
Abstract
Plant litter is an important component in wetland ecosystems, and the role of plant litter decomposition is considered to be important for wetland ecosystem functions and services. However, the consequences of litter inputs have seldom been experimentally tested in real ecosystems such as constructed wetlands (CWs). The enriched nutrients in CWs might weaken the role of litter inputs on soil carbon and nitrogen cycling. Here, we conducted a two-month field experiment to examine the effects of litter inputs on the soils in CWs. Our results showed that litter inputs significantly affected soil microbial (bacterial and fungi) diversities and properties (soil total nitrogen and nitrogen isotopes), and litter species with higher stoichiometry ratios, i.e. C/N, C/P and N/P led to higher microbial diversities. However, litter species had no or weak effects on microbial activities (CO2 and CH4 flux) or on the relative abundance of microbial communities, indicating that other environmental factors in such a CW might have stronger effects on those factors than litter inputs. These results highlighted the importance of submerged plant litter in nutrient-rich wetland ecosystems and provide potential tools for managers to improve the ecosystem functions and/or services via altering microbial diversities.
Similar content being viewed by others
Introduction
Plant litter as the end of primary production entering into detritus food chains plays an important role in wetland ecosystems, and its decomposition can recycle carbon and multiple nutrients, alter environmental variables, and affect wetland ecosystem functions and services1,2,3,4,5. Litter decomposition in wetland ecosystems refers to the respiration and assimilation of plant litter by microbes and invertebrates6, and it can be divided into three interlinked processes, i.e. leaching, fragmentation and microbial decay7,8. Moreover, litter decomposition are commonly considered to be affected by the quality of litter, associated soil microbes and invertebrates and the corresponding environmental factors7,9,10,11,12,13, but in wetlands such as stream and other freshwater ecosystems, litter inputs and biotic or abiotic factors are proven to be paramount14. Given the large variation in plant functional traits among plant species, litter decomposition rate can vary significantly among species due to the various ‘afterlife’ effects of litter traits (i.e. C/N, lignin, base cations and other decomposition related traits)15,16,17, and this interspecific variation in decomposition rates might lead to uncertainties in the effects of wetland plant litter on the soil or water qualities18,19, and thereby other organism in wetland ecosystems20,21,22.
Previous studies have proved the significant and diverse effects of plant litter on soil physio-chemical properties and microbial communities18,23,24,25,26,27,28, and there were plenty of evidence either from a specific ecosystem type at local scale or from different biomes at the global scale29,30. It has been proved that soil ecological processes, including soil C, N cycling, flux of CO2, CH4 are closely related to litter decomposition31,32. There are multiple pathways that plant litter can affect the soils in wetland ecosystems: (1) litter inputs can have various effects on soil invertebrates or microbes via different physical and chemical traits; (2) litter inputs can also provide food, microhabitat or shelter for soil microbes or other soil fauna33,34; (3) litter inputs can have negative effects via releasing leachate from litter which contains detrimental organic carbon and/or other compounds35,36,37. All above mechanisms indicated that plant litter traits as litter qualities can strongly influence the chemical and physical composition of litter inputs, and thereby their decomposability15 and lead to substantial consequences to wetland ecosystems. However, among various litter traits, litter stoichiometry might form the most important constraints of carbon: nitrogen ratios on soil microbial communities38,39 and hence act as the key trait to predict the effects of litter inputs on soil properties and microbial communities in wetlands40.
Moreover, the main methodology to test those effects of plant litter was firstly to sample soils from the field, and then either directly quantify the soil properties and microbial community composition or activity23,25; or subsequently set up new controlled experiments in the lab with the litterbag method or soil-litter mixing method18,26,27,41. For the latter case, knowledge about effects of litter mixing on soil is derived mostly from ‘indoor’ experiments carried out in strictly controlled environments, or via quantifying the litter mass loss and nutrient release42,43. However, very few investigators have addressed such effects of litter mixing on the wetland soils in relatively dynamic and unstable environments, such as constructed wetlands (CWs) with irregular waste-water inputs19. In such kind of constructed wetlands, other environmental factors might weaken the effects of litter inputs on the soil and thereby other soil organism, such as bacteria and fungi.
