
Abstract
How do physical interactions with others change our own motor behavior? Utilizing a novel motor learning paradigm in which the hands of two - individuals are physically connected without their conscious awareness, we investigated how the interaction forces from a partner adapt the motor behavior in physically interacting humans. We observed the motor adaptations during physical interactions to be mutually beneficial such that both the worse and better of the interacting partners improve motor performance during and after interactive practice. We show that these benefits cannot be explained by multi-sensory integration by an individual, but require physical interaction with a reactive partner. Furthermore, the benefits are determined by both the interacting partner's performance and similarity of the partner's behavior to one's own. Our results demonstrate the fundamental neural processes underlying human physical interactions and suggest advantages of interactive paradigms for sport-training and physical rehabilitation.
Similar content being viewed by others


Introduction
From infancy, our motor behaviors are conditioned to respond and adapt to verbal, visual and haptic cues from other humans. These responses manifest as two types. First, cognitive responses related to the explicit knowledge of an interacting agent, defined by the so called theory of mind1,2,3,4,5 and second, motor responses that do not require an explicit interaction with another individual but are driven by the sensory feedbacks characteristic of an interaction. For example, hearing a scream on the radio makes you feel scared and tenses your muscles while looking at a happy face on television implicitly induces a smile on your face. These responses and the consequent motor adaptations are arguably crucial indicators of the fundamental interactive sensory-motor associations that drive human interaction behaviors6.
However, while human interactions have been systematically studied over the last decades across fields such as theory of mind1,2,3,4,5, mirror neurons7,8,9, child development10, observational learning11,12,13 and joint action14,15,16,17,18,19,20, the relative contributions of the cognitive and motor responses to these behaviors are still largely unclear. This is particularly true about physical interactions, the understanding of which has developed mainly in the last decade14 due to the technical difficulties associated with the observation and analysis of the key determinant of physical interaction - haptic feedback14,15.
This study investigates the motor responses and the consequent adaptations that govern physical interactions between humans. For this purpose, we developed a novel interactive learning paradigm using a dual robot system in which pairs of individuals are physically connected during a motor task without conscious knowledge of the connection. In contrast to previous joint action studies14,15,16,17,18,19, this paradigm enabled us to investigate the reactive motor adaptations driven by the haptic signals during physical interaction without complications from cognitive adaptations related to conscious coordination with the partner.
Interestingly, we observed that physical interactions are consistently beneficial to the interacting individuals and enable them to improve their motor performance both during and after interactive practice. We show that these benefits are present only in physical interaction with an active partner and cannot be explained by multisensory integration of the visual and haptic sensory information in an individual21. Furthermore, we modulate the interaction characteristics and partner behavior to show that both the quantitative partner performance and qualitative performance (or nature) affect the benefits experienced during physical interaction. Our results reveal fundamental sensory-motor mechanisms active during inter-personal physical interactions that are distinct from the cognitive interactive mechanisms studied previously as theory of mind.
Results
Interaction experiment
Subjects participated as pairs (or dyads) in our interaction experiment. Each subject held one of two handles of the dual-robot interface (Fig. 1A) and was provided with continuous visual feedback of only his own hand position as a cursor on his monitor. Dyads were required to track a moving target on their respective screens over 60 one-minute trials grouped in four sessions of 10, 20, 20 and 10 trials respectively. Though the target movement changed across trials, the same target was presented for both subjects in a dyad during every trial. A visuo-motor rotation was introduced in the two middle sessions (total 40 trials) in which the cursor movement on the screen was rotated 80° clockwise with respect to hand movement. The subjects were naïve to this new environment and had to learn to work in it by practicing the tracking task.

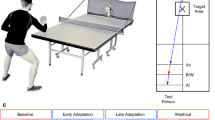
(A) Setup: The figure shows a cartoon of the setup used for the experiment.
Subjects worked in dyads, each holding a handle of the robot interface. They tracked a moving red target on their respective monitor with a cursor (white dot) representing their hand position. They had no visual information of their arm which was covered by the table. The target movement was the same for both subjects in the dyad. In the dual trials, their hands were connected by a virtual elastic band (represented in brown) such that each subject was pulled towards the hand position of the partner. The subjects could see their partner but could not see the partner's hand or monitor. (B) Improvement during interaction: The improvement in task performance in each subject for each dual trial was plotted against the relative performance of their partner. The dual trial improvement was measured by the change in tracking error by a subject during a single trial compared to his individual tracking error in the immmediatly preceding dual trial. It is observed that when connected to a better performing partner (+abcissa), an individual's performance improved. Interestingly, the individual performance improved even when connected to a partner with inferior performance in the task (−abcissa). (C) Learning during interaction: The improvement in task performace (relative to the first trial) across the single trials of the interacting subjects (green trace) in the visuomotor learning sessions was compared to improvement of task performance by solo subjects (red trace) who never interacted with a partner. Note that the trace combines single trials across subjects such that each data point and error bar represents single trials made by ten of the (total of 20) subjects. Intermittent interaction enabled significatly higher motor learning in individuals.
