
Abstract
We present a micro-device in which more than 10,000 asymmetric lipid bilayer membranes are formed at a time on micro-chamber arrays. The arrayed asymmetric lipid bilayers, where lipid compositions are different between the inner and outer leaflets, are formed with high efficiency of over 97% by injecting several types of liquids into a micro-device that has hydrophilic-in-hydrophobic surfaces. The lipid compositional asymmetry is an intrinsic property of bio-membranes and therefore, this micro-device extends the versatility of artificial lipid-bilayer systems, which were previously limited to symmetric bilayer formation and could contribute to the understanding of the role of lipid compositional asymmetry in cell physiology and also to further analytical and pharmacological applications.
Similar content being viewed by others

Introduction
Most biological membranes possess an asymmetric transbilayer phospholipid distribution, in which endogenous enzymes expend energy to maintain the arrangement by promoting phospholipid translocation or flip-flop1. In particular, the plasma membrane of most eukaryotes maintains a high degree of asymmetry, which controls various cellular functions, such as signal transduction, membrane fusion and cell apoptosis2,3,4. Therefore, extensive studies have been conducted to further understand the role of asymmetric distribution in cell physiology by developing in vitro platforms to form lipid bilayer membranes with an asymmetric lipid composition2,5,6,7.
In general, an asymmetric lipid bilayer is formed by assembling two independently prepared lipid monolayers5,6 or altering the distribution of specific phospholipids using chemical or mechanical perturbation, e.g., pH gradient and osmotic pressure2, or using molecules that promote lipid redistribution, e.g., translocase2. Although these techniques are powerful for qualitative analysis of the effect of asymmetric lipid bilayers on cellular functions, it remains difficult to perform quantitative analyses because it is technically challenging to prepare asymmetric lipid bilayers with high reproducibility in a high-throughput manner given the complexity of the bilayer formation process and the size heterogeneity of the formed bilayer membranes. Thus, many aspects remain unclear with respect to the physiological role of lipid compositional asymmetry.
Recently, some techniques using micro-fabricated devices, which enable the controlled handling of less than microliter volumes of fluids, have been developed to create artificial lipid bilayers with a simple procedure8,9. Although these devices can form lipid bilayers with a uniform size in a high-throughput manner, it remains technically difficult to control the transbilayer distribution of phospholipids because they are based on the spontaneous assembly of lipid bilayers. In this study, we address this issue by developing a method to independently prepare two lipid monolayers in a simple manner and assemble them to form asymmetric lipid bilayers with uniform size in a high-throughput manner.
Results
Fabrication of micro device
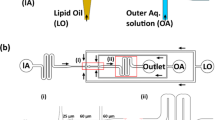
Using conventional soft lithography, we fabricated a micro-device with more than 10,000 through-hole structures (7 fL, Φ = 4 μm and h = 0.5 μm) on a carbon-fluorine hydrophobic polymer (CYTOP, Asahi-glass, Japan) on a hydrophilic glass substrate9,10,11,12 (Fig. 1). Because the fabrication process was well established in our previous studies10,11,12, we are able to efficiently fabricate the micro-device, achieving a fabrication success rate of around 100%8. The through-hole structures were utilized as micro-chambers, whose orifice is suitable to mechanically support the lipid membrane because of its hydrophobicity8. Teflon has high hydrophobicity and has thus frequently been used as a support medium for lipid membranes in previous studies8. For exchange of the sample solution, a flow channel was constructed from the micro-chamber array patterned on a glass substrate, a spacer sheet and a glass block, which had an access port for sample injection (Fig. 1a).

Micro-chamber arrays for asymmetric lipid bilayer formation.
(a) Fabricated micro-device where the substrate of hydrophilic-in-hydrophobic structures is assembled with cover glass that has an access port for sample injection. (b) Bright field image of the through-hole structures of the carbon-fluorine hydrophobic polymer on a hydrophilic glass substrate. (c) Illustration of a fabricated structure (diameter, 4 μm; thickness, 500 nm).