Now in this study, we set up a soil-litter mixing experiment (two months) in an ongoing constructed wetland located in Hanshiqiao wetland, Beijing, China (latitude: 40°07′21.0″, longitude: 116°48′56.7″). Our hypothesis is that the effects of litter inputs in such a constructed wetland might not be as overwhelming as those proven in indoor laboratory experiments, in which the related environmental factors are strictly controlled and relatively stable. We explored the effects of litter traits and incubation time on the soil properties of a constructed wetland including soil CO2/CH4 fluxes, total carbon (TC), total nitrogen (TN), carbon and nitrogen isotope changes (13C, 15N), as well as microbial diversities. The purpose of our work was to fully understand the role of plant litter in constructed wetlands and develop potential tools for the maintenance, improvement and management of constructed wetlands using plant litter materials.
Results
Effects of litter inputs on soil properties
There were significant differences between initial litter traits among four submerged plant species (Table 1). A significant difference was observed for CO2, but no significant difference in the other variables between initial soil and the control treatment (Fig. 1, ANOVA 1; Table S1). Moreover, litter of Potamogeton crispus significantly increased soil bacterial and fungi diversities, and 15N, but litter of Ceratophyllum demersum significantly increased soil TN, CO2 and CH4, but decreased 15N (Fig. 1, ANOVA 2; P < 0.05). However, litter species had no significant effects on either CO2 or CH4 (Fig. S1, Table S2).

The soil microbial diversity, soil CO2, CH4 flux and characters before (IN: initial soil without litter) and after two-month incubation of different litter species (C: Ceratophyllum demersum; H: Hydrilla verticillat; M: Myriophyllum verticillatum; P: Potamogeton crispus). Values are means ± S.E. (n = 3). CK indicates the control treatment. The dotted line is used to separate two groups of ANOVA analyses: (1) ANOVA 1 showed the results between initial soil properties and the control treatment after two-month incubation (without litter mixing). **Indicated P < 0.01; ns indicated no significant differences between initial soil and CK. (2) ANOVA 2 showed the results among different litter species, including CK. Values by the same letter indicated no significant differences (P > 0.05).
Effects of litter inputs on soil microbial communities
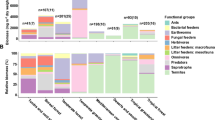
There were significant or marginally significant relationships between soil microbial diversity and litter stoichiometry ratios, i.e. C/N, C/P and N/P, but no significant relationships between soil microbial diversity and any other litter trait (Table 2). Moreover, after two-month incubation, one phylum of soil bacteria, i.e. Planctomycetes, significantly decreased, but another two phyla of soil bacteria, i.e. Bacteroidetes and Firmicutes, significantly increased (IN vs. CK: Fig. 2, P < 0.01). There were no significant differences in the soil fungi composition or the other phyla of soil bacteria before and after incubation (IN vs. CK: Fig. 2, P > 0.05). Overall, different species litter did not drive significant differences in the relative abundances of soil bacteria or fungi, except for some phyla (Firmicutes and Ciliophora) with lower relative abundance (<5%) (among four litter species and CK: Fig. 2, P > 0.05).

The relative abundance of bacteria and fungi in soil of different litter species before and after two-month incubation (IN: initial soil without litter mixing; CK: control treatment; C: Ceratophyllum demersum; H: Hydrilla verticillat; M: Myriophyllum verticillatum; P: Potamogeton crispus).
Discussion
Litter inputs might affect the soil ecological processes, including soil C and N cycling via litter decomposition31,32,44. We indeed observed significant effects of litter inputs on soil properties (soil TN, 13C and 15N) and microbial diversities, and those effects to some extent depended on litter species identity (Fig. 1). These results indicated that in such a real wetland ecosystem mixing litter with soil still played an important role in regulating wetland soils including isotope signatures and affecting the soil microbial diversities. However, this was different from our original hypothesis, and in a way highlighted the importance of (even a small amount) litter inputs might have significant effects on wetland soils. Note that given the initial litter was cut into small pieces before mixing, the observed effects might be strengthened compared to real litter inputs from submerged plants, which is similar to previous laboratory studies28,41,45.