Each experimental session consisted of an equal number of single and dual trials. In the single trials each individual performed the tracking alone, while in the dual trials the robotic interface implemented a virtual (compliant) elastic band (see cartoon of connection shown in brown in Fig. 1A) that physically connected the hands of the individuals in a dyad during the tracking task. The sequence of the trials was pre-determined before the experiment and was unknown to the subjects. Half of the dyads started with a single trial while the rest started with a dual trial. The subjects were not informed on the nature of the forces they experienced in the dual trials and were not consciously aware of the connection with the partner, as was verified by a questionnaire at the end of the experiment.
Although the subjects could feel forces on their hands in the dual trials, the connection compliance allowed them to create their own independent movement. The relatively large compliance (elastic band stiffness was either 60, 120 or 180 N/m) also prevented them from relaxing and letting the forces guide their hand. The subjects had to actively track the target to perform the task. Our experimental paradigm thus differed from previous studies14,15,16,17,18,19,20 in two aspects. First, unlike most previous studies which ask subjects to perform a single task together, subjects in this paradigm performs their own individual tasks. Second, most previous physical interaction studies used tasks in which the partners' limbs were rigidly coupled. In contrast, the two partners in our experiment could perform their own task independently while being connected through a compliant link.
Fig. 1B summarizes the performance, defined as the mean distance from the target over a trial, of 10 dyads (20 subjects) who participated in the main interaction experiment. The figure plots the performance improvement in each subject in the dual trials against the relative performance of his dyad partner. The positive abscissa plots the dual performance improvement for a subject when connected to a partner with superior performance in the task, while the negative abscissa plots the dual performance improvement when the subject was connected to a partner with inferior performance in the task. We observed that the performance of a subject in a dual trial improved when his interacting partner was better than him and that the improvement increased as the performance of the partner increased (R = 0.79, deg. of freedom (n − 2) = 198, p < 10−11, in first quadrant of Fig. 1B). Remarkably, there was an improvement in performance even when coupled to an inferior performer (T(199) = 28.21, p < 10−11, T-test of data points in second quadrant of Fig. 1B). Therefore, connection with a partner improved the task performance in an individual irrespective of whether the partner performance was better or worse than the individual's own performance.
We next analyzed motor learning by examining the evolution of single trial performance of subjects across the two middle sessions where they learned the visuo-motor rotation (green trace in Fig. 1C). We compared this learning performance to that of a separate group of solo subjects (red trace in Fig. 1C) who learned the same task for the same duration but without ever interacting with a partner. It was observed that subjects achieved significantly better performance if they intermittently interacted with a partner compared to subjects who practice the task alone (2 way ANOVA exhibited a main effect of of solo/interaction conditions, F(1,759) = 21.86, p = 3 × 10−4; and main effect of trials F(39,759) = 21.4, p < 10−5).
These surprising results show that the physical connection can lead to motor adaptations in interacting humans even when they are unaware of the presence of a human partner. Interestingly, these adaptations are mutually beneficial. They enable individuals to consistently perform better in their own task (Fig. 1B) during interaction and also helped them to learn a new motor task better than subjects who practiced the task alone for the same amount of time (Fig. 1C; the learning curves of the interacting and solo subjects are presented in supplementary Fig. S1). In order to determine the key factors that enable these benefits during interaction, we varied the behavior of the interacting agent using our robot interface and examined how this affected the interaction behavior.
Control experiments
We first verified if the performance improvement in the dual trials was due to the presence of interaction forces and did not specifically require the forces to be from an interacting partner. The interaction forces could have led to increased system stiffness or subject attention, which in turn could have assisted performance improvement in subjects. An alternative explanation could be that the subjects monitored the interaction forces and chose to follow the forces that contribute to improved task performance while resisting forces that hinder performance. These possibilities were excluded by the results of a control force-playback experiment with a separate group of dyads. While dyads in the force-playback experiment also performed both single and dual trials like the main interaction experiment, the dual trials in force-playback did not involve connection to the partner. Instead, in the dual trials each individual experienced the recorded connection forces from subjects of a previous dyad who had tracked the same target in the interaction experiment. The subjects under force-playback thus experienced forces of similar magnitude, frequency and target dependencies as subjects in the interaction experiment, though the forces were unrelated to their partner's performance. We observed that the dual trials in force-playback consistently degraded the performance of both the interacting partners (dark green trace in Fig. 2). This behavior was significantly different from that of the interaction experiment (F(1,552) = 422.21, p < 10−11, 1 way ANOVA between the black and green trace).