High throughput formation of asymmetric lipid bilayers
Planar lipid membranes were formed on the top orifice of the micro-chambers via sequential injection of liquids from the access port (Fig. 2a). This technique is similar to the conventional technique known as the “painting technique”, which forms lipid bilayers by deposition of lipid solution across a small aperture in a hydrophobic material12,13. The experimental procedures developed in this study to form asymmetric planar lipid bilayers were composed of four steps. First, an aqueous solution was infused into the flow cell. After the infusion, micro-chambers were filled with the aqueous solution. Second, a lipid solution containing 20 μg mL−1 lipid (a 1:50:50 (w:w:w) mixture of fluorescein-DHPE, DOPE and DOPG) in chloroform was infused to flush away the first aqueous solution. After the flushing, water-in-chloroform droplets were formed in the individual chambers. The lipid solution used in this process contained a small amount of lipid, i.e., approximately one thousandth of that used in conventional procedures to form lipid bilayers (~10 mg mL−1)13,14,15 but almost the same amount as that previously employed for lipid monolayer formation at the interface of water-in-oil droplets (~50 μg mL−1)5,16. Therefore, lipid monolayers but not multilayers were presumably formed at the interfaces of individual droplets. Third, a second lipid solution containing 2 mg mL−1 lipid (a 1:1 (w:w) mixture of DOPE and DOPG) in chloroform was infused. Finally, a second aqueous solution was infused to flush the second lipid solution. In this process, the hydrocarbon tails of the residual second lipids zipped with those of the first lipids to form a bilayer membrane. Uniform-sized planar bilayers with an asymmetric lipid composition where fluorescein-DHPE was located only in the inner leaflet of bilayer were accordingly formed on the opened orifice of the micro-chambers (Fig. 2b), with a formation efficiency of over 97% (number of chambers sealed with lipid membranes: 10,545 versus total number of chambers: 10,784, yielding 97.8% efficiency). Here, it should be noted that in this study, we formed lipid bilayers using chloroform as a solvent for the lipid, whereas decane or hexadecane has been used in conventional approaches8. The prominent feature of our approach is that the lipid membranes spontaneously thin down to form bilayers without mechanical perturbation, e.g., hydraulic pressure, which is required for bilayer formation using decane or hexadecane. This mechanical perturbation is a critical issue because it breaks more than half of the lipid membranes, thus resulting in a low efficiency of less than 30% for bilayer formation in the previous approaches16. The thinning of the lipid membranes is driven by drainage of the organic solvent over the hydrophobic support. Chloroform has higher water solubility than decane or hexadecane and is thus more easily dissolved in an aqueous solution, i.e., it is more easily drained over a hydrophobic support and thus, our approach does not require any mechanical perturbation, which contributes to increasing the efficiency of bilayer formation to over 97%. Bright-field observation showed circular interference patterns in the individual chambers after lipid membrane formation (Fig. S1b), whereas a clear circular pattern was not observed before membrane formation (Fig. S1a). This interference pattern is a characteristic optical feature of thin lipid bilayer formation, termed the Plateau-Gibbs border17,18, which separates the thin lipid bilayer and the supporting annulus. Thus, bright-field observation supported the formation of lipid bilayers in this study. Lipid bilayer formation was further confirmed from the activity of a membrane transporter, α-hemolysin19 (see below), which is not functional unless the lipid membrane becomes as thin as the membrane-spanning structure of α-hemolysin.

Formation of lipid bilayer membrane with an asymmetric lipid composition.
(a) Schematic illustration of asymmetric lipid bilayer formation. First, an aqueous solution is injected into the device through an access port. Second, the lipid solution containing fluorescent-labeled lipids (fluorescein-DHPE) is injected. Third, another lipid solution without fluorescein-DHPE is injected. Finally, a second aqueous solution is injected to flush the second lipid solution. In these processes, asymmetric lipid bilayers are formed on the orifices of micro-chambers. (b) Fluorescent image of fluorescein-DHPE composing asymmetric lipid bilayers.