The different responses of wetland soils to litter mixing might result from the different reactions of micro-organisms to different litter traits and/or to different chemical fractions released during litter decomposition processes46. In our study, we observed the highest TN and the lowest 15N in the soil mixed with C. demersum litter after two-month incubation (Fig. 1), and this might be due to the lowest 13C and 15N in the initial litter of C. demersum (Table 1). Isotope signatures can represent the ratios of heavier element (13C and 15N) to lighter element (12C and 14N), to some extent determining the decomposition rates of plant species litter47,48,49. Moreover, we also observed the highest microbial diversities and the highest 15N in the soil mixed with P. crispus litter after two-month incubation, and this might also result from the stoichiometric or isotope ratios of initial P. crispus litter (Table 1). The imbalance among plant litter, microbial biomass and soil stoichiometry might explain this phenomenon18,50, but regrettably we did not have the data for microbial stoichiometry. Instead, we indeed found litter inputs might offset the imbalance between soil and mixed litter, and there was a significant positive correlation between litter stoichiometric ratios and microbial diversities (Table 2), indicating that litter traits related to stoichiometry ratios were still important predictors for the effects of litter inputs on soil C and N cycling50. However, we did not find significant effects of litter inputs on the relative abundances of microbes except for several minor groups (Fig. 2), and the relative abundance of microbes might largely be determined by the long-term waste water inputs rather than the short-term litter inputs.
Litter inputs might also affect the CO2 and CH4 flux via the interactions between microbes and litter sources26,44,51, and litter traits such as C/N ratios were expected to drive those differences52,53. However, our results showed no significant differences in CO2 and CH4 flux among plant species (Table S2), but only the C. demersum litter led to the highest CO2 and CH4 flux after two months (Fig. 1). There might be due to the weaker effects of litter mixing than other factors from the constructed wetlands (Fig. 3), such as temperature and the C/N ratios of the waste water inputs. The continuous inputs of wasted water with extra carbon and nutrients in our study site might be more overwhelming than the effects of such a small amount of litter inputs. Note that the quantity of plant litter and the time of incubation might also matter27, and we suggested that future studies should incorporate both litter quality and quantity effects and put them in a relatively longer incubation period.

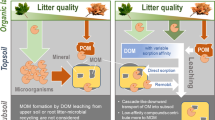
Conceptual framework and experimental arrangements for our study. Four litter species are: C: Ceratophyllum demersum; H: Hydrilla verticillat; M: Myriophyllum verticillatum; P: Potamogeton crispus; CK: control treatment. “ → ” in the constructed wetland is water flow direction.“⭐” is the study site.
Conclusion
In conclusion, our results provided empirical evidence for the effects of submerged plant litter on the soil properties and microbial diversities in a constructed wetland. These findings might have multiple implications for the design, maintenance and management of constructed wetlands: (1) when designing a constructed wetland, it is better not only take the species identity into consideration, but also for the stoichiometry of different growth forms19; (2) for constructed wetlands, plant litter of submerged plants should not have been always considered as wastes and being directly refloated from the CWs. Instead, it is possible to shift the ‘unfavored’ submerged plant litter to ‘useful’ tools to improve the ecosystem functions and services of constructed wetlands; (3) plant litter might be a feasible and economic materials for improving the microbial diversities of CWs, and it is worth to comprehensively study the role of wetland plants in constructed wetlands, especially for the role of plant litter, and this might provide valuable suggestions for managers about the maintenance and management of constructed wetlands.
Materials and Methods
Study site
Our study site was located in the Hanshiqiao wetland, Beijing, China. There was a constructed wetland, which was used to purify the waste water from the pleasure boat area, restaurant and the public toilet in the Hanshiqiao wetland Park (Fig. 3). The CW consists of seven treatment sections, and we conducted a soil-litter mixing experiment in one section of a constructed wetland (CW), i.e. the section III as the study site (marked as ✩ in Fig. 3). The area of the section III is 582.30 m2. The water average depth is about 2.5 m. The dominant plant species in section III are Iris wilsonii, Zizania latifolia, Typha orientalis and Sagittaria sagittifolia, and there were irregular waste water inputs flowing into our study site through early April to late November every year.