Influence of interaction characteristics.
The data from the main interaction experiment (black trace), force-playback control (green trace), trajectory-playback control (orange trace), target-connect (purple trace) and expert-connect (blue trace) were plotted in the same format as Fig. 1b. The positive- abcissa plots an individual's performance improvement when connected to a partner with superior performance in the task. The negative abcissa plots an individual's performance improvement when connected to a partner with inferior performance in the task. We compared the overlapping abscissa between any two partner conditions and observed the behavior in the control experiments (green and orange traces) to be significantly different from the interaction experiment. While connection to the target (purple) or an expert human (blue) were qualitatively similar to connection to a novice human (black trace), they were still quantitatively different, indicating that both the performance and nature of the partner are important determinants of interaction performance in humans.
A more plausible explanation for the mutual benefits during interaction is information integration from multiple senses. While in the single trials the subjects performed the tracking task using only the visual feedback of the target, in the dual trials they also received haptic feedback from the partner performing the same task. Their central nervous system (CNS) can integrate these two feedback signals21,22 and this integration may have been the reason behind the improved task performance in the dual trials. However, if sensory integration was indeed the primary cause of the improved performance, then one could suppose that direct interaction with a partner is not a necessity for performance improvement. Any additional task relevant haptic information should enable subjects to improve performance in the interaction experiment.
To check this possibility, a second control experiment was performed with a separate group of dyads. In this trajectory-playback experiment, instead of being connected online to a partner, each individual was connected, by means of a virtual elastic band similar to the interaction experiment, to the recorded trajectory (time series of position coordinates) of a previous solo subject who tracked the same target alone. Therefore subjects in the trajectory playback experiment could receive haptic feedback of their partner's tracking behavior. Trajectory playback improved the subject performance (Fig. 2, orange trace, 1st quadrant) when the connected solo subject was better, though significantly less than in the interaction experiment. However, if the subject was already better than the connected solo subject, then his performance deteriorated. A 2 way ANOVA between the main and trajectory playback conditions and relative partner performance shows the conditions affect the behavior (F(1,596) = 171.44, p < 10−11), the partner performance affects the behavior (F(7,596) = 22.39, p < 10−11) and significance interaction (F(7,595) = 13.15, p = 8.8 × 10−15) in the dual trials. Overall, this behavior was distinctly different from that of our first interaction experiment (as shown by the interaction result of the 2 way ANOVA), demonstrating that the performance improvement during interaction was not a mere effect of multi-sensory integration.
To understand what prevented the dyads in the control experiments from improving as in the interaction experiment we analyzed which signal was missing in the two control experiments relative to the interaction experiment. In both control experiments the dyads received visual feedback as well as relevant force feedback from the movements of a partner doing the same task. However, force-playback and trajectory-playback presented a one–way connection where a subject could feel and respond to the actions performed by the partner, but the partner (who was a previous subject) could not do the same. In contrast, in the interaction experiment a two-way connection enabled each individual of the dyad to receive both haptic feedback of their partner's behavior and haptic reactions from the partner in response to their own behavior. This two-way connection and specifically the partner reaction, seems to be the essential factor that induced mutual benefits and which was absent in both control experiments. If so then it would suggest that during physical interactions, individuals have implicit expectations, in terms of the haptic forces from a partner, even when they are not consciously aware of his presence.
Effect of partner nature
It is well known that cognitive interactions between humans are modulated by their expectations of their partner's behavior1,2. Although similar expectations have been suggested to affect physical interactions23,24, evidence in this regard has remained absent. In order to verify that partner reaction is indeed a key factor affecting the benefits we observed during physical interaction first, consistent with previous suggestions25,26,27,28, we hypothesized that the partner would be modeled with behavior similar to one's own. In this case we can make two predictions. First, if particular haptic reactions are expected, then the interaction behavior should change with the quantitative changes in haptic reaction forces. Second, the interaction benefits observed in our study should change with an individual's ability to model partner behavior. A human partner should be more beneficial than a non-human partner because a human partner can arguably be modeled more accurately. Furthermore, a human partner using similar control strategies should be more beneficial than a human partner using different strategies.
To examine whether and how, the quantitative characteristics of the partner reaction affects the behavior, the connection stiffness was systematically varied in the interaction and trajectory-playback experiments. Differences were observed during interaction with a superior partner (first quadrant of Fig. S2). While dual performance was observed to improve with partner performance in each case, improvement in the dual trials was maximal at the connection stiffness of 120 N/m and less at both higher (180 N/m; 2 way ANOVA revealed main effect of partner performance F(4,126) = 43.62, p < 10−6 and main effect of connection stiffness F(1,126) = 29.35, p = 2.9 × 10−5) and lower (60 N/m; 2 way ANOVA revealed main effect of partner performance F(4,101) = 29.26, p < 10−3 and main effect of connection stiffness F(1,101) = 6.11, p = 0.01, F(4) = 29.26, p < 10−3) values of stiffness. These differences were observed only in the interaction experiment which involves partner reaction, but not in the trajectory playback experiment which does not involve a partner reaction.