FRAP measurement
The planar lipid bilayers formed in this study were isolated and remained independent of each other, as confirmed by fluorescence recovery after photobleaching (FRAP). As mentioned above, the lipid bilayers formed in this study were composed of fluorescein-DHPE. Thus, fluorescent images of fluorescein-DHPE-containing lipid bilayers were recorded with 10-s intervals by confocal microscopy (Fig. 3). Using the laser scanning system of the confocal microscope, one of the bilayers was selectively photobleached (Fig. 3). A subsequent recording of the fluorescence intensity showed that the bleached bilayer did not recover its original fluorescence, suggesting that the fluorescein-DHPE did not laterally diffuse between bilayers within the 700-s observation period (Fig. 3). Thus, the individual lipid bilayers formed in this study were confirmed to be isolated from each other, i.e. more than 10,000 lipid bilayers were formed at a time on our micro-device.

FRAP measurement of fluorescein-DHPE.
Representative time courses of the fluorescent intensity of fluorescein-DHPE-containing lipid bilayers (gray) and their average (green). The fluorescent intensity was normalized against that at 0 s. The upper panels show fluorescent images of fluorescein-DHPE before and after photobleaching.
Confirmation of lipid compositional asymmetry
To confirm the lipid compositional asymmetry between two leaflets, a pH-jump assay was conducted using fluorescein-DHPE, a pH indicator in which the fluorescence intensity decreases upon acidification (Fig. 4a). Initially, micro-chambers and an upper flow channel were filled with basic buffer (HEPES buffer, pH 8.0). Then, the buffer in the flow channel was exchanged with acidic buffer (MES buffer, pH 6.0). Before and after the buffer exchange, the fluorescent intensity of fluorescein-DHPE was stable (Fig. 4b,c), demonstrating that the hermetic seal of the micro-chamber with a lipid bilayer was sufficiently tight to prevent proton leakage and moreover, fluorescein-DHPE was not located in the outer leaflets of the bilayers, which were exposed to the flow channel. Next, we injected an acidic buffer containing 5 μg mL−1 α-hemolysin, a toxic membrane protein that binds to lipid bilayer membranes and forms transmembrane nanopores19 (Φ = 1~2 nm) that have been used for various biomedical sensors, e.g., nanopore DNA sequencing20. Because α-hemolysin acts as a transporter only when the nanopore penetrates the lipid membrane, the transport activity of α-hemolysin is often used to confirm the formation of a thin lipid bilayer. Accordingly, it is highly expected that the buffer encapsulated in the micro-chambers would be exchanged with that in the flow-channel through nanopores of α-hemolysin if the lipid membranes formed in this study are thin bilayers. After injection of the acidic buffer containing α-hemolysin, the fluorescence intensity of the entire membrane uniformly decreased toward that at pH 6 (Fig. 4c), showing that the lipid membranes that formed in this study were thin enough to functionalize membrane protein activity and moreover fluorescein-DHPE was uniformly located on the entire inner leaflet of bilayers. Thus, the formation of highly asymmetric bilayers was confirmed in this study.

Demonstration of lipid compositional asymmetry in the two leaflets.
(a) Fluorescence intensity of fluorescein-DHPE versus pH. (b) Schematic illustration of pH-jump assay. First, micro-chambers and an upper flow channel are filled with basic buffer (pH 8). Second, the buffer in the flow channel is exchanged with an acidic buffer (pH 6). Third, the α-hemolysin dissolved in acidic buffer is injected into the flow channel. During these processes, we measured the fluorescence intensity of fluorescein-DHPE located in the bilayers. (c) Time course of fluorescence intensity of fluorescein-DHPE in bilayers. Green or grey plots represent the fluorescence intensity in the presence or absence of α-hemolysin after 2000 s. Insets show fluorescent images of fluorescein-DHPE in the representative micro-chamber.
From the pH-jump assay in the absence of α-hemolysin, we investigated the stability of the lipid compositional asymmetry over time. As shown by the gray plot in Fig. 4c, the fluorescence intensity of fluorescein-DHPE located in the inner leaflet of the bilayer remained stable for 4,000 s, which shows that flip-flop of fluorescein-DHPE in the bilayer rarely occurred for 4,000 s.