Experimental design
Plant litter of four submerged species (eg., Ceratophyllum demersum, Myriophyllum verticillatum, Hydrilla verticillata, Potamogeton crispus) were collected from Hanshiqiao wetland park in July 2017 (but not from the constructed wetland). All plant litter (Fig. 3, B1) was air-dried at room temperature for at least one month. The litter was subsequently cut into pieces (<5 mm) in order to increase the decomposition rate of litter and to maximize the effect of litter inputs on the soil in the CW. Initial soil samples (Fig. 3, A1) (upper 10 cm layer) were collected from five random locations in the CW by shovel, and then were thoroughly mixed. Any visible roots or contaminants were removed before the experiment. We prepared 15 plastic buckets (Fig. 3, the bottom diameter is around 22 cm and the height of the bucket is around 27.5 cm), and each bucket has only one litter species (litter species treatments) or no litter species (the control treatment). Within each plastic bucket, we put 400 g wetland soils and 4.0 g air-dried litter, and thoroughly mixed them in order to keep the dry-weight ratio of soil vs. litter consistently across treatments18,27. Finally, we randomly placed all the buckets at the bottom of our study site (submerged by water in the CW) and all the buckets were incubated in the same environment only with different litter input treatments. The distances between each bucket was around 30 cm. The whole experiment last from August 31, 2017 to November 2, 2017, which is also the main senesced period in the study region.
Sampling and measurements
Before incubation, three soil samples (Fig. 3, A2) and five litter samples for each species (Fig. 3, B2) were selected for the initial soil properties and initial litter trait measurements. The initial soil and litter measurements (listed in Table 1) included total carbon (TC), total nitrogen (TN), soil total phosphorus (TP), and stable isotopes (13C and 15N). Total C and Total N content of were assessed using the VarioMAX CN element analyzer (Macro Elemental Analyzer System GmbH, Hanau, Germany). The TP concentration was analyzed by inductively coupled plasma emission spectroscopy (Perkin Elmer Optima 3000 ICP Spectrometer, Waltham, MA, USA) and the isotopes of C and N were subsequently analyzed using an isotope ratio mass spectrometer (Isoprime100; Isoprime Ltd, UK).
Initial soil microbe community, i.e. bacteria and fungi, was a measured using the second generation next throughput sequencing technology (MiSeq high throughput sequencing, 16Sr DNA sequences). Microbial DNA was extracted from soil samples (or from litter-soil mixtures, see below) using the E.Z.N.A.® soil DNA Kit (Omega Bio-tek, Norcross, GA, U.S.) according to manufacturer’s protocols. The bacteria 16S and fungi 18S ribosomal RNA gene were amplified by PCR using primers 515F 5′-GTGCCAGCMGCCGCGG-3′, 907R 5′-CCGTCAATTCMTTTRAGTTT-3′and SSU0817F 5′-TTAGCATGGAATAATRRAATAGGA-3′ and 1196R 5′-TCTGGACCTGGTGAGTTTCC-3′ respectively, where barcode is an eight-base sequence unique to each sample. PCR reactions were performed in triplicate 20 μL mixture containing 4 μL of 5 × FastPfu Buffer, 2 μL of 2.5 mM dNTPs, 0.8 μL of each primer (5 μM), 0.4 μL of FastPfu Polymerase, and 10 ng of template DNA. Amplicons were extracted from 2% agarose gels and purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, U.S.) according to the manufacturer’s instructions and quantified using QuantiFluor™ -ST (Promega, U.S.). We calculated the Shannon diversity indices to represent the diversity of soil bacteria and fungi.
In addition, the CO2 and CH4 fluxes of soil was also measured. Concentrations of CO2 and CH4 were measured using a gas chromatograph (Agilent 7890 A, Santa Clara, CA). At first, we collected soils samples (at the beginning of experiment) or mixtures (2 weeks, 4 weeks, 6 weeks, 8 weeks) (about 10 g dry weight) from the CW by shovel or buckets by self-zip plastic bag. And they were putted in the glass bottle (100 ml). Before gas sampling, we sealed the glass bottle with airtight butyl rubber stoppers. After 24 h of incubation, the headspace gas of the glass bottle were sampled using airtight syringes. All gas samples were measured within 24 h after sampling.