To examine interaction with a non-human agent, a target-connect experiment was carried out in which subjects were connected with the same virtual elastic band as the human interaction experiment (Fig. 2A), but this time directly to the target. This corresponds to the control strategy employed during skill training and physical rehabilitation with robots29. It was observed that, although the target-connect condition provides the best available information on target position, it produced relatively less improvement in performance than connection to a novice human (compare black and pink trace in Fig. 2, F(1,207) = 34.64, p = 1,5 × 10−8, 1 way ANOVA).
The motor control literature has shown that task control mechanisms and accuracy in humans change when one becomes an expert in a task30. Interaction between subjects with different control strategies was thus tested in an expert-connect experiment in which dyads were composed of a novice subject and an expert in the tracking task. The expert in our experiment could track the target movement with little error (1.3 cm average single trial error compared to 5.1 cm for novice subjects). Counter-intuitively, but consistent with our prediction, at any relative performance level, the performance improvement in the novice subjects due to connection with an expert was less than with a novice partner (compare black and cyan trace in Fig. 2, F(1,501) = 147.92, p < 10−11, 1 way ANOVA). Furthermore, even though an expert made occasional errors (unlike the perfect target connection), an expert was observed to be better than direct connection with the target (Fig. 2; F(1,305) = 76.38, p < 10−11, 1 way ANOVA across the target and expert partner conditions).
These results demonstrate that in addition to partner performance (ordinate of Fig. 2), the nature of the partner's behavior (black, cyan, pink traces in Fig. 2) affects the dual trial benefits. While the definition of the behavioral nature of a partner is still unclear, using a model based analysis (see methods) we confirmed that the performance differences between the novices, experts and the target were not the cause of the differences observed with changes in partner nature. On the other hand, movement jerk31 was observed to be different among the novice, expert and target and was one of the possible factors responsible for determining the nature of partner behavior. Further studies are required to clarify all the factors that determine the nature of a partner's behavior.
Discussion
Using a dual robotic system, we created an interaction task in which the individuals were unaware of the presence of a partner. In our experimental paradigm, the two partners, could perform their own task independently while being connected through a compliant link. This setup enabled us to analyze the effect of motor adaptations during physical interactions between two individuals practicing a novel motor task. We showed that a physical connection between two individuals learning a motor task consistently improves their performance, regardless of their partner's performance (Fig. 1B, black trace in Fig. 2). Furthermore, intermittent connection with a partner enabled individuals to learn the task better than subjects who practiced the task alone for the same duration (Fig. 1C). These results indicate that physical connection enables individuals to attain additional task related information from their partner, a result which is consistent with the proposal of a haptic channel by a recent study15. We then demonstrated that the dual trial improvement depends not only on the relative partner performances (as has also been previously observed in interactive decision making32), but also on the nature of the interacting partner (see black, cyan, purple traces in Fig. 2). The improvement of performance is most prominent when the partners are similar, such that interaction with a human is more beneficial than with a non-human agent and interaction with a peer is more beneficial than with an expert.
Our results have several important implications. First, they indicate that physical assistance during rehabilitation and training can benefit from mutually interactive control modalities. A simple physical connection between individuals practicing sports or between patients during rehabilitation may increase performance more than that achieved by practicing the task alone for the same time. Furthermore, with robots seeing increased use for stroke rehabilitation and physiotherapy29,33 it has been a challenge for roboticists to develop robot behaviors that can assist patients similar to physiotherapists. Current robot assistance relies on motion guidance where robots guide and assist the patient's movement along a predefined trajectory. Our results predict that robots can improve performance and learning in patients by providing explicit reactions to patient behaviors during interaction. Finally, our results show for the first time that, similar to visual and auditory feedback, haptic feedback can lead to motor adaptations in humans that do not require explicit interaction with a partner. This result is in agreement with observations of spontaneous synchrony in brain activity during visuomotor interactions34. The existence of specific haptic expectations during physical interactions supports the presence of forward models24 of interacting agents at a much lower level of motor interaction than previously observed1,2,3,4. Further studies are required to clarify how the motor adaptations we observed here interact with the cognitive responses by the subjects, but it is interesting to note that the motor adaptations, driven solely by haptic feedback during physical interactions, are beneficial to individuals. While the exact mechanism by which the haptic expectation enables the benefits is still unclear, our results provide possible new insights into the phylogenesis of motor development; the urge in children to indulge in interactive games involving physical contact may correspond to a natural process to increase success in motor tasks and facilitate learning.
