 18F-FDG-Positron Emission Tomography-Computed Tomography parameters reveal the complexity of the metabolic reprogramming within lung cancer patients
Scientific Reports (2019)
-
TMS reveals inhibitory extrastriate cortico-cortical feedback modulation of V1 activity in humans
Brain Structure and Function (2019)
-
Spontaneous and training-induced cortical plasticity in MD patients: Hints from lateral masking
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