During incubation, litter-soil mixtures were sampled every two weeks and then brought to the lab for the measurement of CO2 and CH4 fluxes. For the last sampling, the properties and microbial communities of litter-soil mixtures were again measured using the methods mentioned above. The whole experiment last two months from August 31, 2017 to November 2, 2017. After incubation, all the plastic buckets were removed from the CW to avoid the continuous disturbance for the CW.
Statistical analysis
All data were checked for assumptions of normality and homogeneity of variance before analysis. We firstly compared the differences of initial litter traits, such as TN, TC, TP, C/N, C/P, N/P, 13C, 15N, among four submerged plant species. Secondly, we conducted one way ANOVAs to examine the differences of soil microbes (relative abundance and Shannon diversity index), TC, TN, 13C, 15N, CO2 and CH4 between the initial soil (before incubation) and the control treatment after two-month incubation in the constructed wetland (Fig. 1, ANOVA 1) and examine the effects of plant species on soil microbes (relative abundance and diversity), TC, TN, 13C, 15N, CO2 and CH4 after two-month litter mixing (Fig. 1, ANOVA 2). Thirdly, we analyzed the relationship between initial litter traits (TN, TC, TP, C/N, C/P, N/P, 13C, 15N) and the change value of soil characters (microbial diversity, TC, TN, 13C, 15N, CO2 and CH4) before and after two-month incubation using regression analysis respectively. In the end, the effects of plant species on microbial respiration (CO2, CH4) during two-month litter mixing were analyzed using repeated measure ANOVA. Differences between means were tested with Fisher LSD tests; effects were considered significant at P < 0.05. All the ANOVA analyses were conducted in SPSS Statistics (SPSS, Chicago, IL, USA), and regression analyses were conducted in R software 3.5.2 (R core Team)54.
References
Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos 439–449 (1997).
Harmon, M. E. et al. Long‐term patterns of mass loss during the decomposition of leaf and fine root litter: an intersite comparison. Global Change Biol. 15, 1320–1338 (2009).
Kremen, C. Managing ecosystem services: what do we need to know about their ecology? Ecol. Lett. 8, 468–479 (2005).
Quétier, F., Lavorel, S., Thuiller, W. & Davies, I. Plant‐trait‐based modeling assessment of ecosystem‐service sensitivity to land‐use change. Ecol. Appl. 17, 2377–2386 (2007).
Schmid, B., Joshi, J. & Schläpfer, F. Empirical evidence for biodiversity-ecosystem functioning relationships. Functional consequences of biodiversity: Experimental progress and theoretical extensions 120–150 (2002).
Brinson, M. M., Lugo, A. E. & Brown, S. Primary productivity, decomposition and consumer activity in freshwater wetlands. Annu. Rev. Eco. S. 12, 123–161 (1981).
Swift, M. J., Heal, O. W. & Anderson, J. M. Decomposition in terrestrial ecosystems. Univ of California Press (1979).
Cotrufo, M. F., Ngao, J., Marzaioli, F. & Piermatteo, D. Inter-comparison of methods for quantifying above-ground leaf litter decomposition rates. Plant & Soil 334, 365–376 (2010).
Couˆteaux, M. M., Bottner, P. & Berg, B. Litter decomposition, climate and litter quality. Trends Ecol. Evol. 10, 63–66 (1995).
Royer, T. V. & Minshall, G. W. Rapid breakdown of allochthonous and autochthonous plant material in a eutrophic river. Hydrobiologia 344, 81–86 (1997).
Morris, J. T. & Bradley, P. M. Effects of nutrient loading on the carbon balance of coastal wetland sediments. Limnol. Oceanogr. 44, 699–702 (1999).
Liski, J., Nissinen, A. R. I., Erhard, M. & Taskinen, O. Climatic effects on litter decomposition from arctic tundra to tropical rainforest. Global Change Biol. 9, 575–584 (2003).
Rejmánková, E. & Houdková, K. Wetland plant decomposition under different nutrient conditions what is more important, litter quality or site quality? Biogeochemistry 80, 245–262 (2006).
Webster, J. R. & Benfield, E. F. Vascular plant breakdown in freshwater ecosystems. Annu. Rev. Eco. S 17, 567–594 (1986).