Methods
Subjects and task
74 subjects, aged 25–42 years, including 34 females, participated in pairs or dyads in the interaction experiment or in one of four subsidiary experiments. All participants were right-handed as assessed by the Edinburgh Handedness Inventory (Oldfield, 1971). The subjects gave informed consent for their participation in the experiments which were conducted according to the principles in the Declaration of Helsinki and approved by the ethics committee at Advanced Telecommunication Research Institute (http://www.atr.jp/). Table 1 summarizes the subject distribution and experiments. The dyads carried out planar arm movements with the dual robotic interface (Fig. 1A). Each subject held a manipulandum handle under the table and the room light was switched off so that the moving arm was not visible during the experiment. The subjects were provided with visual feedback of his hand position by a cursor on an individual computer screen in front of him. Each subject could see the other person sitting in front of him but could not see the hand movements or computer screen of the partner.
The subjects tracked a moving target on the visual screen in consecutive 60-second trials. After each trial, the target was switched off and the hand position was passively returned to the center of the screen by the manipulandum followed by the start of the next trial after a short 20–30 second break. The target movement was defined (in cm) by the multi-sine function

The trial time (t) was defined over a period of 0–80 seconds from which a period of 60 seconds was randomly chosen for each trial:

The target movement was thus different in each trial and did not repeat during one trial. This helped prevent fast learning of the target movement by the subjects.
The tracking task required hand movements over a circular workspace with a 30 cm diameter with average and maximum hand speeds of 0.18 m/s and 0.38 m/s, respectively. In every trial, both individuals of a dyad tracked the same target.
Each dyad performed 4 sessions of 10, 20, 20 and 10 trials with a five minutes rest between sessions. In the second and third sessions, a clockwise visual rotation of 80° was introduced between the subject hand movement and the cursor movement on the screen (visuo-motor rotation). This learning paradigm allowed analysis of the interaction effect in subjects with different levels of motor skill.
The concept of visuo-motor rotation was explained to the subjects before the experiment. The subjects were told that the aim of the experiment was not to find out the angle of rotation but to track the target. The subjects were asked not to stop and think but learn the rotation using smooth and continuous movements. To prevent competitive behaviors, the two subjects were told that they work on the same task but that it was a non-competitive task. Individuals of every dyad were chosen of the same gender in order to prevent any gender-related effects.
The entire experiment lasted little more than 2 hours including time for the instructions, breaks of about 20 ~ 30 seconds between trials and longer breaks of about 3–5 minutes between sessions. The subjects performed the tracking task for 60 min (60 trials × 1 min) within this 2 hour period.
Dual trials
In all but the solo experiment, each session included 50% dual trials distributed across the sessions in which the partners were haptically connected to each other. Subjects were informed about the dual trials as random trials “involving some external forces that would sometimes help the task and sometimes disturb it.” They were not given any explicit information about the connection.
Different connection conditions were used in the different experiments:
In the dual trials of the main interaction experiment, each partner's right hand was connected with an elastic force

produced by the manipulandum controlled at 2 KHz, where FSx and FSy are the x and y forces experienced by a subject at position (xS,yS) when the partner is at position (xP,yP). The stiffness K was set as either 60 N/m, 120 N/m, or 180 N/m (Table 1). A small damping D = 7 Ns/m was used in all experiments. The origin of the coordinate frame of each subject coincided with the center of the display screen such that if the two partners moved along the same trajectory in their respective coordinates they would experience zero force, while if one moved away from the other's position, both experienced an elastic force toward the other.
In the dual trials of force-playback control experiment, the two subjects performing the experiment were not connected together (as in the interaction experiment), but the robot applied (played back) connection forces recorded while a previous dyad performed the interaction experiment tracking the same target movement.
In the dual trials of the trajectory-playback experiment, each subject was connected to the recorded trajectory of a solo subject who had performed the experiment with the same target trajectory, but without ever connecting to a partner. That is, the partner's current position (xP,yP) in Eq. 3 was replaced by (xl,yl), the recorded position time series of a solo subject. Similar to the interaction experiment, three groups of dyads participated in trajectory-playback and experienced an elastic connection with stiffness K = 60 N/m, 120 N/m or 180 N/m, respectively.
In the target-connect experiment, the subjects were connected directly to the target in the dual trials with a stiffness of K = 120 N/m, effectively providing ‘perfect’ guidance by the robot. That is, the partner's current position (xP,yP) in Eq. 3 was replaced by (xt, yt), the current position of the target.
Finally, in the expert-connect experiment, while the subject and experimenter were connected in the same way as in the interaction experiment, the expert tracked the same target in a normal (non-rotated) environment. Furthermore, due to previous exposure, the expert was also familiar with the target movement patterns and produced an average single trial error of 1.3 cm compared to 5.1 cm by novice subjects. All experts were naïve to the purpose of the experiment. However, we tested effect of the expert's knowledge of interaction. In 5 dyads both the expert and the novice were unaware of the interaction with each other while in the other 5 dyads only the expert (and not the novice) was made aware that he interacts with the partner. We found no significance differences in the benefits observed in the novice in the two groups (p = 0.70, 2-way ANOVA) and therefore combined the data from both groups in Fig. 2.