Flip-flop measurement
Instead of fluorescein-DHPE, we used NBD-DPPE to form asymmetric lipid bilayers, where NBD-DPPE is located in the inner leaflet of the bilayer. NBD-DPPE is a head-group-labelled fluorescent lipid that has been widely used as a model lipid for flip-flop measurement21,22,23 (Fig. 5a). To visualize the flip-flop of NBD-DPPE, we performed the NBD-reduction assay using dithionite, which rapidly quenches the fluorescence of NBD21 (Supplementary Information). Initially, the micro-chambers and an upper flow channel were filled with buffer (MOPS buffer, pH 7.0). Then, the buffer in the flow channel was exchanged with the buffer containing 50 mM dithionite. Because dithionite does not permeate through lipid membranes on our system for 7 h (Supplementary Information), the fluorescence of NBD-DPPE will be quenched only when NBD-DPPE is exposed to upper flow channel upon flip-flop. After the buffer exchange, the fluorescent intensity of NBD-DPPE was gradually and exponentially decreased with a time constant of 4.1 h (Fig. 5b,c), which was similar to previous findings in liposomes21. These findings suggest that the fluidity of the lipid in our device, which affects the flip-flop rate, is almost the same as that in liposomes.

Flip-flop measurement using NBD-DPPE.
(a) Quenching of NBD-DPPE resulting from reduction by dithionite. (b) Schematic illustration of NBD-reduction assay. First, micro-chambers and an upper flow channel are filled with buffer (pH 7). Second, the buffer in the flow channel is exchanged with buffer containing 50 mM dithionite. During these processes, we measured the fluorescence intensity of NBD-DPPE located in the bilayers. (c) Time course of fluorescence intensity of NBD-DPPE in bilayers. Green or grey plots represent the fluorescence intensity in the presence or absence of 50 mM dithionite after 40 min, respectively. The orange line represents the fitting curve based on a single-order reaction scheme with a time constant of 4.1 h. Insets show fluorescent images of NBD-DPPE in the representative micro-chamber.
Discussions
In this study, we succeeded in demonstrating real-time flip-flop measurement in planar lipid bilayers, which has been difficult in other membrane systems because of the fast leakage of dithionite, i.e. dithionite can cross the lipid bilayer within an hour21. As mentioned above, our system achieves tight sealing of lipid bilayers, which prevents dithionite leakage across the bilayer for seven hours (Supplementary Information) and therefore, allows us to conduct real-time monitoring of lipid flip-flop in a single planar lipid membrane by using NBD-labelled phospholipid. Real-time monitoring of flip-flop in the single lipid membrane may have the versatile uses in various lipid and membrane protein assays, such as single molecule analysis of flippase activity and thus increase the understanding of the lipid flip-flop mechanism.
We constructed a micro-device for high throughput formation of asymmetric planar lipid-bilayers in micro-chamber arrays. Micro-chamber arrays have recently enabled the development of highly sensitive and quantitative biological assays, such as single-molecule enzymatic assays, digital PCR and digital ELISA9,10,24,25,26. Thus, the micro-device developed in this study has a large variety of potential applications, including highly sensitive analysis of membrane proteins on asymmetric lipid bilayers, which would enable further understanding of the role of lipid compositional asymmetry in the function of membrane proteins, which are optimal pharmacological targets. Additionally, the high-throughput feature of asymmetric bilayer formation in our micro-device may enable screening of pharmacological effects on membrane proteins located in the asymmetric lipid bilayers in vitro.
Methods
Microfabrication
We fabricated hydrophobic through-hole structures on a hydrophilic glass substrate using conventional photolithography. A carbon-fluorine hydrophobic polymer (CYTOP, Asahi-glass, Japan) was spin-coated on a clean cover glass (32 mm × 24 mm) at 4,000 rpm for 30 s and then baked for 1 h at 180°C. The thickness of the CYTOP layer was 0.5 μm. Photolithography was conducted using a positive photoresist (AZP4903, AZ Electronic Materials, Japan) to pattern mask structures on the CYTOP layer. The resist-patterned substrate was dry-etched with O2 plasma using a reactive ion etching system (RIE-10NR, Samco, Japan) to expose an array of hydrophilic SiO2 on the glass surface. The substrate was then cleaned and rinsed with acetone and ethanol to remove the photoresist layer remaining on the substrate. Figure 1 depicts a fabricated device, where the diameter of the CYTOP well is 4 μm.