Cornelissen, J. & Thompson, K. Functional leaf attributes predict litter decomposition rate in herbaceous plants. New Phytol. 135, 109–114 (1997).
Cornwell, W. K. et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol.Lett. 11, 1065–1071 (2008).
Xie, Y. et al. Inhibition of litter decomposition of two emergent macrophytes by addition of aromatic plant powder. Sci. Rep. 7, 16685 (2017).
Cui, J. et al. Interactions between biochar and litter priming: A three-source 14C and δ13C partitioning study. Soil Biol. Biochem. 104, 49–58 (2017).
Pan, X. et al. Plant litter submergence affects the water quality of a constructed wetland. PloS one 12, e0171019 (2017).
Henriksen, T. & Breland, T. Nitrogen availability effects on carbon mineralization, fungal and bacterial growth, and enzyme activities during decomposition of wheat straw in soil. Soil Biol. Biochem. 31, 1121–1134 (1999).
Johnston, C. A. Sediment and nutrient retention by freshwater wetlands: effects on surface water quality. Crit. Rev. Env. Sci. Tec. 21, 491–565 (1991).
Pascoal, C. & Cássio, F. Contribution of fungi and bacteria to leaf litter decomposition in a polluted river. Appl. Environ. Microb. 70, 5266–5273 (2004).
Kourtev, P. S., Ehrenfeld, J. G. & Häggblom, M. Exotic plant species alter the microbial community structure and function in the soil. Ecology 83, 3152–3166 (2002).
Allison, S. D. A trait-based approach for modelling microbial litter decomposition. Ecol. Lett. 15, 1058–1070 (2012).
Schneider, T. et al. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. The ISME journal 6, 1749–1762 (2012).
Begum, N., Guppy, C., Herridge, D. & Schwenke, G. Influence of source and quality of plant residues on emissions of N2O and CO2 from a fertile, acidic Black Vertisol. Biol. Fert. soils 50, 499–506 (2014).
Chen, Y. et al. Chemical diversity and incubation time affect non-additive responses of soil carbon and nitrogen cycling to litter mixtures from an alpine steppe soil. Soil Biol. Biochem. 109, 124–134 (2017).
Chen, H., Li, B., Fang, C., Chen, J. & Wu, J. Exotic plant influences soil nematode communities through litter input. Soil Biol. Biochem. 39, 1782–1793 (2007).
De Deyn, G. B., Cornelissen, J. H. C. & Bardgett, R. D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 11, 516–531 (2008).
Fanin, N., Hättenschwiler, S., Barantal, S., Schimannb, H. & Fromina, N. Does variability in litter quality determine soil microbial respiration in an Amazonian rainforest? Soil Biol. Biochem. 43, 1014–1022 (2011).
Vargas, D. N., Bertiller, M. B., Ares, J. O., Carrera, A. L. & Sain, C. L. Soil C and N dynamics induced by leaf-litter decomposition of shrubs and perennial grasses of the Patagonian Monte. Soil Biol. Biochem. 38, 2401–2410 (2006).
Meier, C. L. & Bowman, W. D. Links between plant litter chemistry, species diversity, and below-ground ecosystem function. P. Natl. A. Sc. 105, 19780–19785 (2008).
Richardson, J. S. Food, microhabitat, or both? Macroinvertebrate use of leaf accumulations in a montane stream. Freshwater Biol. 27, 169–176 (1992).
Yee, D. A. & Juliano, S. A. Consequences of detritus type in an aquatic microsystem effects on water quality, micro‐organisms and performance of the dominant consumer. Freshwater Biol. 51, 448–459 (2006).
Horne, M. T. & Dunson, W. A. The interactive effects of low pH, toxic metals, and DOC on a simulated temporary pond community. Environ. Pollut. 89, 155–161 (1995).
Maerz, J. C., Brown, C. J., Chapin, C. T. & Blossey, B. Can secondary compounds of an invasive plant affect larval amphibians? Funct. Ecol. 19, 970–975 (2005).
Canhoto, C. & Laranjeira, C. Leachates of Eucalyptus globulus in intermittent streams affect water parameters and invertebrates. Int. Rev. Hydrobiol. 92, 173–182 (2007).