Trial sequence
The experiment consisted of a total of 30 single and 30 dual trials distributed over four sessions. We utilized either of two predetermined random trial sequences across all dyads. Each sequence consisted of an equal number of single and dual trials in each session. One sequence started with a single trial in each session and was used for roughly half of the dyads in the interaction, force-playback, expert-connect and target-connect experiments. The other dyads in these experiments started the sessions with dual trials and an inverted sequence formulated by interchanging the single and dual trials of the first sequence. All subjects for the trajectory-playback experiment started with a single trial due to safety considerations. The two sequences used in our experiments were
SEQUENCE-1: {SDSDS SDDSD},{SSDDSDSDSDDSSDSSDDSDSSDDS DS DSDDSSDSSDDSD}, {SDSDS SDDSD}, and
SEQUENCE-2: {DS DSDDS SDS},{DDS SDS DS DSSDDSDDSDS DDS SDS DS DSSDDSDDSDS},{DS DS DDSSDS}, where the middle two sessions with visuo motor rotation have been shown grouped together.
Questionnaire
At the end of the experiment, each subject was asked three questions:
-
“Did you realize what the connection forces were?”
-
“Did you perform better in the presence or absence of interaction forces?”
-
“Did you feel fatigue during the experiment?”
Across the interaction, force-playback, trajectory-playback and expert-connect experiments, only two of 54 subjects realized what the connection forces represented. All but these two subjects indicated that they were completely unaware of the connection to the partner. Interestingly, all subjects believed they performed worse in the dual trials when there were external forces, though the results indicated a performance improvement in the dual trials of some experiments. These results were different in the target-connect experiment. Most subjects in the target-connect experiment realized that they were being pulled toward the target. All felt that the dual trials were easier to perform.
In all of the experiments, the subjects confirmed that they did not feel fatigue. However many subjects complained that their eyes became tired due to the continuous concentration on the moving target.
Plots and statistics
The performance of each subject was measured as the mean distance between the subject position and the target across every trial (recorded at 2 KHz). To evaluate how interaction affects each individual, the tracking error from each subject in every single trial was compared to the error in the previous dual trial. The difference of the two indicated the improvement experienced by the subject when connected to a partner.
It should be noted that we compare with the previous and not subsequent dual trial in order to counter the effects of motor learning on the performance improvement measured through our experiment. Using the previous dual trial in quantifying the improvement resulting from the connection thus represents a conservative estimation of the improvement excluding learning. The actual improvement due to the interaction is in fact larger than what we measure.
The average improvement in each dual trial was plotted against the difference of the single trial performance between the partners in Fig. 1B. The positive abscissa shows the improvement in a subject when interacting with a partner who was better in the single trial, while the negative abscissa represents the performance improvement in a subject when interacting with a partner who demonstrated inferior performance than him in the single trial. As we compare a single trial to an immediately previous dual trial, we isolated pairs of dual followed by single (DS) trials in the trial sequence presented to each subject. SEQUENCE-1 provides 22 DS pairs (shown in bold in the sequence above) and SEQUENCE-2 yields 18 pairs. Therefore, in total we get 22 × 10 points from 10 subjects (5 dyads) and 18 × 10 points from the other half of the subjects. Fig. 1B thus has 400 points in total. The average of these points across bins of 1 cm on the abscissa is shown by the thick black trace in Fig. 1B which is redrawn in Fig. 2.
To evaluate the improvement in an individual's task performance after interaction (their skill level), we examined the average decrease in task error through the single trials in the interacting subjects (green trace in Fig. 1C) and compared this to the error of a separate group of solo subjects (red trace in Fig. 1C) who performed the same task without ever interacting with a partner. As the two trial sequences (see Trial sequence subsection) were made by interchanging the single and dual trials between them, each green data point and error bar in Fig. 1C corresponds to ten (of the 20) subjects. One way ANOVA across the trials and the interaction condition was used to exhibit the improvements due to interaction.
A procedure similar to Figure 1B was used to quantify the improvement in the force-playback (green trace in Fig. 2), trajectory-playback (orange trace in Fig. 2), target-connect (purple) and expert-connect (cyan) experiments respectively. For the force playback plot, the previous subject whose experienced force was played back, was regarded as the ‘partner’. Note that in the case of the target-connect and expert-connect experiments, the novice subject was the ‘inferior performer’ as the partner was either an expert or that target itself and the data from the novice thus resides predominantly in the positive abscissa. A 1-way or 2-way ANOVA was utilized to compare each experiment with the novice-novice experiment. In order to avoid any bias in the results due to the difference in the abscissa spreads across experiments, only the data with overlapping abscissa were considered while comparing any two experiments.