Imaging
Fluorescent time-lapse recordings were taken under a confocal microscope system with a 60× objective lens and photomultiplier tubes (A1R, Nikon, Japan). A laser was used for the excitation of fluorescein-DHPE (λex = 488 nm). The fluorescence intensity was recorded through a 60× objective lens with photomultiplier tubes (Nikon, Japan). Analysis was performed using NIS Elements software (Nikon, Japan). Bright-field images of lipid membrane formation were taken under an optical microscope with a 100× objective lens (IX 71, Olympus, Japan).
Photobleaching assay
The flow cell and micro-chambers were filled with buffer A (50 mM HEPES [pH 8.0]). Using the laser-scanning system of the confocal microscope, one of the chambers was photobleached for a few seconds. The fluorescent images of fluorescein-DHPE were recorded at 10-s intervals.
pH-jump assay
The flow cell and microchambers were filled with buffer A. After lipid membrane formation, the buffer in the flow cell was exchanged to buffer B (50 mM MES [pH 6.0]). After a certain period had elapsed, the buffer in the flow cell was exchanged to buffer B containing 5 μg mL−1 α-hemolysin (Sigma Aldrich, USA). Fluorescence images of fluorescein-DHPE composing lipid bilayers were recorded at 60-s intervals.
NBD-reduction assay
The flow cell and microchambers were filled with buffer C (500 mM MOPS [pH7.0]). After lipid membrane formation, the buffer in the flow cell was exchanged to buffer C containing 50 mM dithionite. Fluorescence images of NBD-DPPE composing lipid bilayers were recorded at 5-min intervals.
References
van Meer, G., Voelker, D. R. & Feigenson, G. W. Membrane lipids: where they are and how they behave. Nat Rev Mol Cell Biol 9, 112–124 (2008).
Boon, J. M. & Smith, B. D. Chemical control of phospholipid distribution across bilayer membranes. Med Res Rev 22, 251–81 (2002).
Bratton, D. L. et al. Appearance of phosphatidylserine on apoptotic cells requires calcium-mediated nonspecific flip-flop and is enhanced by loss of the aminophospholipid translocase. J Biol Chem 272, 26159–65 (1997).
Fadok, V. A., de Cathelineau, A., Daleke, D. L., Henson, P. M. & Bratton, D. L. Loss of phospholipid asymmetry and surface exposure of phosphatidylserine is required for phagocytosis of apoptotic cells by macrophages and fibroblasts. J Biol Chem 276, 1071–7 (2001).
Pautot, S., Frisken, B. J. & Weitz, D. A. Engineering asymmetric vesicles. Proc Natl Acad Sci U S A 100, 10718–21 (2003).
Hwang, W. L., Chen, M., Cronin, B., Holden, M. A. & Bayley, H. Asymmetric droplet interface bilayers. J Am Chem Soc 130, 5878–9 (2008).
Syeda, R., Holden, M. A., Hwang, W. L. & Bayley, H. Screening blockers against a potassium channel with a droplet interface bilayer array. J Am Chem Soc 130, 15543–8 (2008).
Zagnoni, M. Miniaturised technologies for the development of artificial lipid bilayer systems. Lab Chip 12, 1026–39 (2012).
Watanabe, R. et al. Arrayed Lipid Bilayer Chambers Allow Single-Molecule Analysis of Membrane Transporter Activity. Nat Commun 5, 4519 (2014).
Kim, S. H. et al. Large-scale femtoliter droplet array for digital counting of single biomolecules. Lab Chip 12, 4986–91 (2012).
Sakakihara, S., Araki, S., Iino, R. & Noji, H. A single-molecule enzymatic assay in a directly accessible femtoliter droplet array. Lab Chip 10, 3355–62 (2010).