Cleveland, C. C. & Liptzin, D. C: N: P stoichiometry in soil: is there a ‘Redfield ratio’ for the microbial biomass? Biogeochemistry 85, 235–252 (2007).
Martinson, H. M. et al. Detritivory: stoichiometry of a neglected trophic level. Ecol. Res. 23, 487–491 (2008).
Sterner, R. W. & Elser, J. J. Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press (2002).
Lee, H. et al. Soil moisture and soil-litter mixing effects on surface litter decomposition: a controlled environment assessment. Soil Biol. Biochem. 72, 123–132 (2014).
Sun, Z. & Mou, X. Effects of sediment burial disturbance on macro and microelement dynamics in decomposing litter of Phragmites australis in the coastal marsh of the Yellow River estuary, China. Environ. Sci. Pollut. R. 23, 5189–5202 (2016).
Scarton, F., Day, J. W. & Rismondo, A. Primary production and decomposition of Sarcocornia fruticosa (L.) scott and Phragmites australis Trin. Ex Steudel in the Po Delta. Italy. Estuaries 25, 325–336 (2002).
Jiang, J. et al. Litter species traits, but not richness, contribute to carbon and nitrogen dynamics in an alpine meadow on the Tibetan Plateau. Plant & soil 373, 931–941 (2013).
Huang, Y., Zou, J., Zheng, X., Wang, Y. & Xu, X. Nitrous oxide emissions as influenced by amendment of plant residues with different C: N ratios. Soil Biol. Biochem. 36, 973–981 (2004).
Tardif, A. & Shipley, B. The relationship between functional dispersion of mixed‐species leaf litter mixtures and species’ interactions during decomposition. Oikos 124, 1050–1057 (2015).
Högberg, P. Tansley review no. 95 15N natural abundance in soil-plant systems. New Phytol 137, 179–203 (1997).
Kramer, M. G., Sollins, P., Sletten, R. S. & Swart, P. K. N isotope fractionation and measures of organic matter alteration during decomposition. Ecology 84, 2021–2025 (2003).
Craine, J. M. et al. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant & Soil 396, 1–26 (2015).
Kaiser, C., Franklin, O., Dieckmann, U. & Richter, A. Microbial community dynamics alleviate stoichiometric constraints during litter decay. Ecol. Lett. 17, 680–690 (2014).
Meier, C. L., Rapp, J., Bowers, R. M., Silmanb, M. & Fierer, N. Fungal growth on a common wood substrate across a tropical elevation gradient: temperature sensitivity, community composition, and potential for above-ground decomposition. Soil Biol. Biochem. 42, 1083–1090 (2010).
Aulakh, M. S., Walters, D. T., Doran, J. W., Francis, D. D. & Mosier, A. R. Crop residue type and placement effects on denitrification and mineralization. Soil Sci. Soc. Am. J. 55, 1020–1025 (1991).
Baggs, E. M., Rees, R. M., Smith, K. A. & Vinten, A. J. A. Nitrous oxide emission from soils after incorporating crop residues. Soil use manage. 16, 82–87 (2000).
R Core Team R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, http://www.R-project.org/ (2014).
Acknowledgements
This work was funded by the National Key R&D Program of China (2017YFC0506200) and Fundamental Research Funds for the Central Non-Profit Research Institution of CAF (CAFYBB2017SY045). We thank the teachers and students of Hangzhou Normal University for the assistance of analyzing litter qualities. We also thank Zhangjie Cai and Zhiguo Dou for helping with the experiment.
Author information
Authors and Affiliations
Contributions
L.J.C. and X.P. designed the experiment; Y.M.P. and X.P. executed the experiment; Y.M.P., X.P. and W.L. contributed to analyzing the data and making the figures. Y.M.P., X.P., W.L., J.Z.W. and L.J.C. contributed to writing and editing the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ping, Y., Pan, X., Li, W. et al. The soil bacterial and fungal diversity were determined by the stoichiometric ratios of litter inputs: evidence from a constructed wetland. Sci Rep 9, 13813 (2019). https://doi.org/10.1038/s41598-019-50161-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-50161-9
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