In addition, we conducted a model-based analysis to evaluate how the improvements are affected by the absolute performance levels of the partners in relation with the partner's nature (black, purple, cyan traces). The improvement I was regressed as a linear weighted summation of the difference in partner performance D (abscissa of Fig. 2) and the partner performance error E, as well as functions Fj of the partner nature, where j is one of the subscripts p = peer novice, t = target or e = expert:

Three different function types (Fj) were examined to represent the partner nature:
-
Indicator functions {δp, δt, δe} on the set {p, t, e} such that δp(p) = 1 for peer novice partner, δp(t) = 0, δp(e) = 0 and similar for δt and δe.
-
The products {δpED, δtED, δeED} of the performance and difference of performance
-
The squares of performance differences {δpD2, δtD2, δeD2}
The best least-square fit was obtained using the squares of performance differences (R = 0.83, deg. of freedom (n − 2) = 996, p < 10−6) indicating that the linear model composed of this function Fj best represents the improvement data. The resulting coefficients were:
-
α = 0.195 ± 0.018 (95% confidence interval CI)
-
β = 0.18 ± 0.020 (95% CI)
-
γp = 4.295 ± 1.613 (95% CI)
-
γe = 2.085 ± 0.430 (95% CI)
-
γt = 1.587 ± 0.384 (95% CI)
Overall, we found that each of the variables {E, D, D2} contribute significantly to explain the variance in the data (F(996) > 35, p < 1e − 5). Importantly, γp > γe (p < 0.05, as seen from the CI above) and γp > γt (p < 0.05, as seen from the CI above) demonstrate that at a given (D,E) value, interaction with a peer novice leads to better performance than with an expert or target in the tracking task.
References
Adolphs, R. Cognitive neuroscience of human social behaviour. Nat. Neurosci. Rev. 4, 165–78 (2003).
Carruthers, P. & Smith, P. K. Theories of Theory of Mind (Cambridge University Press 1996).
Blakemore, S. J. & Decety, J. From the perception of action to the understanding of intention. Nat. Neurosci. Rev. 2, 561–7 (2001).
Frith, C. D. & Singer, T. The role of social cognition in decision making. Philos. Trans. R Soc. Lond. B Biol. Sci. 363, 3875–86 (2008).
Dunbar, R. [The social brain hypothesis and its relevance to social psychology]. Evolution and the Social Mind (New York: Psychology Press 2007).
Baron-Cohen, S. Mindblindness: an essay on autism and theory of mind (MIT press 1995).
Rizzolatti, G. & Craighero, L. The mirror-neuron system. Annual Rev. Neurosci. 27, 169–192 (2004).
Rizzolatti, G. & Sinigaglia, C. The functional role of the parieto-frontal mirror circuit. Nat. Neurosci. Rev. 11, 264–274 (2010).
Nojima, I., Mima, T., Koganemaru, S., Thabit, M. N., Fukuyama, H. & Kawamata, T. Human motor plasticity induced by mirror visual feedback. J. Neurosci. 32, 1293–1300 (2012).
Kovacs, A. M., Teglas, E. & Endress, A. D. The social sense: susceptibility to others beliefs in human adults and infants. Science 330, 1830–4 (2010).
Ronchi, R., Revol, P., Katayama, M., Rossetti, Y. & Farne, A. Seeing your error alters my pointing: observing systematic pointing errors induces sensori-motor after-effects. PLoS One 6, e21070 (2011).
Aglioti, S. M., Cesari, P., Romani, M. & Urgesi, C. Action anticipation and motor resonance in elite basketball players. Nat. Neurosci. 11, 1109–1116 (2008).
Mattar, A. G. & Gribble, P. L. Motor learning by observing. Neuron 46, 53–160 (2004).
Sebanz, N., Bekkering, H. & Knoblich, G. Joint action: bodies and minds moving together. Trends Cogn. Sci. 10, 70–77 (2006).
van der Wel, R. P. R. D., Knoblich, G. & Sebanz, N. Let the force be with us: dyads exploit haptic coupling for coordination. J. Exp. Psychol. 37, 1420–1431(2011).
Glynn, S. & Henning, R. A. Can teams outperform individuals in a simulated dynamic control task? Proc. Hum. Fact. and Erg. Soc. 6, 141–144 (2000).
Bosga, J. & Meulenbroek, R. G. Joint-action coordination of redundant force contributions in a virtual lifting task. Motor Cont. 11, 234–257 (2007).
Newman-Norlund, R. D., Bosga, J., Meulenbroek, R. G. & Bekkering, H. Anatomical substrates of cooperative joint-action in a continuous motor task: virtual lifting and balancing. Neuroimage 41, 169–77 (2008).