Iino, R. et al. A single-cell drug efflux assay in bacteria by using a directly accessible femtoliter droplet array. Lab Chip 12, 3923–3929 (2012).
Ota, S., Suzuki, H. & Takeuchi, S. Microfluidic lipid membrane formation on microchamber arrays. Lab Chip 11, 2485–7 (2011).
Sandison, M. E., Zagnoni, M. & Morgan, H. Air-exposure technique for the formation of artificial lipid bilayers in microsystems. Langmuir 23, 8277–8284 (2007).
Suzuki, H., Le Pioufle, B. & Takeuhci, S. Ninety-six-well planar lipid bilayer chip for ion channel recording Fabricated by hybrid stereolithography. Biomedical Microdevices 11, 17–22 (2009).
Yanagisawa, M., Iwamoto, M., Kato, A., Yoshikawa, K. & Oiki, S. Oriented Reconstitution of a Membrane Protein in a Giant Unilamellar Vesicle: Experimental Verification with the Potassium Channel KcsA. J Am Chem Soc 133, 11774–11779 (2011).
Fujiwara, H., Fujihara, M. & Ishiwata, T. Dynamics of the spontaneous formation of a planar phospholipid bilayer: A new approach by simultaneous electrical and optical measurements. J Chem Phys 119, 6768–6775 (2003).
Wang, W. et al. Activity monitoring of functional OprM using a biomimetic microfluidic device. Analyst 137, 847–852 (2012).
Song, L. et al. Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 274, 1859–66 (1996).
Branton, D. et al. The potential and challenges of nanopore sequencing. Nat Biotechnol 26, 1146–53 (2008).
Moreno, M. J., Estronca, L. M. & Vaz, W. L. Translocation of phospholipids and dithionite permeability in liquid-ordered and liquid-disordered membranes. Biophys J 91, 873–81 (2006).
Sampaio, J. L., Moreno, M. J. & Vaz, W. L. Kinetics and thermodynamics of association of a fluorescent lysophospholipid derivative with lipid bilayers in liquid-ordered and liquid-disordered phases. Biophys J 88, 4064–71 (2005).
Abreu, M. S., Moreno, M. J. & Vaz, W. L. Kinetics and thermodynamics of association of a phospholipid derivative with lipid bilayers in liquid-disordered and liquid-ordered phases. Biophys J 87, 353–65 (2004).
Rissin, D. M. et al. Single-molecule enzyme-linked immunosorbent assay detects serum proteins at subfemtomolar concentrations. Nat Biotechnol 28, 595–9 (2010).
Rondelez, Y. et al. Microfabricated arrays of femtoliter chambers allow single molecule enzymology. Nat Biotechnol 23, 361–5 (2005).
Ottesen, E. A., Hong, J. W., Quake, S. R. & Leadbetter, J. R. Microfluidic digital PCR enables multigene analysis of individual environmental bacteria. Science 314, 1464–7 (2006).
Acknowledgements
We thank all members of the Noji Laboratory. This work was supported by Grant-in-Aid for Scientific Research no. 18074005 to H.N. and no. 30540108 to R.W. from the Ministry of Education, Culture, Sports, Science and Technology, Japan and Precursory Research for Embryonic Science to R.W. from the Japan Science and Technology Agency.
Author information
Authors and Affiliations
Contributions
R.W. designed and performed experiments and analyzed data; N.S. and T.Y. gave technical supports; H.N. designed experiments, built whole story and wrote papers with R.W.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Watanabe, R., Soga, N., Yamanaka, T. et al. High-throughput formation of lipid bilayer membrane arrays with an asymmetric lipid composition. Sci Rep 4, 7076 (2014). https://doi.org/10.1038/srep07076
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep07076
This article is cited by
-
Compounds composition of pollen tubes of Scots pine (Pinus sylvestris L.)
Physiology and Molecular Biology of Plants (2023)
-
Membranes on nanopores for multiplexed single-transporter analyses
Microchimica Acta (2016)
-
Attolitre-sized lipid bilayer chamber array for rapid detection of single transporters
Scientific Reports (2015)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