Knoblich, G. & Jordan, J. S. Action coordination in groups and individuals: learning anticipatory control. J. Exp. Psych. Learning, Mem. and Cogn. 29, 1006–1016 (2003).
Reed, K. B. & Peshkin, M. Physical collaboration of human-human and human-robot teams. IEEE Trans. Haptics 1, 108–120 (2008).
Ernst, M. O. & Banks, M. S. Humans integrate visual and haptic information in a statistically optimal fashion. Nature 45, 429–433 (2002).
Kording, K. & Wolpert, D. M. Bayesian integration in sensorimotor learning. Nature 427, 244–47 (2004).
Oztop, E., Wolpert, D. & Kawato, M. Mental state inference using visual control parameters. Cogn. Brain Res. 22, 129–51 (2005).
Wolpert, D. M., Doya, K. & Kawato, M. A unifying computational framework for motor control and social interaction. Philos. Trans. R Soc. Lond. B Biol. Sci. 358, 593–602 (2003).
Csibra, G. [Action mirroring and action understanding: an alternative account]. Sensorimotor foundations of higher cognition (Oxford University Press 2007).
Zentgraf, K., Munzert, J., Bischoff, M. & Newman-Norlund, R. D. Simulation during observation of human actions - Theories, empirical studies, applications. Vision Res. 51, 827–835 (2011).
Gallese, V. Before and below ‘theory of mind’: embodied simulation and the neural correlates of social cognition. Philos. Trans. R Soc. Lond. B Biol. Sci. 362, 659–69 (2007).
Gallese, V. & Goldman, A. Mirror neurons and the simulation theory of mind-reading. Trends in Cogn. Sci. 2, 493–501 (1998).
Marchal-Crespo, L. & Reinkensmeyer, D. J. Review of control strategies for robotic movement training after neurologic injury. J NeuroEng. and Rehab. 6, 10.1186/1743-0003-6-20 (2009).
Yarrow, K., Brown, P. & Krakauer, J. W. Inside the brain of an elite athlete: the neural processes that support high achievement in sports. Nat. Neurosci. Rev. 10, 585–96 (2009).
Flash, T. & Hogan, N. The coordination of arm movements: an experimentally confirmed mathematical model. J. Neurosci. 5, 1688–1703 (1985).
Bahrami, B., Olsen, K., Latham, P. E., Roepstorff, A., Rees, G. & Frith, C. D. Optimally interacting minds. Science 329, 1081–1085 (2010).
Balasubramaniam, S., Klein, J. & Burdet, E. Robot-assisted rehabilitation of hand function. Curr. Op. Neurol. 23, 661–670 (2010).
Yun, K., Watanabe, K. & Shimojo, S. Interpersonal body and neural synchronization as a marker of implicit social interaction. Sci. Rep. 2, 10.1038/srep00959 (2012).
Acknowledgements
This research was supported by a contract with the National Institute of Information and Communications Technology entitled, Development of network dynamics modeling methods for human brain data simulation systems', as well as MIC Information and Communication Technology R&D “Novel and innovative R&D making use of brain structure”. RO was supported by the Program for Next Generation World-Leading Researchers. EB and AT were supported in part by the EU FP7 grants HUMOUR (ICT-231724), CONTEST (PEOPLE-ITN-317488) and BALANCE (ICT-601003). The authors thank Dr. Wayne D. Dailey for his help with editing the manuscript.
Author information
Authors and Affiliations
Contributions
G.G. developed the concept. G.G. and E.B. designed the experiments. G.G., A.T. and T.Y. collected and analyzed data. G.G., M.K. and E.B. developed the analysis. G.G., R.O., M.K. and E.B. wrote the manuscript. All authors reviewed the manuscript.
Ethics declarations
Competing interests
Japanese patent 特願 2011-027711 (pending) on the use of collaborative paradigms for sports and rehabilitation.
Electronic supplementary material
Supplementary Information
Supplementary figures
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Ganesh, G., Takagi, A., Osu, R. et al. Two is better than one: Physical interactions improve motor performance in humans. Sci Rep 4, 3824 (2014). https://doi.org/10.1038/srep03824
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03824
This article is cited by
-
Low-force human–human hand interactions induce gait changes through sensorimotor engagement instead of direct mechanical effects
Scientific Reports (2024)
-
Learning acquisition of consistent leader–follower relationships depends on implicit haptic interactions
Scientific Reports (2023)
-
Leader–Follower Dynamics in Complex Obstacle Avoidance Task
International Journal of Social Robotics (2023)
-
Humans utilize sensory evidence of others’ intended action to make online decisions
Scientific Reports (2022)
-
Inter-personal motor interaction is facilitated by hand pairing
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
